









-
Fragrance is one of the important traits of ornamental plants. Many flower species, such as roses, peonies, lilies, and chrysanthemums, exhibit distinct fragrance profiles, which play a crucial role in the application and development of horticultural plants. Unique odors not only determine the interaction between plants and other organisms or abiotic environments, but also affect the sensory experiences of horticultural plants and their derivative products[1,2]. Among various fragrance types, sweet-aroma is associated with the positive emotions, such as happiness, and it is also an important indicator for evaluating product applications. Consequently, ornamental varieties with sweet fragrances, and their derivative products, are more likely to become best-sellers in the market[3]. However, most commercial rose varieties lack the sweet aroma, while only a few essential oil-producing germplasms possess fragrance with fresh, sweet aroma, such as Rosa damascena. In contrast, the rose varieties cultivated in China, such as R. rugosa 'Fenghua' and 'Kushui', have poor aroma traits with minimal sweet scents, significantly limiting their potential for the development of high-end products[4,5]. Therefore, elucidating the differentiation of scent traits in rose petals, particularly the formation of sweet-aroma, is of great significance for improving fragrance traits of rose and developing their derivative products.
The fragrance in plants is identified based on various volatile organic compounds (VOCs). The content and the odor activity value (OAV) of each VOC codetermines the perception of aroma characteristics, among which OAV is a crucial indicator for assessing the importance of aromatic compounds[6]. The higher the OAV level is, the greater the contribution of the substance to the overall fragrance. The key aromatic compounds and fragrance trait differentiation could be identified through OAV analysis. For example, Wuyi rock tea is characterized by a distinctive 'rock flavor', which has been attributed to 2-ethyl-3,5-dimethylpyrazine, a substance with a roasted odor, through OAV analysis and aroma recombination experiment[7]. The scents of 17 peony cultivars were divided into three types based on OAV analysis, while four compounds, namely citronellol, geraniol, linalool, and β-phenylethanol, were found as the major characteristic aroma substances in the petals by measuring VOC content[8]. Furthermore, 17 tree peony cultivars were classified into four distinct groups, i.e., flower-sweet scent, flower-rose scent, herb scent, and mixed scent, among which linalool contributed 94.5% to the aroma in flower-sweet scented cultivars[9]. Previous studies related to rose fragrance, mainly focused on the absolute content of each VOC, which cumulatively measured the VOC profiles in the petals of more than 20 rose germplasm, and then suggested that citronellol and β-phenylethanol were the main VOCs with the highest content[10−13]. In some plant species, the note of β-phenylethanol is generally classified as rose scent along with citronellol and geraniol[8−9], however, which cannot explain the differences in fragrance types among different rose varieties, especially for the varieties with strong sweet-aroma, likely R. damascena and R. rugosa 'Bazizhi'[4,13]. Therefore, studying on the content and OAV levels of VOCs in rose could provide a basic for identifying the key aromatic compounds associated with the differentiation of fragrance types and the formation of sweet-aroma.
The formation of fragrance traits in higher plants is associated with the biosynthesis and accumulation of VOCs, especially the terpenoids and benzenoids/phenylpropanoids. Terpenoids, the most diverse class of VOCs, are synthesized using prenyl diphosphates as precursors, likely geranyl pyrophosphate (GPP), farnesyl diphosphate (FPP), and geranyl geranyl diphosphate (GGPP)[14]. These precursors are catalyzed by members of the terpenoid synthase (TPS) family to produce various terpenoids, including monoterpenoids, diterpenoids, and sesquiterpenoids[15−17]. Benzenoids/phenylpropanoids are another important VOC category that influences the formation of fragrance traits. β-Phenylethanol is a major benzenoid/phenylpropanoid compound in many species, and its biosynthesis is catalyzed by a series of enzymes, such as aromatic amino acid transaminase (AAAT) and phenylpyruvate decarboxylase (PPDC), alcohol dehydrogenase (ADH) and phenylacetaldehyde reductase (PAR)[18,19]. The formation of fragrance traits is the result of the coordinated action of numerous structural genes involved in VOC biosynthesis. In previous studies, a series of genes have been identified to be related to aromatic substance biosynthesis in Rosa species[17−19]. However, the genetic basis for the spectrum of scent phenotypes and their regulatory mechanisms across different rose germplasms has not been uncovered. The genes related to key aromatic compounds play important roles in the differentiation of fragrance traits, which could serve as key targets for modifying the fragrance traits of horticultural plants through biotechnological approaches[8,20]. Therefore, identifying the key genes regulating the biosynthesis of rose aromatic components and the differentiation of petal fragrance is of great significance for the targeted improvement of rose fragrance traits and the breeding of novel varieties with unique aromas.
In this study, a total of 32 rose cultivars were selected for VOC detection using solid-phase microextraction coupled with gas chromatography-mass spectrometry (SPME-GC/MS). By measuring the VOC profiles, calculating the OAV levels, and performing the related gene expressions, the following questions were addressed: (1) What is the diversity of VOCs in rose petals? (2) What are the key characteristic components that affect the differentiation of fragrance traits, especially the formation of sweet-aroma? (3) Which key genes are responsible for the formation of fragrance with a sweet scent? These answers could provide gene resources and theoretical foundations for targeted improvement of rose fragrance traits and the cultivation of novel varieties.
-
The rose resources sampled in this study were growing under natural conditions at the Rose Germplasm Nursery, Pingyin Rose Institute, Jinan, China (36°15′24″ N, 116°23′29″ E). In late April 2024, the plants with similar growth conditions of 32 varieties were selected for VOC measurement (Supplementary Table S1). At 5:00 a.m. to 6:00 a.m. on May 2nd, the petals at the half-opening stage, from three individual plants of each germplasm were sampled and pooled, immediately frozen in liquid nitrogen, and then stored at –80 °C for further study.
SPME-GC/MS analysis
-
After crushing into pieces under liquid nitrogen conditions for weighing accurately, 1.0 g of petals were weighed and then transferred into a 50 mL conical flask sealed with filter paper. A total of 10 μL of 3-nonanone solution (0.3 μg·μL−1) was added as an internal standard substance. The vial was incubated for 35 min at 45 °C for VOC adsorption by the SPEM syringe (65 μm PDMS/DVB, Supelco, USA). The syringe was then put into the injector port of the GC-MS for desorption for 3 min at 250 °C. The aromas absorbed on the SPME were analyzed using a GCMS-QP2010 Plus system (Shimadzu, Japan) equipped with an Rtx-5MS column (30 m × 0.25 μm × 0.25 mm). The GC condition and MS method were set according to previous studies[21]. The injector temperature was 250 °C and helium was used as a carrier gas at a flow rate of 1 mL·min−1. The initial GC temperature was maintained for 2 min at 40 °C, gradually increased to 100 °C at 5 °C·min−1, and then maintained for 1 min at 100 °C. It was then raised to 160 °C at 10 °C·min−1 and maintained for 1 min at 160 °C. Finally, it was increased to 250 °C at 15 °C·min−1 and maintained for 5 min. Mass spectrometry was operated in full scan mode over an m/z range of 35−450.
