









-
Global climate change has led to an increase in extreme heat events, posing significant threats to the quality traits and economic value of ornamental plants[1,2]. As a globally important ornamental species, rose (Rosa spp.) exhibits marked susceptibility to heat stress. Sustained high-temperature conditions reduce its ornamental and economic value by decreasing antioxidant enzyme activities, elevating malondialdehyde (MDA) content, damaging the chloroplasts' ultrastructure, and suppressing anthocyanin accumulation[3,4]. Consequently, elucidating the molecular regulatory mechanisms of heat tolerance in rose holds both theoretical and practical significance.
Current research has identified that the molecular mechanisms underlying plant abiotic stress tolerance involve multiple regulatory networks. The trehalose metabolism pathway plays a crucial role in plant abiotic stress resistance through its involvement in energy metabolism and signal transduction. Trehalose, a nonreducing disaccharide composed of two α-glucose molecules linked by an α-1,1-glycosidic bond, is found in bacteria, yeast, plants, and various other organisms[5,6]. Trehalose biosynthesis in plants comprises two steps: first, trehalose-6-phosphate synthase (TPS, EC 2.4.1.15) catalyzes the synthesis of trehalose-6-phosphate (T6P) from uridine diphosphate glucose and glucose-6-phosphate; subsequently, trehalose-6-phosphate phosphatase (TPP, EC 3.1.3.12) dephosphorylates T6P to yield trehalose[7,8].
Owing to its unique physical and chemical properties, such as the lack of reducing ends related to the formation of glycosidic bonds, trehalose can stabilize enzymes, proteins, and lipids within cellular membranes under heat stress or water-limited conditions[6,9]. Moreover, numerous studies have shown that exogenous trehalose treatment can improve the heat tolerance of plants. For example, research has indicated that exogenous trehalose application safeguarded chloroplasts' integrity in Paeonia lactiflora under heat stress[10]. Furthermore, exogenous trehalose treatment mitigated membrane lipid peroxidation and restored mycelial growth in Pleurotus ostreatus under heat stress[11]. Additionally, exogenous trehalose treatment can facilitate adventitious root development in cucumber under drought stress through the coordinated regulation of reactive oxygen species (ROS), carbohydrate metabolism, and phytohormone pathways[12].
Furthermore, T6P, an intermediate product of trehalose's biosynthetic pathway, serves as an important signaling molecule in glucose metabolism and signal transduction under abiotic plant stress[13]. For instance, in Arabidopsis thaliana, T6P integrates temperature signaling with endogenous sugar distribution via the SNF1 KINASE HOMOLOG 10 (KIN10)–PHYTOCHROME INTERACTING FACTOR 4 (PIF4) module to regulate hypocotyl elongation[14]. Trehalose biosynthesis is governed by the TPS gene family, which is phylogenetically divided into the Class I and Class II subfamilies on the basis of their conserved protein domains. The size of the TPS gene family varies across species; for example, A. thaliana contains 11 AtTPS genes (AtTPS1–AtTPS11)[15], Oryza sativa has 11 OsTPS genes[16], Solanum tuberosum has 8 StTPS genes[17], Cucumis sativus has 7 CsTPS genes[18], and Phaseolus vulgaris has 12 PvTPS genes[19]. Class I members (e.g., AtTPS1–AtTPS4 in A. thaliana and their orthologs in other species) possess a conserved N-terminal TPS-like domain and a less conserved C-terminal TPP domain, primarily exhibiting catalytic activity. In contrast, Class II members (e.g., AtTPS5–AtTPS11 in A. thaliana) retain dual domains but with a highly conserved C-terminal TPP domain that typically plays a role in abiotic stress resistance in plants[8,20]. Notably, Class II TPS genes play pivotal roles in abiotic stress adaptation across different species. For example, TaTPS11 from Triticum aestivum enhances the cold tolerance of A. thaliana by regulating carbohydrate metabolism[21]. GhTPS11 in cotton (Gossypium hirsutum) improves drought and salt tolerance by promoting trehalose accumulation[22]. Studies have proposed that the ClTPS3 gene of watermelon (Citrullus lanatus) is regulated by myeloblastosis (MYB) and basic helix-loop-helix (bHLH) transcription factors to enhance salt tolerance[23].
Our previous research identified nine RcTPS genes in R. chinensis 'Slater's Crimson China', including RcTPS7b, a Class II TPS member closely related to AtTPS7 in A. thaliana. RcTPS7b displayed significantly higher transcription levels in roots, stems, and leaves under heat stress compared with other family members, and trehalose content was also elevated in roots and stems under these conditions[24]. Furthermore, multiple stress-related cis-acting elements were identified in the promoter region of RcTPS7b, including abscisic acid response element (ABRE) motifs that are responsive to abscisic acid (ABA), suggesting potential regulation by environmental signals. To further investigate the regulatory effects of exogenous trehalose and RcTPS7b on the heat tolerance of R. chinensis, we cloned RcTPS7b from R. chinensis 'Slater's Crismson China'. We then compared the phylogenetic relationship and protein structure of RcTPS7b with other TPS7 orthologs from diverse plant species. Our experiments demonstrated that exogenous trehalose enhanced the heat tolerance of R. chinensis and upregulated the expression level of RcTPS7b. Moreover, overexpression of RcTPS7b conferred increased heat tolerance in both A. thaliana and R. chinensis, whereas silencing RcTPS7b reduced heat tolerance in R. chinensis. These results help elucidate the roles of trehalose and RcTPS7b in regulating heat stress responses in R. chinensis and provide a foundation for further mechanistic research.
-
Stock plants of R. chinensis 'Slater's Crimson China' were maintained in the ornamental plant germplasm center at Hunan Agricultural University, Changsha, China. One-year-old healthy cuttings were cultivated in plastic pots filled with a growth substrate mixture of coconut coir, peat moss, and perlite (4:3:3, v/v/v) under controlled environmental conditions: a photon flux density of 17 µmol/m2/s, 65% relative humidity, a temperature regime of 25/20 °C (light/dark), and a 16-h/8-h photoperiod.
A. thaliana (Columbia subtype) and Nicotiana benthamiana seeds were germinated on Murashige and Skoog (MS) basal medium (Coolaber, Beijing, China; Cat. No. PM10111). Two-week-old seedlings were transplanted into plastic pots and cultivated under the following conditions: a photon flux density of 17 µmol/m2/s, 65% relative humidity, a temperature regime of 23/20 °C (light/dark), and a 16-h/8-h photoperiod.
Freshly cut flowers of R. chinensis 'Gaoyuanhong' were purchased from the Hongxing Flower Market in Changsha, Hunan Province, China, for the transient overexpression experiments.
Phylogenetic and bioinformatic analysis
-
Total RNA was isolated from mature leaves of R. chinensis 'Slater's Crimson China' using a plant RNA extraction kit (Accurate Biotechnology, Cat. No. AG21017). Complementary DNA (cDNA) was synthesized with the HiScript II First-Strand cDNA Synthesis Kit (Vazyme, Nanjing, China; Cat. No. R211). Based on the TPS7 reference sequence (GenBank: NC_037092) of R. chinensis 'Old Blush' from the National Centre for Biotechnology Information (NCBI) database (
www.ncbi.nlm.nih.gov ), the full-length coding sequence (CDS) of RcTPS7b in R. chinensis 'Slater's Crimson China' was amplified using the gene-specific primers shown in Supplementary Table S1. Homologous TPS7 amino acid sequences from 13 species were retrieved from the NCBI database to construct a phylogenetic tree (accession numbers are provided in Supplementary Table S2). A neighbor-joining tree was constructed using MEGA 11.0 with the following parameters: JTT+G substitution model, pairwise deletion, and 1,000 bootstrap replicates.Protein domains were predicted using the SMART database (
http://smart.embl-heidelberg.de ). Proteins' physicochemical properties, including molecular mass, instability index, and hydrophilicity, were determined using ExPASy-ProtParam (https://web.expasy.org/protparam/ ).Tissue-specific expression analysis
-
Total RNA was extracted from the following tissues of R. chinensis 'Slater's Crimson China': lateral roots (5-cm segments from 1-year-old plants), young stems (second internode from the apex), mature leaves (third fully expanded leaf from the shoot tip), and petals (dissected from fully opened flowers, excluding sepals, stamens, and pistils). Reverse transcription quantitative polymerase chain reaction (RT-qPCR) was performed using SYBR Green Master Mix in 20-µL reactions on a Bio-Rad CFX Connect™ Real-Time PCR Detection System (Bio-Rad, CA, USA) to quantify RcTPS7b expression levels. The relative expression levels of RcTPS7b were normalized to RcCTP and calculated using the 2−ΔΔCᴛ method[24]. All primers used are listed in Supplementary Table S1.
