









-
Salinity is one of the most detrimental environmental factors affecting the growth and productivity of cultivated crops. High salt concentrations in the soil and irrigation water lead to a progressive build-up of sodium (Na+) and chlorine (Cl−) in the plant cells. This causes dehydration, nutrient imbalances, and oxidative stresses[1,2] which ultimately restricts the growth and yield of cultivated crops[3−5]. Tomato (Solanum lycopersicum L.) is among the most widely produced and consumed vegetables. Although it is considered a moderately salt-sensitive crop[6,7], a 10% decrease in yield has been reported for each unit of soil electrical conductivity (EC) above a threshold value of 2.5 dS·m–1[8].
Unfortunately, excess soil salinity is an increasing constraint for crop production worldwide. Excessive use of chemical fertilizers, the high salt content of irrigation water, and climate change are considered the main factors deteriorating the situation. Thus, 25% of irrigated agricultural land in the Mediterranean regions is significantly affected by high salinity[9]. This significantly limits their production capacities and farming effectiveness.
Replacing highly saline soils with substrates, or simply growing plants in artificially prepared nutrient solutions, is considered a reliable alternative. Soilless culture systems have been recently developed and are gradually expanding[10]. However, the spatial confinement of roots into a specific, well-defined root zone is a big constraint for these production systems[11]. In container-grown plants, roots grow very dense to compensate for limited root zone volume[12] and are much more exposed to environmental changes and human-imposed mistakes that can impact their morphology and nutrient uptake capabilities[13]. Furthermore, the poor quality of irrigation water remains a primary concern for soilless cultivation systems[14,15].
To relieve salinity pressure, different agricultural practices are applied in both soil-based and soilless production systems. In addition to the development of salt-tolerant cultivars[16], plant inoculation with arbuscular mycorrhizal fungi[17,18]—alone, or in combination with plant growth-promoting bacteria[19]—and plant growth enhancement through high nitrogen doses[20,21] are among commonly used agricultural practices.
In addition, grafting is considered one of the most effective agronomic interventions to alleviate the negative effects of salinity on cultivated crops[22−24]. Although the published results are often contradictory, grafting tomatoes is often considered a powerful tool to increase the yield[25,26]. However, the result depends on the characteristics of the scion and rootstock, their interaction, and stress intensity[6].
As increased market demands and climate changes expose vegetable producers to new challenges, breeding new high-yielding, resilient, resource-use efficient, and high-quality cultivars is a common attitude of the industry[27]. In addition, new rootstocks may help introduce other sources of genotypic and phenotypic variability into the crop[28]. Testing and evaluating their effectiveness under commercial production systems and finding the most appropriate rootstock-scion combination under diverse production systems and growing conditions is a continuously ongoing task.
Although numerous studies have been conducted, the underlying mechanisms by which grafting improves the salt tolerance of vegetable crops remain largely unknown[29]. Recently, we reported that root-restricted tomato plants benefit from appropriate rootstock-scion combinations. Increased root density, steadier growth under fluctuating salinity levels in the irrigation water due to a more stable uptake of nutrient elements, and higher sodium compartmentalization capabilities of the rootstocks were the most distinguished benefits[30]. The question that follows is whether these advantages will be reflected in a higher yield or improved fruit quality of the grafted tomato plants under root restriction saline conditions. Hence, this research aimed to evaluate the effectiveness of several tomato rootstocks, most recently introduced, on the growth and yield of a specific tomato cultivar under root-restricted, saline conditions.