Volatile metabolome analysis
-
The VOCs were analyzed qualitatively and quantitatively. Qualitative analysis was performed according to the NIST 17s standard library (Hewlett Packard, USA). Spectrometric data were compared with the mass spectrum obtained from the original NIST 17s library and the Wiley library. The mass spectra were manually resolved and confirmed by comparing the Kovats' retention indices and relative reports from the literature. Only results identified with positive and negative matching values of more than 800 (the maximum is 1,000) were selected and identified. Quantitative analysis was done using 3-nonanone as the internal standard through the response factor method combined with the internal standard method. The mass fraction of headspace compounds was calculated using the following equation: Content of each component (μg·g−1 FW) = [Peak area of each component / Peak area of internal standard × Concentration of internal standard (μg·μL−1) × Volume of internal standard (μL)]/Fresh sample weight (g).
Calculation of odor activity values
-
To assess the influence of the VOCs on the overall rose petal aroma, the OAVs were calculated by dividing the concentrations of aroma compounds by their odor thresholds. The odor descriptions of VOCs and odor thresholds in water were mainly as referred to by Burdock[22], and the Volatile Compounds in Food online database (
www.vcf-online.nl/VcfHome.cfm ). OAVs are calculated as follows: OAV = Each component content (μg·g−1 FW) / Odor threshold (μg·g−1). Only the compounds with an OAV greater than 1 contribute individually to the rose petal aroma.Gene expression analysis
-
For analyzing the molecular mechanism of aromatic substance biosynthesis, the expression patterns of aroma-related structural genes were analyzed by quantitative reverse transcription PCR (RT-qPCR), in accordance with our previous study[23]. Based on our previous transcriptome data of transcriptome profiles of rose petals at different developing stages (stored in NCBI with the accession number of SRR34878993~SRR34879010), a total of 14 structural genes involved in the biosynthesis of key characteristic components were selected to measure their expression trends in four representative rose germplasms. Then, five key genes identified by correlation analysis were verified in 21 germplasms. All primers are listed in Supplementary Table S2. All actions were performed on a CFX Opus 96 Real-time System (Bio-Rad, USA) using SYBR Green Dye with three independent biological replicates for each sample. The expression patterns were analyzed using the 2−ΔΔCᴛ method, with Actin as the internal control[18].
Gene functional verification
-
Germplasm #27 was selected for biological function verification of RrTPS1 by overexpression and VIGS experiments. Twigs with uniform flower buds at early-opening stage were cut off, and then taken back into the laboratory through hydroponic cultivation. The RrTPS1 CDS was fused into the pCY-H05252 vector. A 210-bp fragment cloned from RrTPS1 CDS was inserted into the pTRV2-GFP vector, with TRV1 as a helper plasmid. After transformation and incubation, the pCY-RrTPS1 and the mixed pTRV1 + pTRV2-RrTPS1 bacterial solution were injected into the petals of isolated flower buds of #27, with the petals harboring the empty vector as the control (pCY-H05252 for overexpression and pTRV1 + pTRV2 for VIGS). The petals were incubated in petri dishes for 24-h dark/48-h light cultivation for overexpression, and 24-h dark/72-h light cultivation for silencing expression, respectively. The petal area with GFP signal was collected for VOC profile measurement. Germplasm #30 was selected for the function analysis of RrAAAT1 and RrPAR2. The co-expression of RrAAAT1 and RrPAR2 was also performed via the experiments above[23].
Statistical analysis
-
Values are expressed as mean ± SD (n = 3). The principal component analysis (PCA), and hierarchical cluster analysis (HCA) were performed via Metware Cloud (
https://cloud.metware.cn ). One-way analysis of variance (ANOVA) with repeated measures was used to test for changes in aroma content among different varieties. Pearson correlation coefficients were used to examine relationships between volatile metabolites within species. All statistics were performed using Origin 2022 (OriginLab, USA). For all parameters measured, α was set at 0.05 and p values less than α were considered significant. -
The VOC profiles among 32 rose germplasms were detected to comprehensively investigate the changing patterns of floral fragrance. Both principal component analysis (PCA) and hierarchical cluster analysis (HCA) indicated the data had good repeatability (Supplementary Fig. S1). A total of 80 VOCs were identified in rose petals. The total VOC contents ranged from 6.16 ± 0.03 μg·g−1 FW (#01) to 28.26 ± 0.18 μg·g−1 FW (#32), with an average of 13.80 μg·g−1 FW (Fig. 1a, Supplementary Table S1). The VOC types varied between 40 (#24) to 61 (#20), and the average number was 49. Taking 10 and 20 μg·g−1 FW as references, all the rose germplasms were classified into three clusters with low (Group I, 15 germplasms), medium (Group II, 8), and high (Group III) VOC contents, respectively. The germplasms belonging to Group III were regarded as important resources for developing new rose varieties with strong-scent.
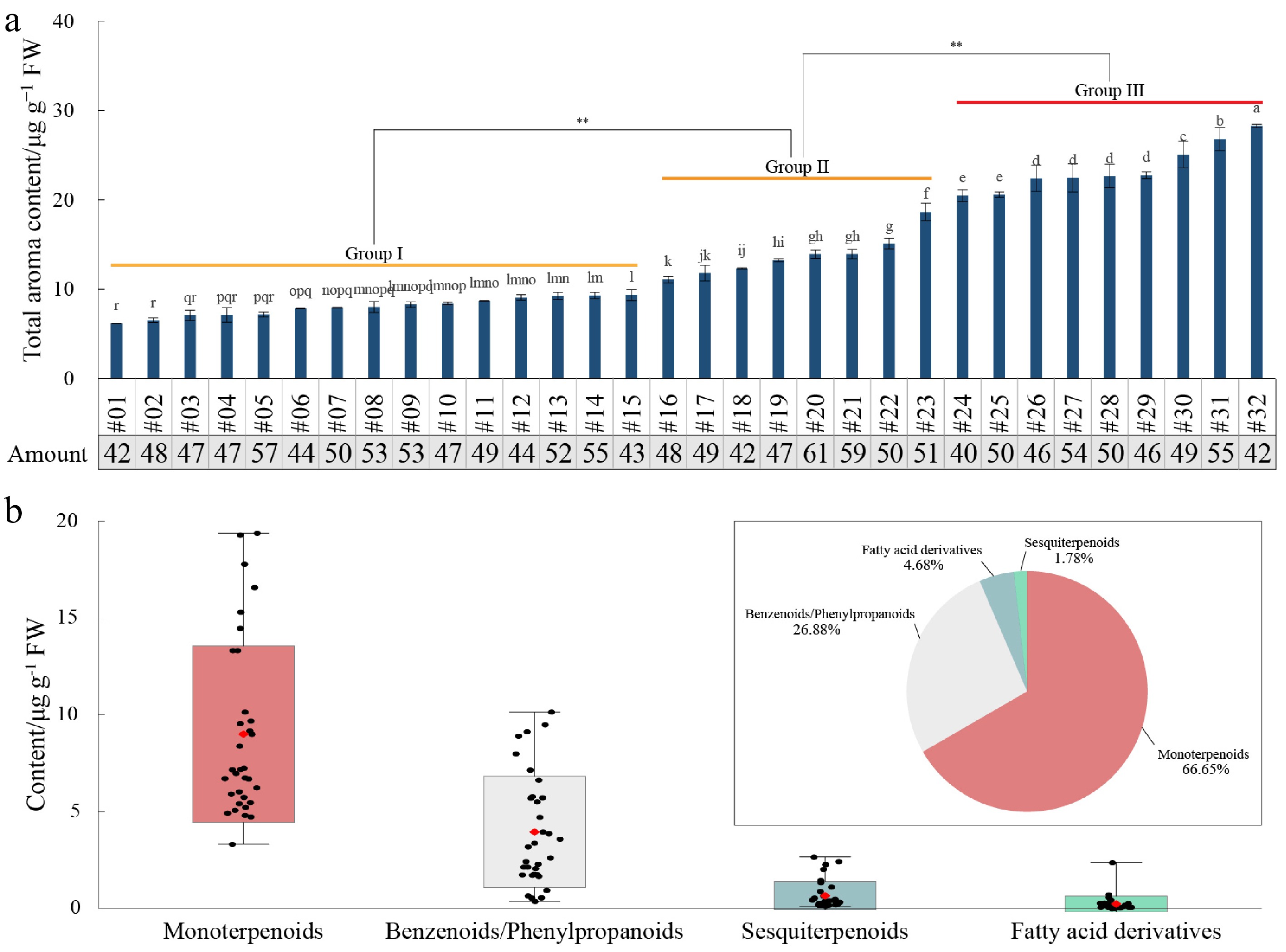
Figure 1.