Exogenous trehalose treatment
-
Rooting cutting seedlings of R. chinensis 'Slater's Crimson China' with a uniform culture duration and growth phenotype were selected. Leaves were sprayed with 0, 2, 5, 10, or 20 mmol/L of trehalose (Macklin, Shanghai, China; Cat. No. D807342) at a fixed time for 3 d, followed by heat stress treatment at 45 °C for 36 h. Thirty plants per concentration were treated across three biological replicates.
After heat stress, the heat injury phenotypes of the plants were observed and graded as follows[25]: 0. No injury; 1. Mildly dehydrated and yellowing of 1–5 leaves; 2. Mildly dehydrated, yellowing of 5–10 leaves, and death of 1–3 leaves from dehydration; 3. Severe dehydration and yellowing of >10 leaves; 4. Whole-plant dehydration and wilting. The heat injury index was calculated as:
$ \rm{H}eat\; injury\; index=\dfrac{\text{∑}\; (\rm{X}\times\rm{N}_{\rm{x}})}{\text{A × N}} $ where, X is the injury level (0–4), Nₓ is the number of plants at level X, A is the highest injury level (4), and N is the total number of plants surveyed.
To assess the effect of exogenous trehalose on the transcription of RcTPS7b during heat stress, mature leaves were collected from plants treated with 0 and 5 mmol/L trehalose after 0, 1, and 2 h of heat stress. All samples were collected from three biological replicates and immediately frozen in liquid nitrogen and stored at –80 °C for subsequent RT-qPCR analysis.
All physiological indices were measured uniformly at the end of 36 h of heat stress. In addition, mature leaves (0.1 g per sample, three biological replicates) were collected at 36 h for the superoxide dismutase (SOD) activity assay (Solarbio, Beijing, China; Cat. No. BC0170). The maximum quantum yield of Photosystem II (PSII) (Fv/Fm) was determined after 36 h using a chlorophyll fluorescence imaging system (Pen FP110, FluorCam, Czech Republic; three biological replicates, each with three technical replicates).
Subcellular localization
-
The pBI121-GFP empty vector and pBI121-RcTPS7b-GFP recombinant plasmid were introduced into Agrobacterium tumefaciens strain GV3101 (optical density at 600 nm (OD600) = 0.6–0.8). Bacterial suspensions (OD600 = 0.6) in an infiltration buffer (10 mmol/L MgCl2, 200 µmol/L acetosyringone, and 10 mmol/L 2-morpholinoethanesulphonic acid [MES]; pH 5.6) were infiltrated into the abaxial side of 4–5-week-old N. benthamiana leaves using a 1-mL sterile syringe. A plasma membrane marker (DsRed) was co-infiltrated as a control. After 48 h of incubation in the dark, green fluorescent protein (GFP) and membrane-localized DsRed signals were visualized using a fluorescence microscope (CarlZeiss LSM710, Oberkochen, Germany).
Genetic transformation of A. thaliana
-
The RcTPS7b CDS was cloned into the pCAMBIA1305 vector (CaMV35S promoter) to generate the 35S::RcTPS7b overexpression construct. The pCAMBIA1305-RcTPS7b bacterial culture was enriched by centrifugation, and the pellet was reconstituted in a 5% (w/v) sucrose solution. Genetic transformation of A. thaliana was subsequently performed using the floral dip method. In total, 15 A. thaliana plants were transformed, yielding 10 T1 transgenic plants. T1 lines showing relatively high RcTPS7b expression levels by RT-qPCR were advanced to the T3 generation. Homozygous T3 lines were selected on a medium containing 30 mg/L of hygromycin (Solarbio, Beijing, China; Cat. No. H8070).
To compare heat tolerance between transgenic and wild-type (WT) plants, WT and three independent homozygous T3 lines (OE-2, OE-3, and OE-4) at different growth days were subjected to heat stress, the phenotypes of heat damage were observed, and the enzyme activity and transcription level of related genes were measured.
Seven-day-old seedlings were cultured on MS medium plates (30 seedlings per line per plate). Three plates constituted one biological replicate, with three independent biological replicates performed (totaling nine plates). Seedlings were exposed to 45 °C heat stress for 90 min, followed by a 7-day recovery period, after which the heat damage phenotypes and transcript levels of target genes were assessed.
Twenty-day-old plants were transplanted into individual pots containing three WT plants and three plants of a single transgenic line per pot. For each transgenic line, five pots constituted one biological replicate, with three independent biological replicates established (totaling 15 pots per line). Plants were subjected to 45 °C heat stress for 12 h, and leaves were collected for physiological assays. Following 12 h of heat stress, SOD activity, catalase (CAT) activity (Solarbio, Cat. No. BC4780), MDA (Boxbio, Cat. No. AKFA013C), and H2O2 content (Boxbio, Cat. No. AKAO009C) were measured in leaf samples (0.1 g per sample, three biological replicates) from WT and transgenic lines.
Thirty-day-old plants (10 per line × 3 replicates) were exposed to 45 °C heat stress for 36 h, and heat damage phenotypes were recorded. Subsequently, the chlorophyll content was determined. Fresh leaves (0.1 g per sample, pooled within replicates) were collected and weighed. Samples were flash-frozen in liquid nitrogen and ground to a fine powder. Subsequently, 10 mL of an extraction reagent (acetone : ethanol : water = 9:9:2, v/v/v) was added, and the mixture was incubated in darkness for 24 h until complete tissue decolorization (n = 3 biological replicates). Absorbance was measured at 665 nm and 649 nm using an ultraviolet–visible spectrophotometer (UV-4802, Unico, Shanghai, China). Chlorophyll a and b contents were calculated using Arnon's equations[26].
Total RNA was extracted from 7-day-old WT and homozygous T3 seedlings (OE-2, OE-3, OE-4). Expression levels of the transgene RcTPS7b, the endogenous gene AtTPS7 (AT1G06410) and key heat-responsive genes, namely HEAT SHOCK FACTOR A1a (AtHSFA1a, AT4G17750), HEAT SHOCK FACTOR A3 (AtHSFA3, AT5G03720), DEHYDRATION-RESPONSIVE ELEMENT BINDING PROTEIN 2A (AtDREB2A, AT2G40340), MULTIPROTEIN BRIDGING FACTOR 1C (AtMBF1c, AT3G24500), and HEAT SHOCK PROTEIN 70 (AtHSP70, AT3G12580), were quantified using AtActin2 (AT3G18780) as the reference gene[27].
Transformation of R. chinensis
-
Virus-induced gene silencing (VIGS) of RcTPS7b was performed according to a previously described protocol with modifications[28]. A 350-bp fragment of RcTPS7b was amplified and cloned into the pTRV2 vector to generate pTRV2-RcTPS7b. The pTRV1, pTRV2, and pTRV2-RcTPS7b plasmids were introduced into Agrobacterium tumefaciens GV3101 (OD600 = 1.3–1.4). Equal volumes (OD600 = 1.0) of pTRV1 + pTRV2 or pTRV1 + pTRV2-RcTPS7b suspensions were mixed and incubated at room temperature for 4 h. The rooted cuttings of R. chinensis 'Slater's Crimson China' were vacuum-infiltrated (0.7 MPa, 10 min × 2 cycles). In total, 30 seedlings were infiltrated with the pTRV2 empty vector, and 45 with pTRV2-RcTPS7b. Treated plants were maintained for 45 d until the emergence of new leaves. Silencing efficiency and the expression of RcHSF1 (a heat-responsive marker) were confirmed by RT-qPCR in newly emerged leaves. Fourteen RcTPS7b-silenced plants were identified by RT-PCR screening, representing a silencing efficiency of approximately 46.7%. Twelve RcTPS7b-silenced plants and twelve empty vector controls (four plants per biological replicate, three replicates) were exposed to 45 °C for 10 h. Phenotypes were documented, and physiological parameters (SOD/CAT activity, MDA/H2O2 content) were analyzed.
Transient overexpression of RcTPS7b in R. chinensis 'Gaoyuanhong' petals was performed according to the methods of Wu[29] and Geng[30], with minor modifications. The 35S::RcTPS7b recombinant plasmid and pCAMBIA1305 empty vector were introduced into Agrobacterium tumefaciens GV3101 (OD600 = 1.0–1.2). Bacterial suspensions were prepared in an infiltration buffer (10 mmol/L MgCl2, 200 µmol/L acetosyringone, 10 mmol/L MES; pH 5.6). Healthy petals were selected, and discs 1 cm in diameter were excised from the adaxial side using a sterile cork borer. Discs (25 per dish, 3 dishes per treatment) were immersed in suspensions (35S::RcTPS7b or pCAMBIA1305 empty vector) and vacuum-infiltrated (0.7 MPa, 10 min × 2 cycles). Three biological replicates (nine dishes in total per treatment) were performed. After 96 h of incubation in the dark at 22 °C, RcTPS7b (overexpression validation) and RcHSF1 expression levels were analyzed by RT-qPCR using RNA from a subset of discs before heat stress. Subsequently, infiltrated petals discs were exposed to 45 °C heat stress for 10 h. Phenotypic damage (wilting and color fading) was recorded, and physiological parameters were assessed as previously described.