-
The experiment was conducted in a plastic greenhouse in Tirana, Albania, in the spring (March 24−June 23) of 2022. Six different rootstocks (Arnold, Dohkko, Emperador, He-man, Kaiser, and Suzuka; details are presented in Supplementary Table S1) and a tomato cultivar (Izmir) were included in the study. The grafted and self-grafted seedlings were produced by a commercial seedling producer nursery. Grafting was performed using the splice grafting method. Immediately after grafting, all plants were placed in a growth chamber. In the first 3 d, the air temperature was maintained at 26 °C, whereas the relative humidity was 100% at the start and then gradually decreased to 90%. A photosynthetic photon flux density (PPFD) of 100 μmol·m−2·s−1(white, fluorescent lamps) was maintained, with a photoperiod of 12 h. On the fourth day after grafting, plants were moved to a heated greenhouse (air temperatures 18−24 °C, relative humidity 80%, natural lighting) until the experiment started. 65 d after sowing, the grafted and self-grafted seedlings were arranged in a factorial experiment conducted in a non-heated greenhouse. For that purpose, the plants were transplanted in 5 L plastic pots, one plant per pot, filled up with a mixture of peat (Klasmann-Deilmann GmbH, Geeste, Germany) and vermiculite (Dupré Minerals Ltd, Staffordshire, England) (2:1, v/v). The total number of plants of each graft combination was split in half and treated, respectively with the standard nutrient solution (EC 2.2 mS·cm−1), or a 50 mM (EC 8 mS·cm−1) saline solution obtained by adding sodium chloride (NaCl, 3 g·L−1) to the nutrient solution. A mixture of commercial chemical fertilizers containing N, P2O5, K2O, S, and micronutrients was diluted in tap water (pH 7.6, EC 1.2 mS·cm−1) to prepare the nutrient solution (N: 100 mg·L−1, P2O5 25 mg·L−1, K2O 140 mg·L−1). Since the water already contained considerable amounts of Ca (80 mg·L−1) and Mg (35 mg·L−1), no extra amounts of these elements were added to the nutrient solution. The pH of nutrient solutions was 7.6 and 8, respectively; no acidification was performed. A total of eight plants were used in each experimental plot-treatment. The plants were irrigated daily with equal amounts, equivalent to ETp values (3 mm·day−1 at the start of the experiment, 5 mm·day−1 during the harvesting time) corrected with the respective crop coefficient factors, of the normal or saline nutrient solution. To avoid salinity build-up in the root zone, a 20%−25% drainage rate coefficient was applied to all variants. At the beginning of the experiment, pH and EC values measured in the drainage water were similar to the respective values of the fresh nutrient solution. Later, pH of the drainage water remained almost constant, but slightly increased EC values (2.4 mS·cm−1 for the non-saline, and 8.3 mS·cm−1 for saline variants) were measured.
All plants grew up with a single (primary) stem, and plant height and leaf number were measured and recorded sequentially during the experiment. Further, the plant relative growth rate concept[31] was adapted to calculate the relative stem elongation rate (RSER, cm·cm−1·d−1) and relative new leaf emerging rate (RLER, leaf·leaf−1·d−1) in the period from day 1 after transplanting, until day 42 after transplanting.
Three randomly selected plants were harvested 42 d after transplanting for biomass assessment and nutrient analyses. The roots were dissected from the shoots and washed off adhering soil particles, dried (65 °C, 72 h), and the dry matter of roots and shoots of each plant was determined separately to an accuracy of ± 1 mg (TP 303; Denver Instruments GmbH, Göttingen, Germany). Dried shoots (stems + leaves) of the self-grafted plants and two hetero-graft combinations (Dohkko/Izmir, and Suzuka/Izmir) were separately ground with a planetary mill (Pulverisette5; Fritsch, Idar-Oberstein, Germany) for mineral analysis, and nutrient contents (Ca, K, Mg, Na, and P) were analyzed by Inductively Coupled Plasma Optical Emission Spectroscopy (ICP-OES) (Optima 8300; Perkin Elmer, Waltham, USA). For that purpose, the samples were first dissolved in nitrohydrochloric acid to ensure that all minerals were transferred in a liquid solution. The liquid sample was transferred into an argon plasma at a high temperature (6,000−10,000 K), which excites the atoms and ions in the sample, causing them to emit light at characteristic wavelengths. The emitted light is passed through a spectrometer, which separates it into component wavelengths.
The remaining (5) plants were topped up after the fifth cluster was established. Fruits were harvested at full maturity and weighed separately for each experimental unit. The number of fruits was counted for each harvesting date, and the average fruit weight was calculated by dividing the weight of harvested fruits by the respective fruit number. Five randomly selected fruits at the third harvesting date were used to measure soluble solids content (SSC, Brix) and fruit acidity using a digital refractometer (Atago Co. Ltd., Tokyo, Japan).
Statistical analyses
-
A factorial arrangement of 14 treatments (seven rootstock-scion combinations × two levels of salinity), eight replicates each, was employed in a factorial randomized complete block design.