The change trends of VOCs in the petals of 32 rose germplasms. (a) The total contents and types of VOCs. Different lower case letters indicate significant differences between germplasms at α = 0.05. ** Indicates significant differences at the 0.01 level. (b) Distribution of four VOC categories.
Based on their chemical structures, the detected substances were classified into three major classes (Supplementary Table S3): terpenoids (39), benzenoids/phenylpropanoids (nine), and fatty acid derivatives (32). Among these, terpenoids were further divided into monoterpenoids (25) and sesquiterpenoids (14). Among the 32 rose germplasms, monoterpenoids were the predominant compounds, accounting for 39.01% (#18) to 94.66% (#24) of the total VOC content, with an average of 66.65% (Fig. 1b, Supplementary Table S4). This was followed by benzenoids/phenylpropanoids, which accounted for between 2.67% (#24) and 58.06% (#18) of the total VOC content, with an average of 26.88%. Fatty acid derivatives and sesquiterpenoids had lower contents, accounting for only 4.68% and 1.78%, respectively. Therefore, monoterpenoids and benzenoids/phenylpropanoids were the main VOC categories in rose petals, of which relative contents showed significant variation among different germplasms.
Geraniol, citronellol, β-phenylethanol, and linalool were the key characteristic components of fragrance traits in rose petals
-
To seek out the predominant aromatic components in rose petals, the change trends of monoterpenoids and benzenoids/phenylpropanoids across 32 rose germplasms were analyzed (Fig. 2). For the 25 monoterpenoids, citronellol exhibited the highest average content at 3.44 μg·g−1 FW in 32 rose germplasms, representing 38.04% of the total monoterpenoid content. This was followed by geraniol (2.77 μg·g−1 FW), and α-citral (0.59 μg·g−1 FW), which accounted for 31.69% and 6.07%, respectively. β-Phenylethanol was the most abundant benzenoid/phenylpropanoid (3.47 μg·g−1 FW), representing 83.27% of the total content of 9 benzenoids/phenylpropanoids. Therefore, based on content level, geraniol, citronellol, and β-phenylethanol were the predominant aromatic substances in rose petals.
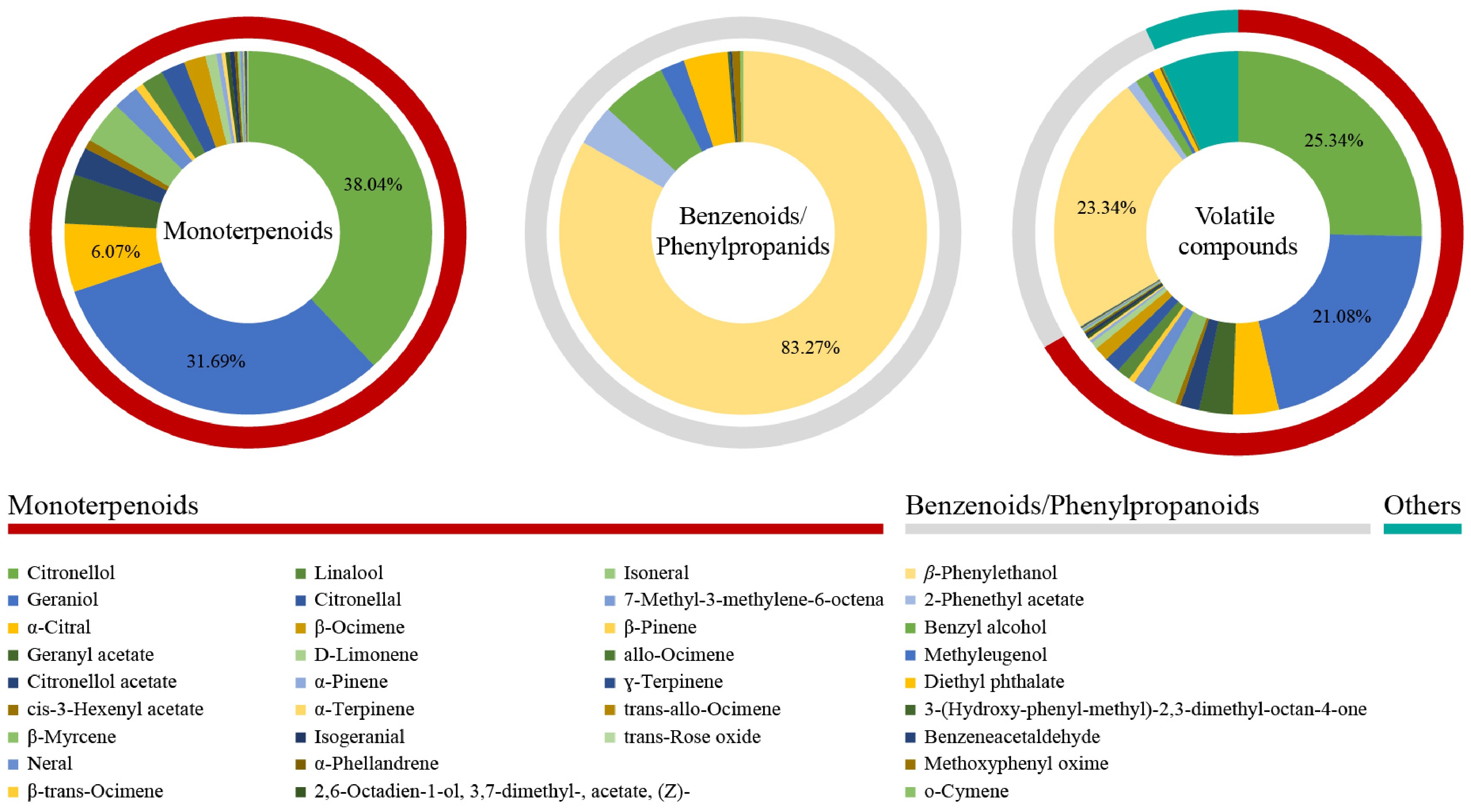
Figure 2.
Compound composition of monoterpenoids and benzenoids/phenylpropanoids in the petals of 32 rose germplasms.