Statistical analysis
-
Experimental data were processed in Microsoft Excel 2016 and analyzed using IBM SPSS Statistics 20. Multi-group comparisons were evaluated by one-way analysis of variance (ANOVA) with Duncan's multiple range test (p < 0.05), with homogeneous subsets denoted by lowercase letters (a > b > c). Pairwise comparisons used Student's t-test, with significance denoted as ND (no difference), * p < 0.05, or ** p < 0.01. Graphs were generated in Origin 2022, presenting the data as mean ± standard deviation (SD) of three biological replicates.
-
The full-length CDS of RcTPS7b (2,565 bp, encoding 854 aa) was amplified from the leaves of R. chinensis 'Slater's Crimson China' (Fig. 1a). Alignment with the TPS7 ortholog in R. chinensis 'Old Blush' revealed eight nucleotide substitutions (Positions 39, 147, 636, 1,071, 1,288, 1,677, 1,993, and 2,481). Bioinformatic analysis predicted RcTPS7b to be an unstable hydrophilic protein with the molecular formula C4297H6707N1165O1273S39, a molecular mass of 96.31 kDa, an aliphatic index of 85.81, a theoretical pI of 5.93, an instability index of 46.67, and a grand average of hydropathicity of –0.266. Conserved protein domain analysis showed both the characteristic GT20-TPS and trehalose_PPase domains in the amino acid sequence (Fig. 1b).
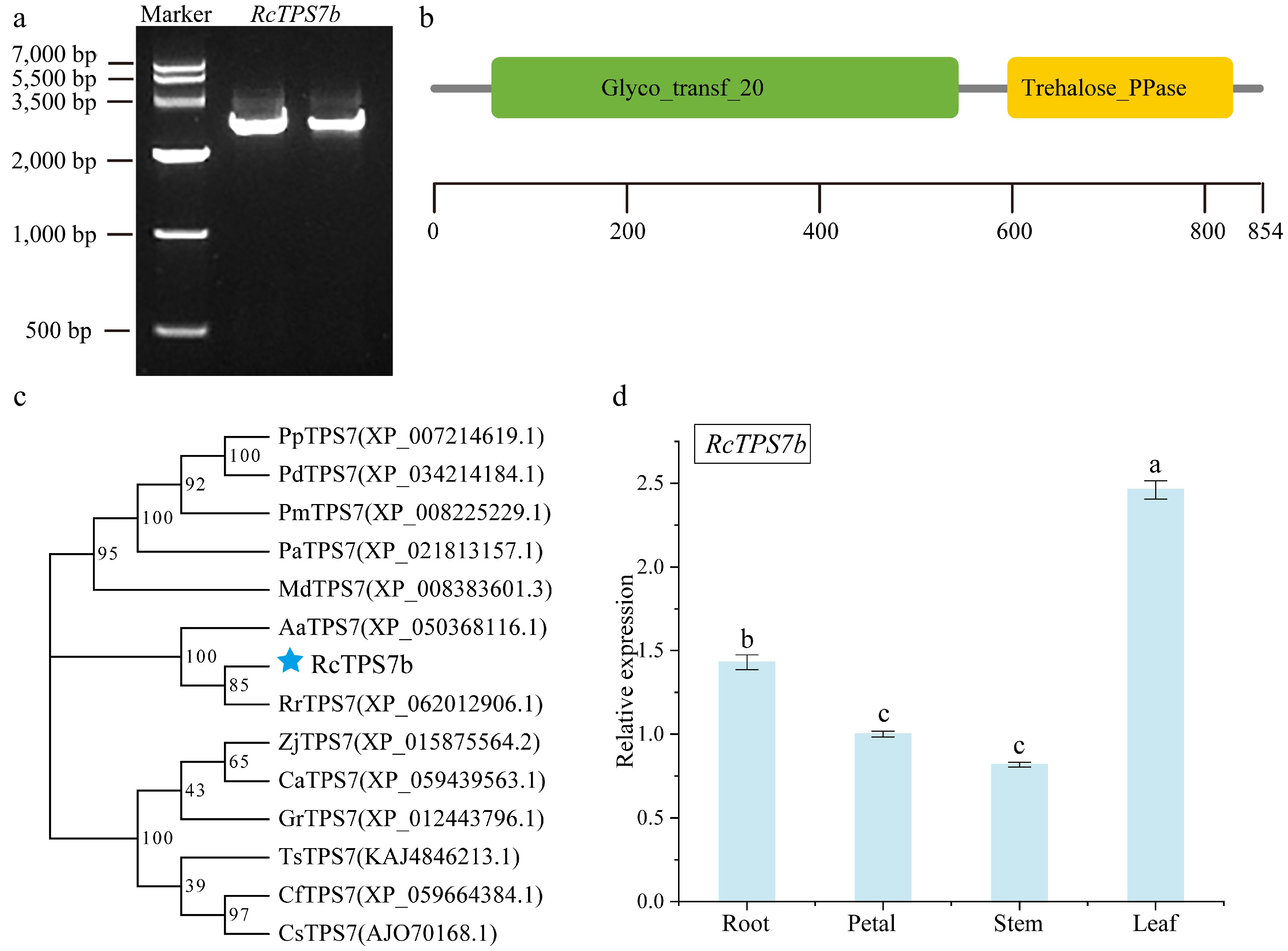
Figure 1.
Characterization and phylogenetic analysis of RcTPS7b. (a) PCR amplification of RcTPS7b. (b) Conserved protein domains analysis of RcTPS7b. The green box represents the Glyco_transf_20 (TPS) domain, and the yellow box represents the trehalose_PPase (TPP) domain. (c) Phylogenetic tree of TPS7 orthologs. Pp, Prunus persica; Pd, Prunus dulcis; Pm, Prunus mume; Pa, Prunus avium; Md, Malus domestica; Aa, Argentina anserina; Rr, Rosa rugosa; Zj, Ziziphus jujuba; Ca, Corylus avellana; Gr, Gossypium raimondii; TS, Turnera subulata; Cf, Cornus florida; Cs, Camellia sinensis. Bootstrap values are shown at the nodes. The blue star indicates RcTPS7b. (d) Tissue-specific expression of RcTPS7b in roots, stems, leaves, and petals (mean ± SD; ANOVA; a > b > c labels; p < 0.05).
Based on the phylogenetic classification of RcTPS7b as a Class II TPS member closely related to AtTPS7[24], a phylogenetic tree was constructed using the TPS7 orthologous amino acid sequence identified from 13 species including Rosa rugosa, Argentina anserina, Prunus mume, and so on. The results showed that RcTPS7b clustered most closely with RrTPS7 from R. rugosa (Fig. 1c), reflecting their closely phylogenetic relationship within the Rosa genus.
Tissue-specific expression analysis revealed RcTPS7b transcript levels were highest in the leaves (2.46-fold relative to the reference tissue), significantly exceeding other tissues. The transcription accumulation of RcTPS7b in roots (1.43-fold) was second, followed by petals (1.0-fold), and the lowest in stems (0.82-fold) (Fig. 1d).
Exogenous trehalose treatment enhances heat tolerance in R. chinensis
-
In order to evaluate the regulatory effect of exogenous trehalose treatment on R. chinensis 'Slater's Crimson China' under heat stress, we systematically analyzed heat damage phenotypes, physiological responses, and RcTPS7b expression after 36 h of heat stress. The results showed that treatment with 5 mmol/L trehalose effectively alleviated leaf curling and dehydration, significantly reduced the heat damage index, and improved antioxidant capacity and photosynthetic performance (Fig. 2a, b). SOD activity of the plants under the 5 mmol/L trehalose treatment increased to 114.9 ± 12.8 U/g FW (68.9% higher than the controls; p < 0.05), indicating efficient ROS scavenging (Fig. 2c). Meanwhile, the maximum quantum efficiency of PSII (Fv/Fm) remained at 0.64 ± 0.03 under 5 mmol/L trehalose, significantly higher than the control values (p < 0.05). Moreover, the Fv/Fo ratios under the 5 mmol/L trehalose treatment were also significantly higher than in the control, suggesting activated nonphotochemical quenching (Fig. 2d). Therefore, these results indicated that the heat tolerance of R. chinensis could be improved by applying 5 mmol/L trehalose.
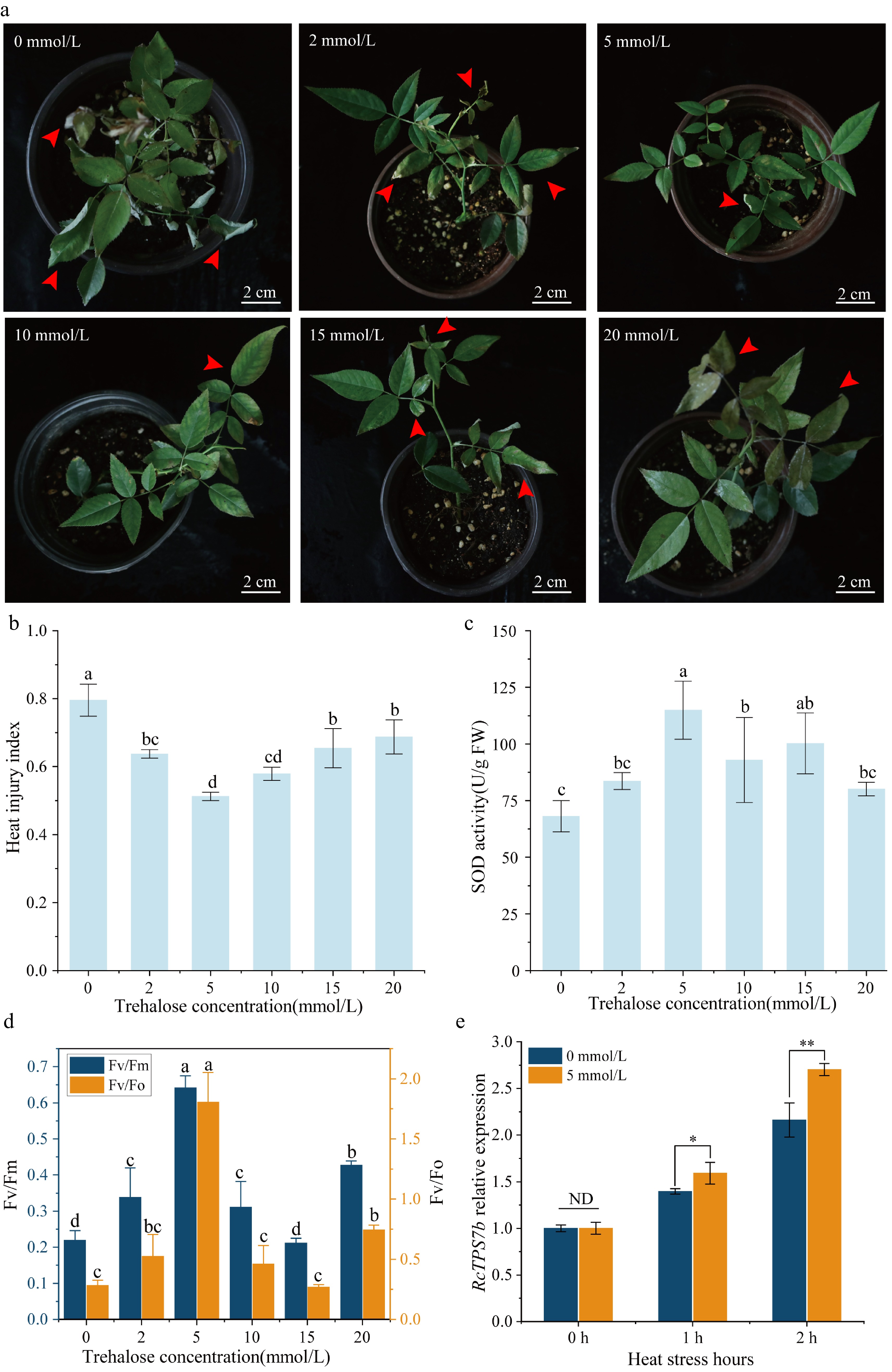
Figure 2.
Exogenous trehalose treatment improved the heat tolerance of R. chinensis 'Slater's Crimson China'. (a) Phenotypic analysis of R. chinensis 'Slater's Crimson China' treated with 0–20 mmol/L trehalose under 45 °C for 36 h. (b) Heat injury index analysis. (c) SOD activity analysis. (d) Parameter analysis of the photosynthetic system. (e) Transcription analysis of RcTPS7b in R. chinensis plants treated with 0 or 5 mmol/L trehalose under heat stress. Data are presented as the mean ± SD of three biological replicates. In (b)–(d), letters denote significant differences (ANOVA; a > b > c >d; p < 0.05). In (e), * p < 0.05 and ** p < 0.01 (Student's t-test; ND, no difference).
In addition, heat stress treatment induced the high expression of RcTPS7b in R. chinensis. The expression of RcTPS7b increased gradually with 1–2 h of heat stress. Remarkably, the upregulated expression of RcTPS7b was more pronounced in plants treated with 5 mmol/L trehalose compared with the control. The relative expression of RcTPS7b in the 0 mmol/L trehalose control group increased to 1.40 after 1 h of heat stress, whereas plants treated with 5 mmol/L trehalose showed a 13.6% higher expression level (1.59, p < 0.05). This difference became more marked after 2 h of heat stress, and the relative expression of RcTPS7b in plants treated with 5 mmol/L trehalose being 25.0% higher than that in the control plants (p < 0.01). These results indicated that exogenous trehalose treatment further enhanced the high expression of RcTPS7b under heat stress, thereby improving the heat tolerance of R. chinensis (Fig. 2e).
Subcellular localization of RcTPS7b
-
To determine the subcellular localization of RcTPS7b, we performed transient co-expression assays in N. benthamiana epidermal cells using 35S::RcTPS7b-GFP with the membrane marker 35S::HY5-DsRed. Fluorescence signals from RcTPS7b-GFP exhibited precise overlap with the DsRed-tagged cell membrane marker, while free GFP controls showed a diffuse distribution throughout the cytoplasm and nucleus. These observations confirmed that RcTPS7b was localized to the cell membrane (Fig. 3).
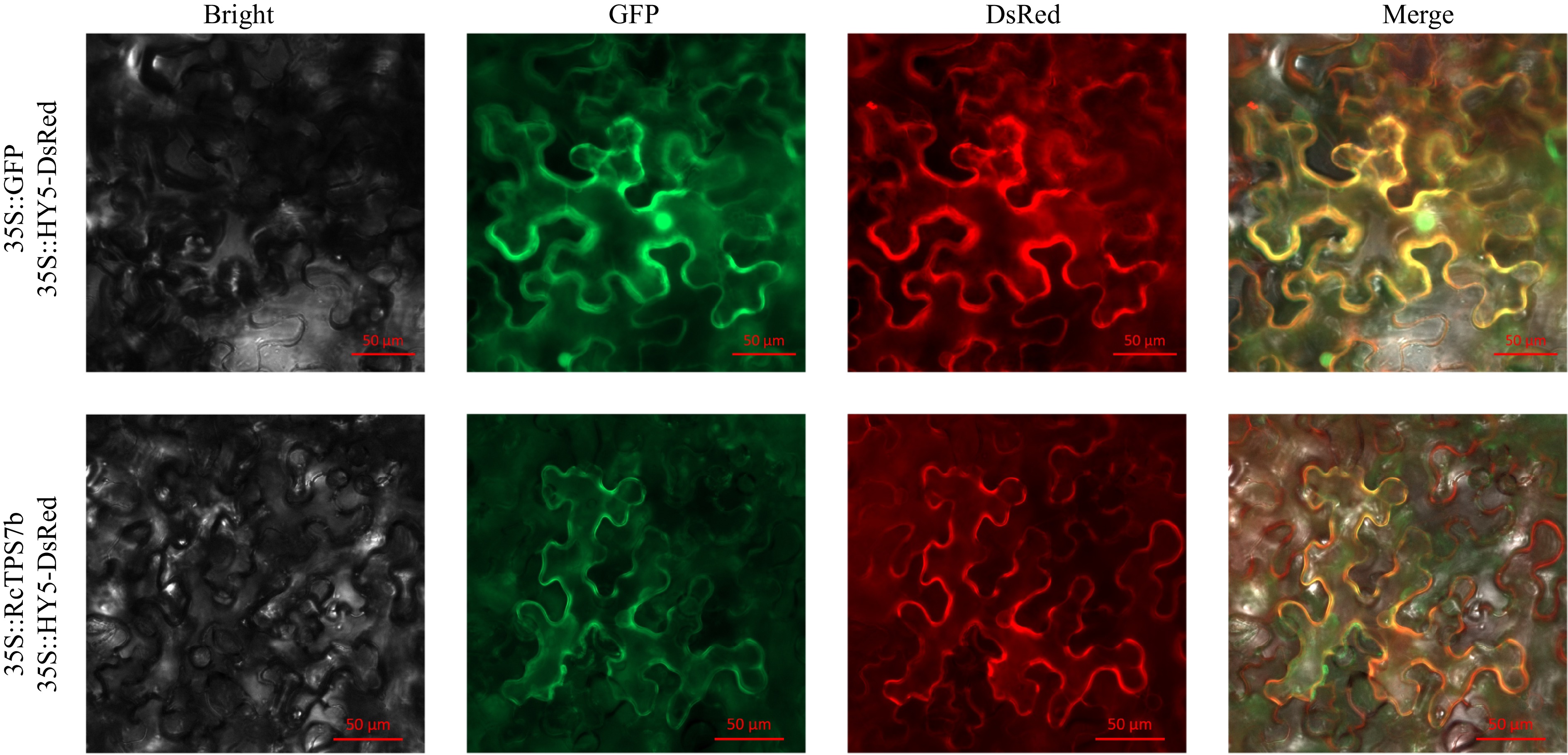
Figure 3.