By calculating OAVs, a total of 15 aromatic compounds, with OAVs greater than 10 were identified (Fig. 3, Supplementary Table S5). Among these, geraniol had the highest OAV level of 363.83, and all rose germplasms exhibited geraniol OAVs greater than 80. Following geraniol, citronellol demonstrated the second-highest average OAV of 312.31, with all germplasms showing citronellol OAVs greater than 80 except #01. The average OAV of linalool in 32 rose petals was as high as 100.29, and that was greater than 80 in 12 rose germplasms, because of its low odor threshold (only 0.0015 μg·g−1). β-Phenylethanol has a higher odor threshold (0.045 μg·g−1), resulting in a low average OAV level of 77.28. The OAVs of β-phenylethanol in all germplasm with high aromatic contents from Class III were greater than 80, except for #24. Additionally, other aromatic substances, such as cis-3-hexenyl acetate, β-myrcene, and α-citral, showed relatively high OAVs in certain germplasms; however, which were significantly lower than the OAV levels of major components like citronellol and geraniol. For example, β-myrcene in germplasm #24 had an OAV of 62.48, which was only 1/11 and 1/10 of the OAVs of citronellol (635.73) and geraniol (720.57), respectively. Therefore, from the perspective of odor activity values, geraniol, citronellol, linalool, and β-phenylethanol were deemed the key characteristic aromatic components in rose petals.
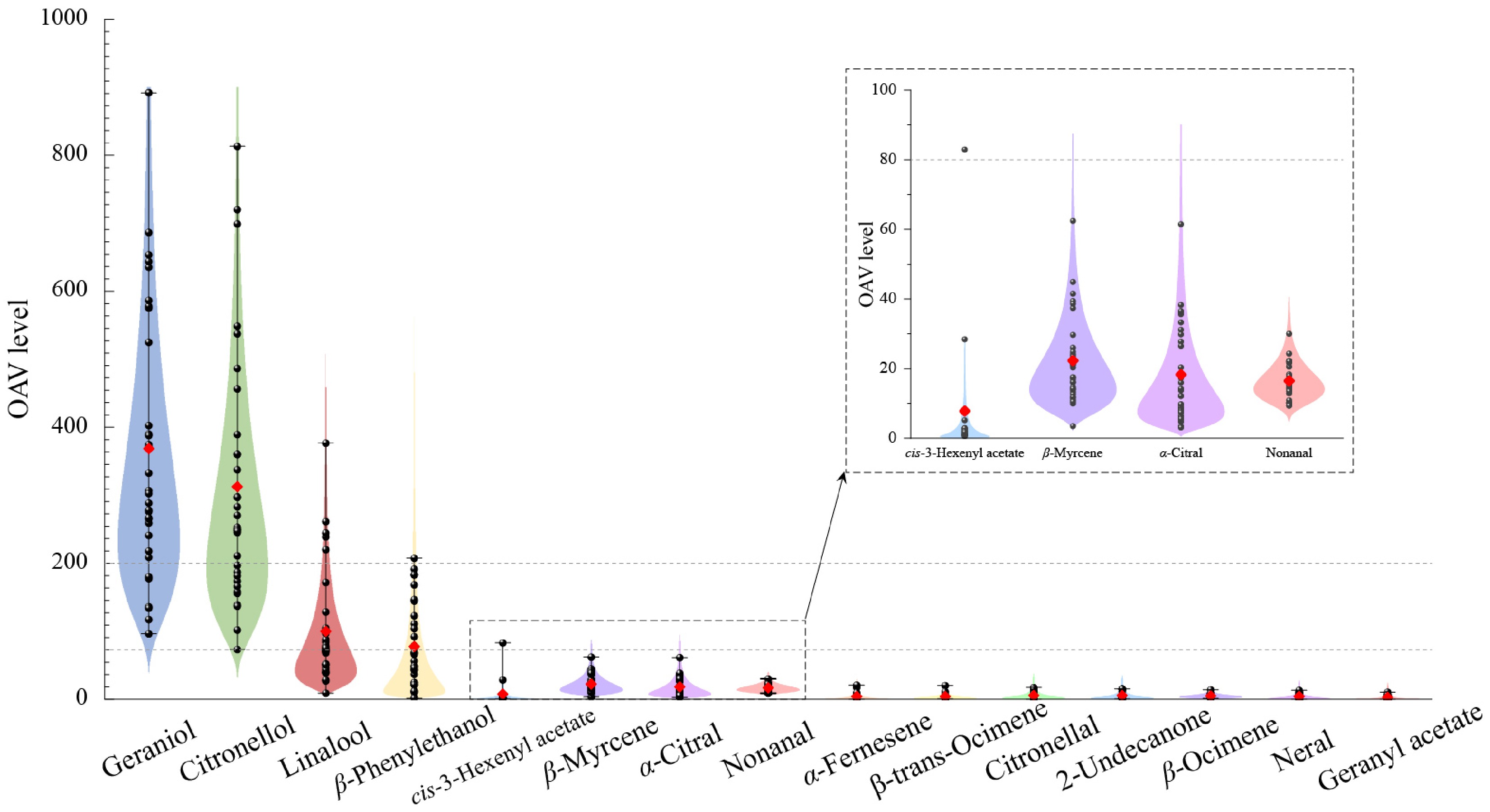
Figure 3.
The OAV changes of 15 aromatic compounds in the petals of 32 rose germplasms.
The scents of 32 rose germplasms were divided into five types based on the OAVs of four key characteristic aromatic components
-
To define the scent types of different rose germplasms, the odor descriptions of all aromatic components in rose petals were discerned and categorized. A total of 15 aromatic compounds with OAV greater than 10 were classified into eight scent types: balsamic, fatty, floral-geranium, floral-green rose, floral-lily, floral-sweet rose, fruity-citrus, and herbal (Supplementary Table S5). A fragrance radar chart of 32 rose germplasms was built based on the OAV percentages of eight types calculated by 15 aromatic compounds (Fig. 4a). The four scent types with high OAV percentages were floral-geranium (38.74%), floral-green rose (32.81%), floral-lily (10.99%), and floral-sweet rose (7.86%), which were primarily contributed by four key aromatic components: i.e., geraniol, citronellol, linalool, and β-phenylethanol. The cumulative OAVs of these four key components accounted for 90.40% of the total OAVs in rose petal, suggesting their potential as representative components for identifying fragrance types in rose germplasms. To validate this hypothesis, a fragrance radar chart of the OAV percentage of these four key components was drawn, which was highly consistent with the chart based on the 15 aromatic compounds (Fig. 4b). Consequently, these four compounds, namely geraniol, citronellol, linalool, and β-phenylethanol, emerged as representative components for identifying fragrance types in rose petals and are important target substances for improving the aromatic traits of roses.