Subcellular localization of RcTPS7b in tobacco leaves. Bright, brightfield; GFP, GFP fluorescence; DsRed, cell membrane marker fluorescence; Merge, superimposed field.
Overexpression of RcTPS7b enhances heat tolerance in A. thaliana
-
To further verify whether RcTPS7b can enhance heat tolerance, we performed heterologous overexpression of RcTPS7b in A. thaliana. RT-qPCR analysis confirmed that the expression level of the endogenous AtTPS7 gene was not significantly altered in the RcTPS7b overexpression lines compared with the WT plants (Supplementary Fig. S1). We found that heterologous overexpression of RcTPS7b in A. thaliana significantly enhanced the heat tolerance of transgenic plants. After exposure to 45 °C for 90 min followed by a 7-day recovery at 22 °C, the survival rate of 7-day-old seedlings with overexpression of RcTPS7b exceeded 50%, while that of WT plants showed less than 10% survival (Fig. 4a). Moreover, compared with WT plants, the expression of the heat stress-response genes AtHSFA1a, AtHSFA3, AtHSP70, AtDREB2A, and AtMBF1c in the three RcTPS7b-overexpressing lines (OE-2, OE-3, OE-4) showed an upregulated trend after heat stress (Fig. 4b).
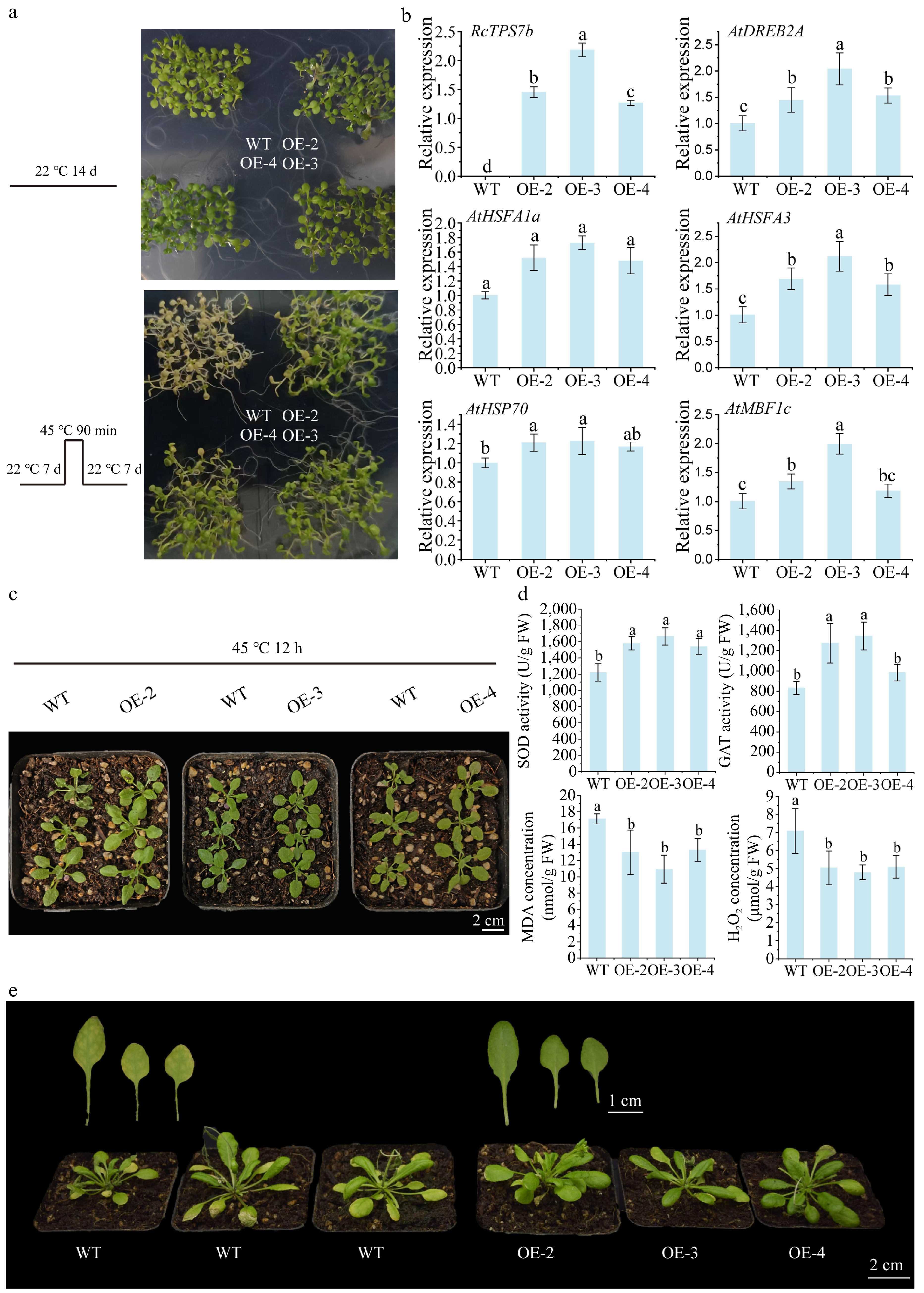
Figure 4.
Overexpression of RcTPS7b improved the heat tolerance of A. thaliana. (a) The phenotypic changes in 7-day-old WT and overexpression (OE) lines were observed at 22 °C for 14 days, at 22 °C for 7 d then at 45 °C for 90 min, and then at 22 °C for 7 d. (b) Transcriptional expression of RcTPS7b and heat stress response genes in 7-day-old WT and OE lines after 90 min of heat stress at 45 °C followed by 7 d of recovery growth at 22 °C (ANOVA; a > b > c > d; p < 0.05). (c) Phenotypic changes in 20-day-old WT and OE lines under heat stress at 45 °C for 12 h. (d) Determination of the content of physiologically active substances in 20-day-old WT and OE lines under 45 °C heat stress for 12 h (ANOVA; a > b; p < 0.05). (e) Phenotypic changes in 30-day-old WT and OE lines under heat stress at 45 °C for 36 h. Data are presented as the mean ± SD of three biological replicates.
After the 20-day-old WT and overexpressing lines were subjected to heat stress for 12 h, the overexpression lines showed slight wilting, while the WT plants showed severe wilting (Fig. 4c). In addition, the average SOD activity of transgenic plants was 31% higher than that of the WT, the average CAT activity was 44% higher than that of the WT, the average MDA activity was 27% lower than that of the WT, and the average H2O2 activity was 30% lower than that of the WT (Fig. 4d).
After the 30-day-old WT and overexpressing lines were treated at 45 °C for 36 h, 80% of the leaves of WT plants turned yellow, but only about 20% of the leaves of overexpression lines turned yellow (Fig. 4e). Consistent with this phenotypic observation, results revealed that the chlorophyll a and chlorophyll b contents in all three RcTPS7b-overexpression lines (OE-2, OE-3, OE-4) were significantly higher than those in WT plants under heat stress (p < 0.05), with increases ranging across approximately 6%–23% for chlorophyll a and 12%–41% for chlorophyll b (Supplementary Fig. S2). These results confirmed that RcTPS7b may play an important role in the regulation of heat tolerance.