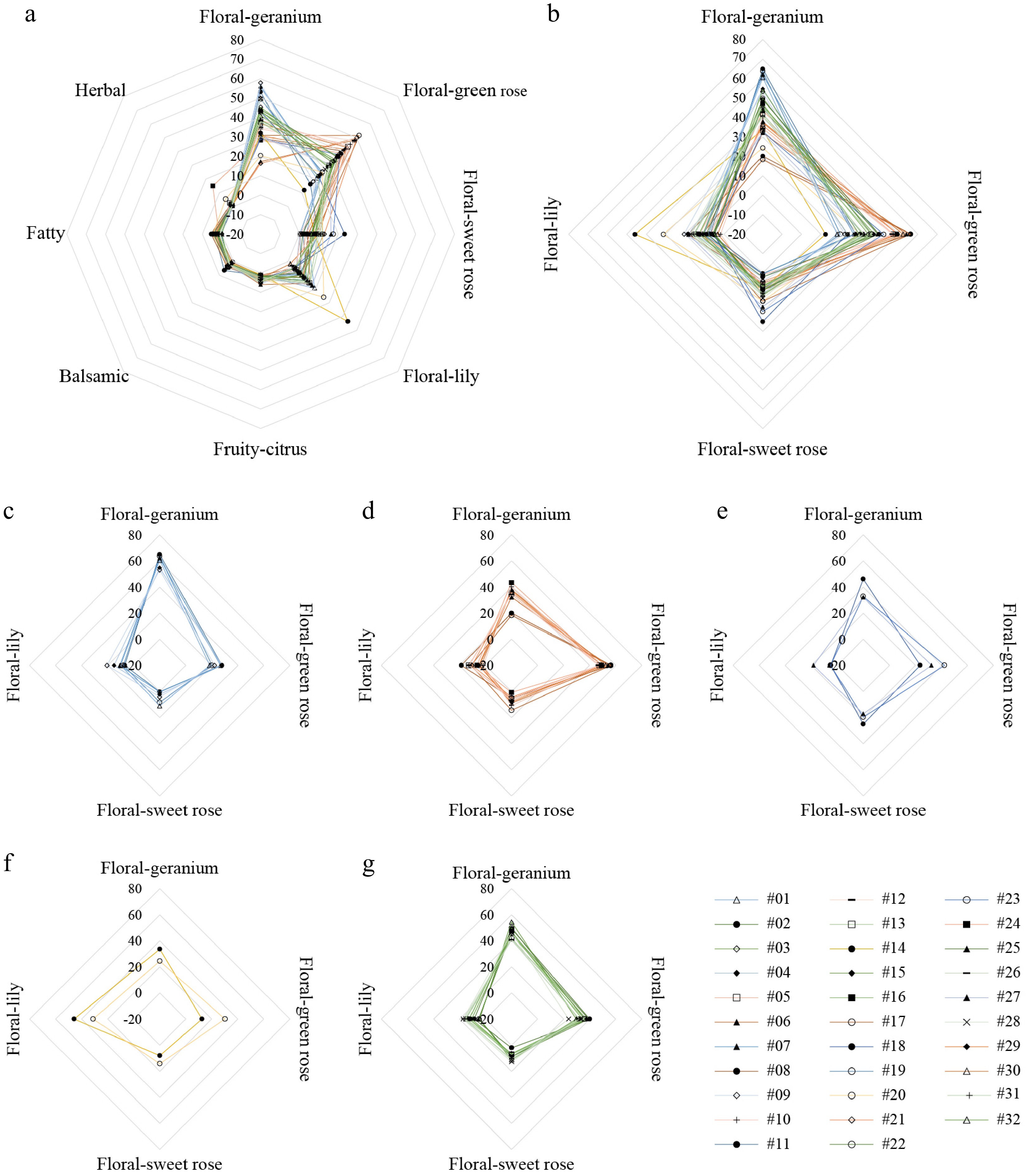
Figure 4.
The radar chart of different scent types in the petals of 32 rose germplasms. (a) The radar chart of eight scent types. (b) The radar chart of four main scent types. (c) The scent traits of floral-geranium scent germplasms. (d) The scent traits of floral-green rose scent germplasms. (e) The scent traits of floral-sweet rose scent germplasms. (f) The scent traits of floral-lily scent germplasms. (g) The scent traits of floral-mined scent germplasms.
Based on the OAV percentages of four key characteristic components, the scents of 32 rose germplasms were classified into five types.
(1) Floral-geranium scent. The fragrance of germplasms from this type was a mild, sweet rose odor, with a slight bitter taste. The primary key component was geraniol, which accounted for more than 50% and 33% of the cumulative OAV and total content of four components, respectively. This category included six germplasms: #01, #04, #07, #09, #11, and #19 (Fig. 4c).
(2) Floral-green rose scent. The germplasms emitted a fresh, green rose scent with a lemon aroma, with citronellol being the predominant component that accounted for more than 40% and 33% of the total OAV and content, respectively. Ten germplasms were found in this category: #05, #06, #08, #10, #12, #17, #21, #24, #29, and #30 (Fig. 4d).
(3) Floral-sweet rose scent. These germplasms exhibited a sweet and pleasurable rose scent that was related to β-phenylethanol, mainly representing over 15% and 45% of the total OAV and content, respectively. This category contained three germplasms: #18, #23, and #27 (Fig. 4e).
(4) Floral-lily scent. The fragrance of germplasms from this type was identified as a sweet odor, obtaining a strong aroma of lily of the valley, accompanied by a woody note, which was contributed by linalool, with percentages over 25% and 3% for total OAV and content, respectively. Only two germplasms were found in this category: #14 and #20, whose fragrance was evaluated as sweet-scent in the industry (Fig. 4f).
(5) Floral-mined scent. In this fragrance type, both geraniol (OAV ratio: 35%~50%, relative content: 20%~35%) and citronellol (20%~35% and 20%~35%) were the main aromatic components. Eleven germplasms were identified in this category: #02, #03, #13, #15, #16, #22, #25, #26, #28, #31, and #32 (Fig. 4g).
The sweet-aroma formation may be related to the different expressions of RrTPS1, RrAAAT1, and RrPAR2
-
To further reveal the molecular basis of fragrance trait formation in the rose petals, a total of nine genes involved in monoterpenoid accumulation and five genes associated with β-phenylethanol biosynthesis were selected based on our previous transcriptome profiles (Supplementary Table S2). The expression levels of these 14 structural genes were measured among five representative germplasms, namely #07, #20, #27, #30, and #32 (Fig. 5a). Correlation analysis suggested that five structural genes, i.e., Nudix hydrolase (RrNUDX1-1a), cinnamyl alcohol dehydrogenase (RrCAD1), RrTPS1, RrAAAT1, and RrPAR2, were significantly associated with variations in contents of key characteristic components (Fig. 5b). Among these, the expression of RrNUDX1-1a exhibited significant positive correlations with both terpenoid (0.91) and monoterpenoid (0.92) contents. RrCAD1 expression was significantly positively correlated with citronellol content, with a correlation coefficient of 0.93. A significant correlation was identified between the expression of RrTPS1 and the content of linalool (0.96). Additionally, RrAAAT1 and RrPAR2 expressions were significantly positively correlated with β-phenylethanol content, with correlation coefficients of 0.95 and 0.96, respectively.