Transient silencing of RcTPS7b reduce the heat tolerance of R. chinensis
-
In order to verify the function of RcTPS7b in regulating heat tolerance in R. chinensis, RcTPS7b was chosen for VIGS using a pTRV-based system. Compared with the pTRV2 control plants, RcTPS7b-silenced plants exhibited leaf curl and necrosis phenotypes after heat stress at 45 °C for 16 h (Fig. 5a). The expression of RcTPS7b in virus-induced RcTPS7b-silenced plants was significantly lower than that in pTRV2 control plants (transformed with the pTRV2 vector) (Fig. 5b). RT-qPCR analysis showed that the expression level of the heat stress-responsive transcription factor gene RcHSF1 was significantly reduced in RcTPS7b-silenced plants compared with the pTRV2 controls (p < 0.01; Fig. 5c). Physiological analysis showed that compared with the pTRV2 control plants, the content of H2O2 in RcTPS7b-silenced plants increased by 64.2%, and the content of MDA increased by 16.2% (p < 0.05; Fig. 5d, e). Meanwhile, SOD activity decreased by 29.2%, and CAT activity decreased by 29.4% (p < 0.01; Fig. 5f, g). These data demonstrate that RcTPS7b silencing impairs heat tolerance by downregulating RcHSF1, compromising ROS scavenging capacity and enhancing membrane peroxidation.
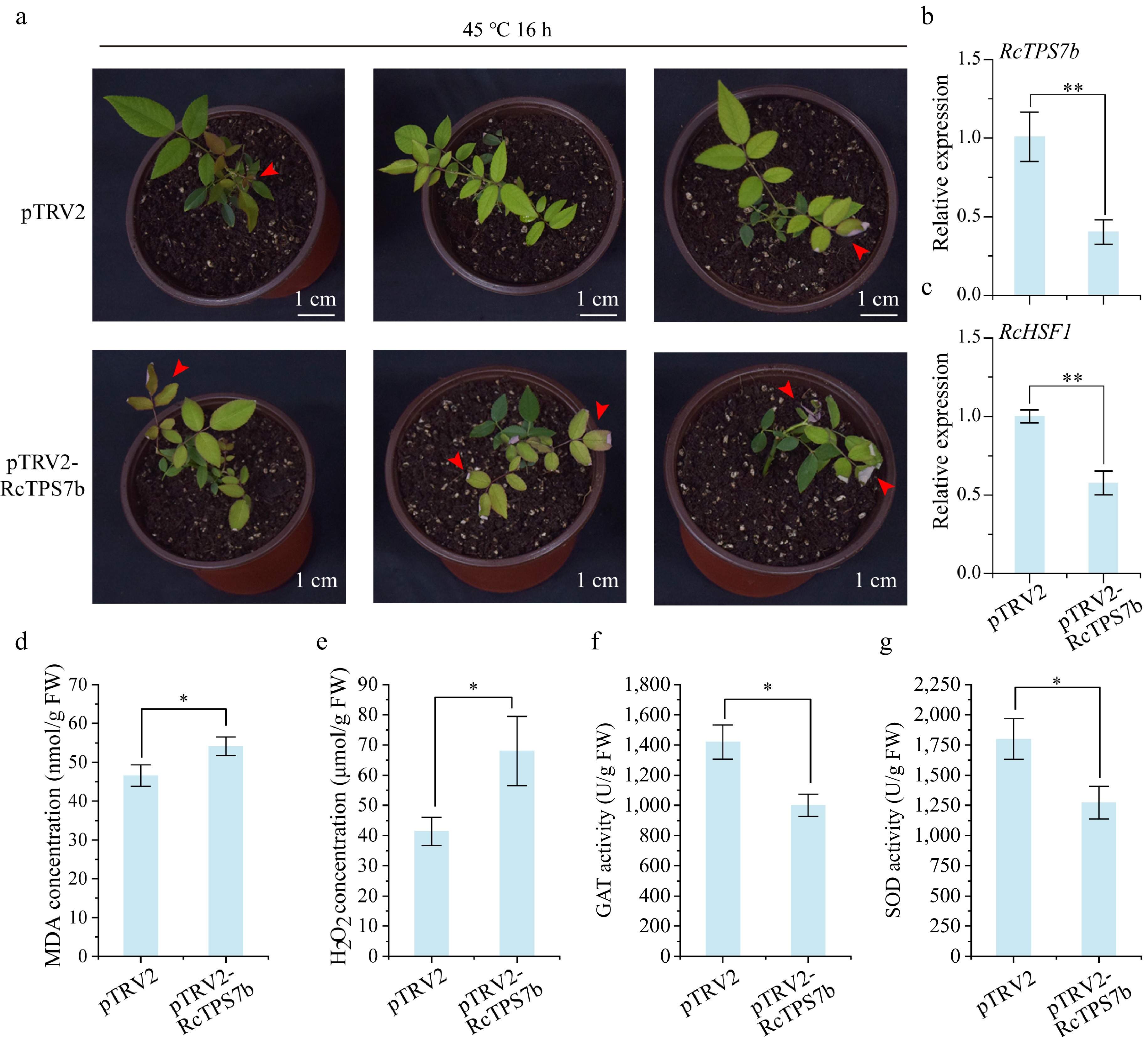
Figure 5.
Transient silencing of RcTPS7b decreased the heat tolerance of R. chinensis plants. (a) Phenotypic changes in RcTPS7b-silenced and control plants after 45 °C heat stress for 16 h. (b), (c) Transcript levels of RcTPS7b and RcHSF1 in VIGS-treated plants. (d)–(g) The content of physiologically active substances in RcTPS7b-silenced and control plants after 45 °C heat stress for 16 h. Data are presented as the mean ± SD of three biological replicates. Statistical significance: * p < 0.05 and ** p < 0.01 (Student's t-test).
Transient overexpression of RcTPS7b enhanced the heat tolerance of rose petals
-
In order to further verify the function of RcTPS7b in heat tolerance regulation in rose, RcTPS7b was transiently overexpressed in rose petals, and the freshness of petals with RcTPS7b overexpression was significantly higher than that of the control after 45 °C for 16 h (Fig. 6a). Moreover, the expression of RcTPS7b was 2.1 times higher that of the control (Fig. 6b). RT-qPCR analysis showed that the expression level of RcHSF1 was significantly increased in RcTPS7b-overexpressed petals compared with the control (p < 0.05; Fig. 6c). Physiological analysis showed that compared with control petals, SOD activity increased by 63.5%, and CAT activity increased by 111% in RcTPS7b-overexpressing petals (p < 0.01; Fig. 6f, g). Meanwhile, the content of H2O2 in RcTPS7b-overexpressing petals decreased by 34.9%, and the content of MDA decreased by 31.9% (p < 0.01; Fig. 6d, e). These results demonstrate that RcTPS7b overexpression confers heat tolerance by upregulating RcHSF1, attenuating oxidative damage, and reducing membrane lipid peroxidation.
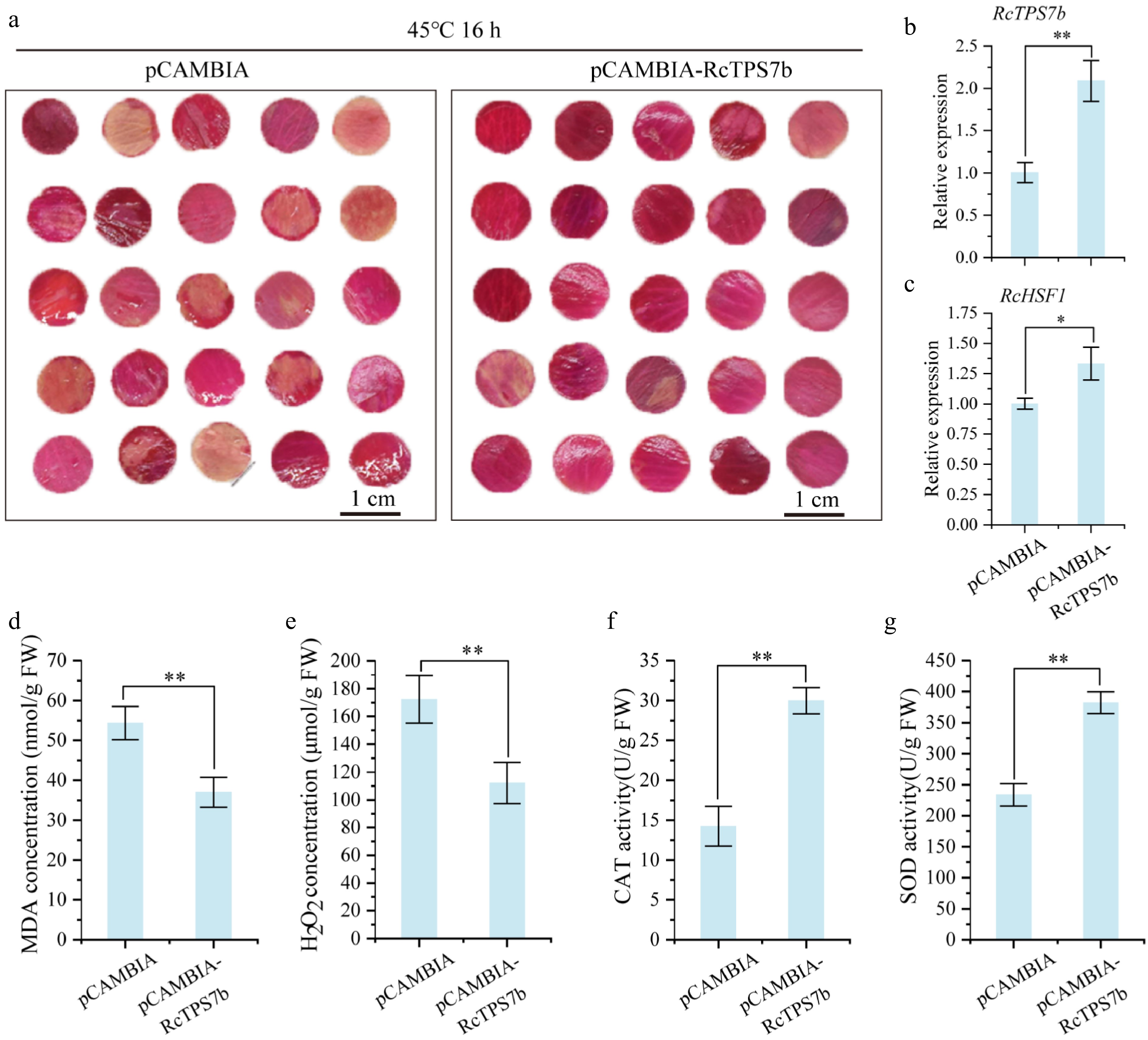
Figure 6.
Transient overexpression of RcTPS7b increased the heat tolerance of rose petals. (a) Phenotypic changes in RcTPS7b-overexpressing and control petals after 45 °C heat stress for 16 h. (b), (c) RcTPS7b/RcHSF1 transcript levels after overexpression. (d)–(g) The content of physiologically active substances in RcTPS7b-overexpressing and control petals after 45 °C heat stress for 16 h. Data are presented as the mean ± SD of three biological replicates. Statistical significance: * p < 0.05 and ** p < 0.01 (Student's t-test).
-
Phylogenetic analysis revealed that RcTPS7b clusters with the TPS7 orthologs of other species (Fig. 1c), indicating the RcTPS7b may share the conserved functions of Class II members of the TPS gene family involved in regulating stress resistance. In this study, we observed that the external application of trehalose enhance the heat tolerance of rose. Under heat stress, exogenous trehalose significantly reduced the heat damage index of rose leaves, increased SOD activity, and mitigated oxidative damage (Fig. 2). This finding aligns with the previously reported mechanism of SOD protection mediated by trehalose in T. aestivum[31]. Moreover, exogenous trehalose may confer protection to heat-stressed plants through a synergistic mechanism involving SOD-mediated ROS scavenging and membrane stabilization[32,33].
Moreover, similar to its effects in wheat, where it significantly scavenges ROS (e.g., 95% H2O2 clearance at 50 mmol/L)[31], exogenous trehalose likely mitigates oxidative stress in rose. Furthermore, exogenous trehalose stabilized PSII complexes (Fv/Fm 0.64 vs 0.22 in controls; Fig. 2d), suggesting that it maintains chloroplast membrane integrity to support electron transport chains' functionality[34]. This stabilization may function via a mechanism analogous to the trehalose-mediated D1 protein (a core component of the PSII reaction center) protection reported in T. aestivum[35]. This protective role of trehalose on the photosynthetic machinery aligns with the findings in wheat, where trehalose pretreatment preserved the chloroplasts' ultrastructure[34]. Notably, the transcriptional levels of RcTPS7b increased under exogenous trehalose treatment during heat stress (Fig. 2e), paralleling the upregulation of SlTPS1, SlTPS5, and SlTPS7 in trehalose-pretreated Solanum lycopersicum seedlings under saline conditions[36]. Critically, our data demonstrate that this transcriptional response is linked to functional protection: transgenic A. thaliana overexpressing RcTPS7b exhibited significantly higher chlorophyll retention under heat stress (Supplementary Fig. S2), which helps explain the observed stabilization of PSII. This cross-species similarity suggests that exogenous trehalose not only provides immediate protection but also enhances the transcription of TPS genes such as RcTPS7b. This phenomenon establishes a unique loop: exogenous trehalose leads to more RcTPS7b expression, potentially creating a metabolic memory that amplifies stress responses.
Overexpression of RcTPS7b enhances heat tolerance and antioxidant capacity in A. thaliana
-
Subcellular localization analysis showed that RcTPS7b localizes at the cell membrane. This localization supports its potential role as a membrane-associated signaling hub, potentially interacting with receptor-like kinases to initiate early stress responses. This hypothesis is corroborated by functional studies of the Class II TPS protein OsTPS8 in rice, which exhibits a similar plasma membrane localization and directly participates in stress signaling through interaction with SAPK kinases (SnRK2 family) to regulate downstream ABA-responsive pathways[37]. Consistent with this signaling role, we detected upregulated AtHSF/P gene expression in transgenic A. thaliana expressing RcTPS7b (Fig. 4b), providing further experimental evidence for its signaling function in stress-responsive transcription.
The increased expression of AtHSFA1a/AtHSP70 (Fig. 4b) and enhanced SOD/CAT activity (Fig. 4d) in RcTPS7b-overexpression lines of A. thaliana indicate that heat tolerance involves both heat-response signaling and antioxidant defense. This is consistent with the known role of HSF genes in controlling heat stress responses, orchestrating the expression of HSPs and other protective genes to mitigate heat-induced damage[38]. While this dual-path mechanism shared functional similarity with TPSP-mediated HSF/P activation in seeds of S. lycopersicum[39]. The conserved HSF/P activation across in A. thaliana and S. lycopersicum systems implies the ancient evolutionary conservation of TPS–HSF/P crosstalk in land plants. However, the causal relationships remain unresolved: whether antioxidant activation operates downstream of HSF signaling or represents a parallel effect of trehalose metabolism requires validation in R. chinensis, as the current evidence is limited to heterologous systems.
Furthermore, membrane-proximal regulation may explain the 25% higher RcTPS7b expression in trehalose-treated plants versus the controls during early heat stress (Fig. 2e), establishing an exogenous replenishment-endogenous potentiation loop analogous to stress – TPS feedback loops in Tamarix hispida and A. thaliana[40,41].
RcTPS7b exhibits unique thermotolerance-regulating functions in R. chinensis
-
We further verified the positive regulation of heat tolerance in rose by transient silencing and overexpression of RcTPS7b. Silencing the RcTPS7b in rose increased oxidative damage in the leaves (Fig. 5), which is consistent with the antioxidant dysfunction observed in CaTPS1-silenced peppers[42]. This supports conserved roles of TPS genes in maintaining redox homeostasis, as demonstrated by how ThTPS enhanced salt/osmotic tolerance in T. hispida through antioxidant modulation and AhTPS9 improved cold adaptation in Arachis hypogaea[40,43]. Transient overexpression of RcTPS7b not only reduced petal H2O2 content by 31.9%, but also significantly elevated SOD/CAT activity and delayed heat-induced petal fading (Fig. 6). These findings reveal functional parallels with the stress-adaptive roles of other TPS genes—though operating through distinct mechanisms—such as OsTPS8 enhancing salt tolerance via ABA-dependent root suberization in rice, and AtTPS9 mediating salt tolerance through endodermal barrier reinforcement in A. thaliana[37,44]. Collectively, these results underscore the functional diversification of TPS genes across plant species and tissues.
RcTPS7b belongs to the Class II TPS proteins, which generally lack direct involvement in trehalose biosynthesis but primarily function in regulatory roles. For example, BnaC02.TPS8 exhibits no catalytic activity yet regulates carbon allocation in rapeseed, demonstrating its role in metabolic regulation[45]. Another member, AtTPS9, controls root development in A. thaliana, showing its function in developmental regulation[44]. On the basis of the family characteristics and homologous protein functions, RcTPS7b is postulated to function as a signaling regulator rather than a biosynthetic enzyme.
While RcTPS7b enhances heat tolerance, TaTPS11 in wheat improves cold adaptation, and AtTPS5 in A. thaliana regulates pathogen defense[21,41]. This functional variation demonstrates how TPS genes adapt to different environmental challenges, exemplified by CkTPS-mediated drought tolerance in Caragana korshinskii[46]. A schematic summary of this proposed trehalose-RcTPS7b regulatory network is presented in Fig. 7.