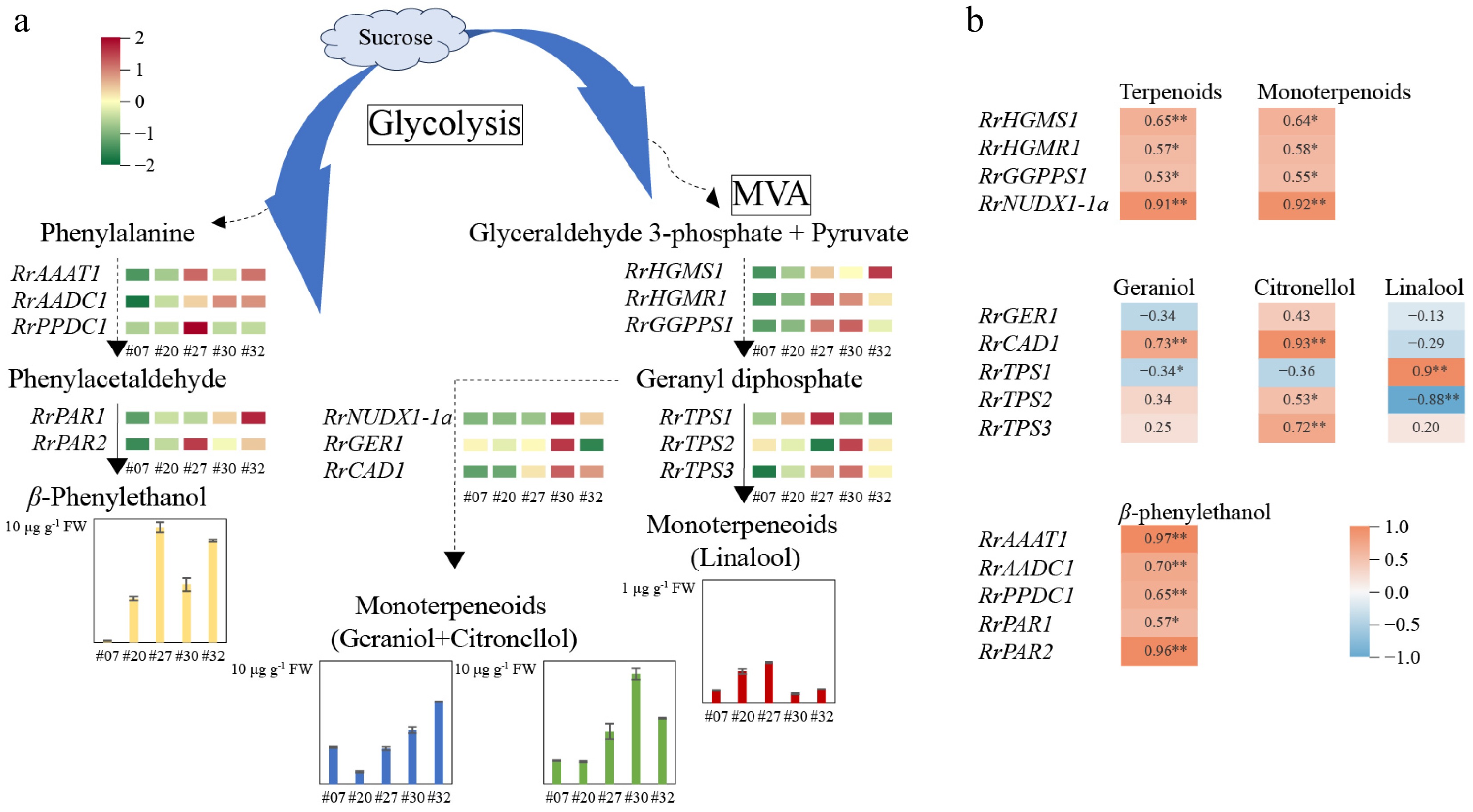
Figure 5.
The level changes of key characteristic component contents and related gene expressions in the petals of five rose germplasms. (a) Change of aromatic contents and gene expression patterns. (b) Correlation analysis results. ** Indicates a correlation significant at the 0.01 level.
Moreover, the expression patterns of these five structural genes, across 21 rose germplasms, were performed for further confirmation of the relationship between key regulatory gene expressions and aromatic substance accumulation (Fig. 6). The expression of RrCAD1 had a significantly positive correlation with citronellol content (0.76), while RrNUDX1-1a expression showed obviously positive correlations with monoterpenoid (0.72) and citronellol (0.77) contents. Moreover, RrTPS1 expression was significantly positively correlated with linalool content (0.73). RrAAAT1 and RrPAR2 expressions were closely related to β-phenylethanol content, with the correlation coefficients of 0.70 and 0.72, respectively. Therefore, the five genes, namely RrNUDX1-1a, RrCAD1, RrTPS1, RrAAAT1, and RrPAR2, had significant relationships with the accumulation of geraniol, citronellol, linalool, and β-phenylethanol in rose petals. Moreover, RrTPS1, RrAAAT1, and RrPAR2 may regulate the biosynthesis of linalool and β-phenylethanol, acting as key candidate genes for the formation of sweet-aromas.

Figure 6.
The level changes of four key characteristic components and five key gene expressions in the petals of 21 rose germplasms. (a) Change of aromatic contents. (b) Change of gene expressions. (c) Correlation analysis results. ** Indicates a correlation significant at the 0.01 level.
RrTPS1, RrAAAT1, and RrPAR2 regulated the formation of sweet aroma in rose
-
To further verify the biosynthesis functions of RrTPS1, RrAAAT1, and RrPAR2 in sweet-aroma trait formation, transient expression experiments were performed in #27 and #30 petals. Compared with the empty vector, overexpression of RrTPS1 in #27 petals significantly increased the content of linalool by 135.67%, while the levels of other key components had no significant changes (Fig. 7a). Consistently, the odor level of floral-lily scent contributed by linalool increased from 36.98 to 93.64, with its relative proportion elevating from 16.93% to 33.77%. The single overexpression and co-expression of RrAAAT1 and RrPAR2 were performed in #30 petals, among which the co-expression experiment showed more remarkable change trends in β-phenylethanol content (Fig. 7b and Supplementary Fig. S2). Co-expression of RrAAAT1 and RrPAR2 in #30 petals resulted in 11.23- and 13.71-fold increases in their respective gene expressions, respectively. The content of β- phenylethanol showed a significant up-trend of 203.58%, while the contents of other substances remained unchanged. The OAV of floral-sweet rose scent contributed by β-phenylethanol increased from 22.24 to 67.51, and its relative proportion increased from 6.27% to 16.08%. Furthermore, the VIGS experiments of these three key genes were performed, respectively (Supplementary Fig. S3). The silencing expression of RrTPS1 led to a 55.25% decrease in the content of linalool in #27 petals, while the silencing expression of RrAAAT1 and RrPAR2 resulted in downward trends in β-phenylethanol contents by 51.34% and 36.56% in #30 petals, respectively. Consequently, RrTPS1, RrAAAT1, and RrPAR2 significantly regulated the absolute and relative contents of linalool and β-phenylethanol, which could enhance the fragrance traits of rose petals by increasing the proportion of key target substances with a sweet scent.
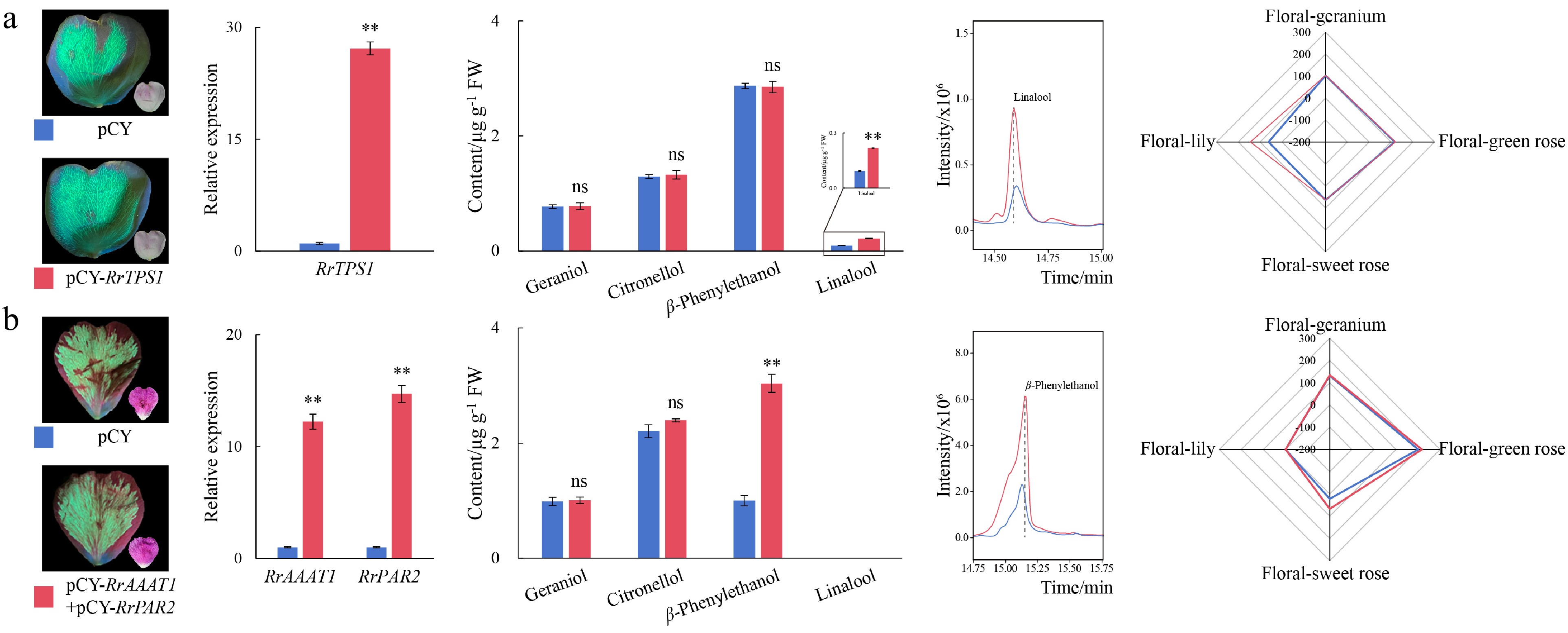
Figure 7.
The changes of fluorescent phenotype, gene expression, aromatic compound content, and OAV levels after transient expression of RrTPS1, and co-expression of RrAAAT1 and RrPAR2. (a) The changes after overexpression of RrTPS1. (b) The changes after co-expression of RrAAAT1 and RrPAR2. ** Indicates significant differences at the 0.01 level. ns indicates no statistically significant difference.
-
The ornamental traits in horticultural plants include florescence, flower shape, color, and fragrance, among which the research on flower fragrance is currently relatively scarce[24−26]. Fragrance trait is an important indicator for the product quality evaluation and variety cultivation of aromatic plants[4]. Peony varieties were classified into different grades with strong fragrance, light fragrance, and no fragrance through sensory evaluation, which was closely related to the accumulation of total aroma in petals[8]. In this study, all the rose germplasms were classified into three groups based on the total VOC content. And more, the rose varieties with wide industrial applications were mainly identified in high-aroma group, such as #30 (R. rugosa 'Fenghua'), #27 (R. damascena 'NO.1'), and #24 (R. rugosa 'Hansa'). However, germplasm #11 (R. rugosa 'Kushui'), widely planted in Gansu, China, showed relatively low content of total VOCs, which may be related to planting conditions[6,27].
In the industry, different fragrance phenotypes of floral derivatives result in a wide range of applications, while consumers tend to prefer the high-end derivatives with sweet fragrance, such as perfumes made from R. centifolia and Convallaria keiskei (lily of the valley). Besides the total VOC content, the differentiation of fragrance traits is closely related to the aroma composition, especially the percentage of key characteristic components[28]. Geraniol, citronellol, and β-phenylethanol were found as the majority compounds in rose petals. In many aromatic plants, such as peony and lily, the odors of the above three substances were broadly categorized as rose scent, without further differentiation of their aromatic nuances[8,20,25]. Geraniol and citronellol are regarded as the characteristic components in the petals of Rosaceae plants[26]. However, compared with β-phenylethanol and linalool, geraniol and citronellol lack sweetness[8,22]. In this study, most rose germplasms (27) presented geraniol and citronellol as key characteristic aromatic components, exhibiting floral-geranium scent, floral-green rose scent, and floral-mixed scent, generally lacking sweet notes. Only three germplasms, namely #18, #23, and #27, had relatively high proportions of β-phenylethanol. Unlike geraniol and citronellol, β-phenylethanol imparts a sweet rose scent, blending floral and sweet notes, i.e., floral-sweet rose scent. For example, germplasm #27, with β-phenylethanol as the majority key characteristic component, is usually used for producing essential oil and food additives with a sweet flavor, widely favored by consumers[8,22,29]. Moreover, the odor analysis could simultaneously integrate the two most important indicators of fragrance trait, namely the concentration and odor threshold of each aromatic substance, and could identify important trace substances with relatively low threshold, such as linalool. The odor threshold of linalool (0.0015 μg·g−1) is much lower than that of other key components, making it easier to be discerned at low content levels[8]. And the note of linalool is described as a sweet aroma with a strong aroma of lily of the valley, which could significantly enhance the percentage of sweet traits in the whole petal fragrance. For instance, in germplasms #14 and #20, linalool accounted for 8.36% and 3.88% of the total content of the four key components, yet its OAV contribution ratios were as high as 45.85% and 31.17%, respectively, resulting in a unique and charming fragrance with sweet characteristics. Rose germplasms with sweet fragrance traits are currently rare. β-Phenylethanol and linalool enhance the percentage of sweet scent in petal fragrance traits[30,31], especially linalool, which as even minor changes in its content could dramatically alter the sweet fragrance phenotype[8]. Thus, these two components can serve as important target compounds for precise improvement of rose fragrance traits, providing a theoretical basis for cultivating rose varieties with unique sweet fragrance traits.
The formation of the sweet-aroma trait is involved in the different expressions of key structural genes
-
The rate and direction of metabolic flow directly affects the biosynthesis and accumulation of aromatic substances[32,33]. In rose, the major aromatic substances were monoterpenoids and benzenoids/phenylpropanoids, which affect the differentiation of fragrance traits as key compounds in petals. The biosynthesis of abundant terpenoids is regulated by a series of TPS members, such as DoTPS10, LiTPS2, PlTPS1, and PlTPS4, which are respectively regarded as the key regulatory genes for linalool and geraniol in ornamental plants[8,20,34]. For rose, the biological functions of some TPS genes have been confirmed through in vitro enzyme activity measurement, and these experiments suggested that the products of TPS members were diverse, including trace monoterpenoids[17,26]. In this study, the correlation analysis and transient overexpression verification suggested that RrTPS1 was the key regulatory gene for linalool biosynthesis and the proportion increase of floral-lily scent in rose petals. In addition to the TPS-dependent pathway, rose species possess a novel terpenoid biosynthesis pathway in which NUDX1 converts GPP to geraniol. This is subsequently converted to citronellal, citronellol, and other substances under the catalysis of several enzymes, such as geranial reductase (GER) and CAD, resulting in geraniol and its derivatives are the predominant terpenoids in rose petals[1,26,35]. In this study, the expression of RrNUDX1-1a had significant positive correlation with monoterpenoid and citronellol content, indicating that RrNUDX1-1a may influence monoterpenoid accumulation by regulating the metabolic flux of the geraniol branch. Furthermore, RrCAD1 expression was closely related to citronellol content, suggesting the coordinated regulation of RrNUDX1-1a and RrCAD1 on the distribution of geraniol and citronellol and the differentiation of fragrance traits. There were three branches for the production of phenylacetaldehyde in higher plant, which were catalyzed by a series of enzymes, such as AAAT and PPDC. After that, PAR catalyzes phenylacetaldehyde to form β-phenylethanol as the last and important enzyme[36,37]. In this study, RrAAAT1 and RrPAR2 showed significantly positive correlations with β-phenylethanol content, consisted with a previous study[18]. In summary, different expressions of these structural genes resulted in the diversity of fragrance traits in rose petals by regulating the biosynthesis of key characteristic components.