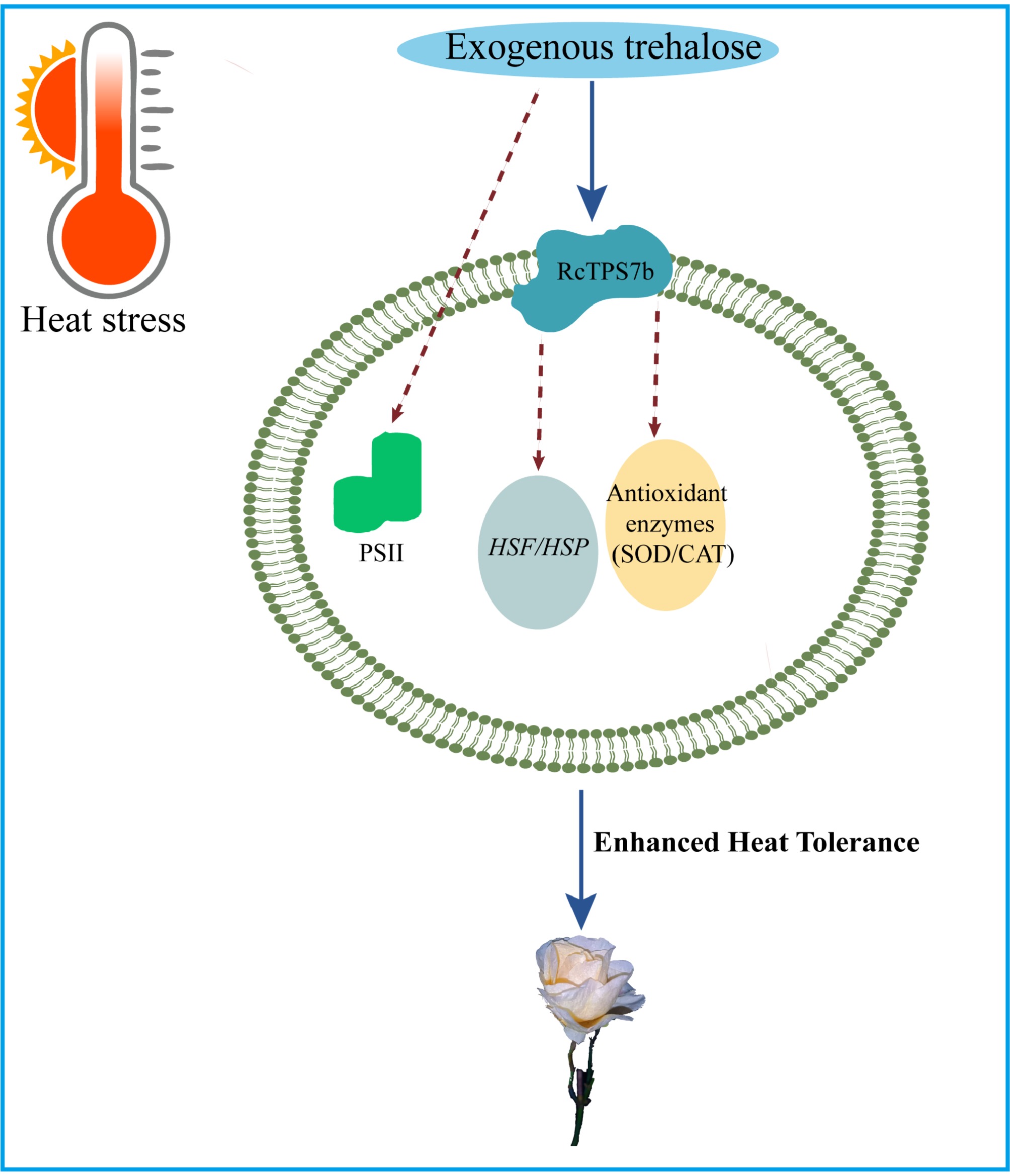
Figure 7.
Trehalose-RcTPS7b-HSF/antioxidant network in heat stress tolerance in R. chinensis.
From an applied perspective, the 50% survival rate of RcTPS7b-overexpressing A. thaliana under heat stress combined with petal protection efficacy (Fig. 6) positions RcTPS7b as a prime candidate for precision breeding. In the future, we will construct transformed plants that stably overexpress RcTPS7b and clustered regularly interspaced short palindromic repeat (CRISPR)-edited RcTPS7b in rose, and analyze the molecular mechanism of heat-resistant signal transmission of RcTPS7b in the cell membrane, as well as the molecular mechanism of its interaction with other heat-responsive genes.
-
This study elucidated the synergistic mechanism of exogenous trehalose and endogenous RcTPS7b in regulating heat tolerance in R. chinensis. We demonstrated that exogenous trehalose enhanced transcriptional levels of RcTPS7b to amplify heat stress responses, while RcTPS7b coordinated dual pathways (activating heat shock signaling and maintaining redox homeostasis) to improve thermal resilience. The work elucidates the dynamic interactions between metabolites and genetic networks, advancing our understanding of plants' environmental adaptation. These findings provide a theoretical basis for the molecular mechanism of heat tolerance in rose.
This work was supported by the Subproject of National Key research and development program, China (Grant No. 2023YFD120010509) and the Natural Science Foundation of Hunan Province of China (Grant No. 2023JJ30292).
-
The authors confirm contributions to the paper as follows: study conception and design: Ma YW, Lin XL, Chen JR, Zhu F, Li YF; data collection: Ma YW, Lin XL, Ding QH, Du JL, Dong XK, Liu A, Liu XD, Zhang YJ; analysis and interpretation of results: Ma YW, Lin XL, Chen HX, Chen JR, Li YF; draft manuscript preparation: Ma YW, Lin XL, Li YF. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yu-Wan Ma, Xin-Lan Lin
- Supplementary Table S1 Primers used in this paper.
- Supplementary Table S2 The TPS7 protein sequences used to reconstruct phylogenetic trees.
- Supplementary Fig. S1 Expression level of endogenous AtTPS7 gene in A. thaliana. (ANOVA, p < 0.05;). Data are presented as mean ± SD of three biological replicates.
- Supplementary Fig. S2 Chlorophyll a and b contents in 30-d-old WT and OE lines after 36 h of heat stress at 45 °C (ANOVA, a > b > c, p < 0.05;). Data are presented as mean ± SD of three biological replicates.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ma YW, Lin XL, Ding QH, Du JL, Dong XK, et al. 2025. Functional identification of trehalose and a trehalose-6-phosphate synthase gene involved in heat stress tolerance of rose. Ornamental Plant Research 5: e039 doi: 10.48130/opr-0025-0038 

Functional identification of trehalose and a trehalose-6-phosphate synthase gene involved in heat stress tolerance of rose
- Received: 11 March 2025
- Revised: 08 August 2025
- Accepted: 21 August 2025
- Published online: 20 October 2025
Abstract: Global warming-induced heat stress increasingly threatens the ornamental quality and productivity of roses. Previous studies have indicated that trehalose and the trehalose-6-phosphate synthase (TPS) gene family regulate plants' stress resistance, yet their roles in thermotolerance in rose remain uncharacterized. We previously identified RcTPS7b as a putative heat-responsive gene in Rosa chinensis 'Slater's Crimson China'. This study aimed to investigate whether exogenous trehalose enhances heat tolerance in rose, and elucidate the functional role of RcTPS7b in heat stress regulation. The results showed that exogenous application of 5 mmol/L trehalose significantly enhanced rose's heat tolerance by reducing the heat damage index, elevating superoxide dismutase (SOD) activity, and preserving the efficiency of Photosystem II (PSII). Overexpression of RcTPS7b in Arabidopsis thaliana enhanced the heat tolerance and antioxidant enzyme activities in transgenic plants, concomitant with upregulated expression of heat-resistant genes such as HEAT SHOCK FACTOR/PROTEIN (AtHSF/P). Furthermore, tobacco rattle virus-induced silencing of RcTPS7b in R. chinensis compromised thermotolerance and induced severe oxidative damage. Transient overexpression of RcTPS7b in rose petals reduced heat damage and maintained petal integrity through redox homeostasis under high temperature treatment. Collectively, this study demonstrates that exogenous trehalose potentiates RcTPS7b expression to enhance thermotolerance, revealing the trehalose metabolism pathway's pivotal role in heat stress regulation in R. chinensis, and will be helpful for the molecular breeding of heat tolerance in R. chinensis.
-
Key words:
- Rosa chinensis /
- Trehalose-6-phosphate synthase gene /
- Heat stress /
- Antioxidant defense