Transient expression experiment has been used for gene biological function verification[38,39]. However, the functions of aroma-related genes have not been verified in rose petals. In this study, two rose germplasms cultivated worldwide, i.e., #27 (R. damascena) and #30 (R. rugosa 'Fenghua'), were selected for the biological functions of key genes involved in sweet-aroma traits formation. Germplasm #30 is the main edible rose variety cultivated widely in China, but it is restricted in the production of essential oils and fragrances due to its lack of sweetness in petals[4]. It's necessary to enhance sweet fragrance profiles in roses, such as #30, to improve their industrial application potential. β-phenylethanol is an important natural component and provides a sweet scent for flavoring in food and cosmetics[22]. In this study, the transient expression verification showed that the expression of RrAAAT1 and RrPAR2 improved the absolute and relative contents of β-phenylethanol in #30 petals, indicating that these two genes played core roles in regulating the biosynthesis of β-phenylethanol. Moreover, linalool is a key characteristic component with extreme low content but high OAV level in rose petals[8]. The low content of linalool limited the performance of the function verification of RrTPS1 in germplasm #30. In this study, the transient expression of RrTPS1 suggested its key functions in linalool biosynthesis. Therefore, the biological functions of RrTPS1, RrAAAT1, and RrPAR2 on the modification of fragrance traits were performed, resulted in that these genes were potentially key target genes for creating novel rose varieties with sweet scents. Moreover, RrTPS1 enhanced the proportion of the floral-lily scent by promoting the biosynthesis of linalool, a trace aromatic component, while RrAAAT1, and RrPAR2 improved β-phenylethanol content to increase the percentage of floral-sweet rose scent. These two regulatory pathways were verified separately in this study. Subsequently, the coordinated regulation of linalool and β-phenylethanol accumulations can be further performed, that is, to promote the biosynthesis of the two sweet substances and increase the proportion of sweet scent through effectively enhancing the expressions of RrTPS1, RrAAAT1, and RrPAR2 simultaneously. Furthermore, the identification of related transcription factors is crucial for constructing the transcriptional regulatory mechanism on the sweet-aroma trait formation in rose petals[40], which could greatly improve the efficiency of improving the fragrance trait of rose petals.
-
In this study, a total of 32 rose germplasms were selected for the VOC profile analysis. Monoterpenoids and benzenoids/phenylpropanoids were the majority VOC categories with great variation among different germplasms. Geraniol, citronellol, β-phenylethanol, and linalool were identified as the key characteristic components in rose petals, among which β-phenylethanol and linalool were regarded as target compounds for sweet-aroma trait. The scents of all the 32 rose germplasms were divided into five types, i.e., floral-geranium scent, floral-green rose scent, floral-sweet rose scent, floral-lily scent, and floral-mixed scent. Five key structural genes, namely RrNUDX1-1a, RrCAD1, RrTPS1, RrAAAT1, and RrPAR2, may affect the differentiation of fragrance formation as key candidate genes. And RrTPS1, RrAAAT1, and RrPAR2 regulated the biosynthesis of linalool and β-phenylethanol, respectively, which were regarded as target genes for creating novel rose varieties with sweet scent. These findings revealed the diversity and differentiation mechanism of fragrance traits in rose petals, which could provide important gene resources for the precise improvement of fragrance traits and the cultivation of novel varieties with sweet aroma.
This work was funded by the Key R&D Program of Shandong Province, China (Grant No. 2024LZGC025), and the Shandong Province Natural Science Foundation (Grant Nos ZR2023MC015; ZR2024MC116). We would like to acknowledge Mr. Bo Wang from Xinyao Rose Technology Co., Ltd. for providing the assistance in identifying different rose germplasms.
-
The authors confirm contribution to the paper as follows: study conception and design: Yu Y, Lu J, Wu Q, Xing S; data collection: Li X, Wang J, Lu X, Meng X; analysis and interpretation of results: Li X, Wu Q, Wang C, Xing G, Han G, Sheng S; draft manuscript preparation: Li X, Lu J, Wu Q, Yu Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xiaotong Li, Qikui Wu
- Supplementary Table S1 The list of rose germplasms used in this study.
- Supplementary Table S2 All the primer sequences used in experiments.
- Supplementary Table S3 The contents (μg g−1 FW) of identified VOCs in the petals of different rose germplasms.
- Supplementary Table S4 The content (μg g−1 FW) and percentage (%) of different VOC categories in the petals of different rose germplasms.
- Supplementary Table S5 The OAV of different aromatic substances in the petals of different rose germplasms.
- Supplementary Fig. S1 The PCA (a) and HCA (b) results of aromatic metabolome in the petals of different rose germplasms.
- Supplementary Fig. S2 The changes of gene expression and aromatic compound content after single overexpression of RrAAAT1 (a) and RrPAR2 (b) in rose petals.
- Supplementary Fig. S3 The changes of gene expression and aromatic compound content after silent expression of RrTPS1 (a), RrAAAT1 (b) and RrPAR2 (c) in rose petals.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li X, Wu Q, Wang J, Wang C, Lu X, et al. 2025. Analysis of volatile organic compound diversity revealed the differentiation of scent traits and candidate genes for sweet-aroma formation rose petals. Ornamental Plant Research 5: e042 doi: 10.48130/opr-0025-0040 

Analysis of volatile organic compound diversity revealed the differentiation of scent traits and candidate genes for sweet-aroma formation rose petals
- Received: 06 June 2025
- Revised: 09 September 2025
- Accepted: 13 October 2025
- Published online: 29 October 2025
Abstract: Roses exhibit significant potential as important ornamental plants and industrial crops with petals having high volatile organic compound (VOC) content, diverse aromatic profiles, and pleasant fragrance. The fragrance traits of petals, especially the sweet-aroma, are closely related to the evaluation of petal-derived products, and the development of novel cultivars. However, the scent diversity and key regulatory gene expression among different germplasms remain unclear. In this study, the petal VOC profiles of 32 rose varieties were analyzed. A total of 80 VOCs were detected, and the total VOC content ranged from 6.16 ± 0.03 to 28.26 ± 0.18 μg·g−1 FW, with an average of 13.80 μg·g−1 FW. Monoterpenoids and benzenoids/phenylpropanoids accounted for 93.53% of the total VOC content. Based on the content and odor activity value of each compound, geraniol, citronellol, β-phenylethanol, and linalool were identified as key characteristic components. The scents of all 32 germplasms were classified into five categories: floral-geranium, floral-green rose, floral-sweet rose, floral-lily, and floral-mixed. β-Phenylethanol and linalool were regarded as the target compounds for the characteristic sweet-aroma in floral-sweet rose and floral-lily scented germplasms. Gene expression analysis suggested that RrNUDX1-1a, RrCAD1, RrTPS1, RrAAAT1, and RrPAR2 were related to the differentiation of petal fragrances. Transient overexpression and silencing expression experiments verified that RrTPS1, RrAAAT1, and RrPAR2 regulated the biosynthesis of β-phenylethanol and linalool, emerging as potential targets for breeding sweet-scented cultivars. These findings explored the diversity of VOC in rose, providing both theoretical foundations and gene resources for rose floral characteristics improvement and novel cultivar development.
-
Key words:
- Rose /
- Volatile organic compound /
- Fragrance trait /
- Sweet aroma