The contribution of different growth and yield parameters under non-saline and saline conditions to the diversity of different rootstock-scion combinations was assessed by the principal component analysis (PCA). To produce a graphical evaluation of their relationships, the respective heat map was generated via the ClustVis (
https://biit.cs.ut.ee/clustvis/ , accessed on 20 October 2024) online program package.Furthermore, to determine the influence of growth rate on plant yield, the respective correlograms of RSER and LRER with yield were accomplished via STHDA (
www.sthda.com ), separately for saline and non-saline conditions.Residuals of all variables were tested for equality of variances and normality using Brown-Forsythe and Shapiro-Wilk tests, respectively. Differences in DM, RSER, RLER, shoot nutrient concentration, yield, and yield quality parameters were tested by two-way ANOVA using the PC program SigmaPlot 13 (Systat Software Inc., San Jose, CA, USA). Each significant ANOVA result (p < 0.05) was followed by a Holm-Sidak test at p < 0.05 as a post hoc test. Values given throughout the text are means ± standard errors (SE).
-
Only slight differences were found among the self-grafted and hetero-grafted variants regarding plant height under normal, non-saline conditions. Although the self-grafted variant (Izmir/Izmir) was among the tallest, the difference with most of the hetero-grafted variants was not significant (Table 1). Significantly fewer leaves (LN) were found in Arnold/Izmir vs the self-grafted variant, which led to a significantly higher internode length (IL). Some variations regarding LN and IL did exist among other grafted variants, but were less important. No differences were found among the self-grafted and hetero-grafted variants under non-saline conditions regarding relative stem elongation rate (RSER), although some variation was found regarding relative new leaves emerging rate (RLER).
Table 1. Plant growth parameters*.
Factors PH (cm) LN IL (cm) RSER (cm·cm−1·d−1) RLER (leaf·leaf−1·d−1) Salinity (S) 0 mM 103.1 ± 1.31A 25.6 ± 0.58A 4.08 ± 0.11A 0.039 ± 0.00A 0.029 ± 0.00A 50 mM 89.8 ± 0.91B 23.7 ± 0.52B 3.84 ± 0.09B 0.036 ± 0.00B 0.027 ± 0.00B Graft combination (Rst/Sc) Arnold/Izmir 95.6 ± 2.78AB 20.8 ± 0.68C 4.63 ± 0.19A 0.040 ± 0.00A 0.023 ± 0.00B Dohkko/Izmir 92.1 ± 2.94B 22.6 ± 0.58BC 4.09 ± 0.14AB 0.037 ± 0.00AB 0.026 ± 0.00B Emperador/Izmir 95.2 ± 2.23AB 23.0 ± 1.04BC 4.20 ± 0.18AB 0.038 ± 0.00AB 0.026 ± 0.00B He-man/Izmir 92.5 ± 2.55B 25.1 ± 0.78B 3.72 ± 0.18B 0.038 ± 0.00AB 0.029 ± 0.00AB Kaiser/Izmir 102.4 ± 2.67A 27.6 ± 0.77A 3.71 ± 0.06B 0.039 ± 0.00A 0.031 ± 0.00A Suzuka/Izmir 98.9 ± 3.05A 27.8 ± 0.57A 3.56 ± 0.12B 0.039 ± 0.00A 0.031 ± 0.00A Izmir/Izmir 98.6 ± 3.74A 26.2 ± 0.81AB 3.79 ± 0.18B 0.035 ± 0.00B 0.029 ± 0.00AB Graft combination
performance under
different salinity
conditions (Rst/Sc × S)Non-saline Arnold/Izmir 103.0 ± 2.66ab 21.2 ± 1.24b 4.92 ± 0.32a 0.041 ± 0.000a 0.024 ± 0.001b Dohkko/Izmir 99.0 ± 3.01b 23.2 ± 0.97ab 4.28 ± 0.15a 0.039 ± 0.001a 0.027 ± 0.000ab Emperador/Izmir 100.0 ± 2.70ab 25.0 ± 1.22a 4.04 ± 0.25a 0.038 ± 0.001a 0.028 ± 0.000ab He-man/Izmir 95.6 ± 4.41b 26.0 ± 0.89a 3.71 ± 0.28b 0.039 ± 0.001a 0.029 ± 0.000ab Kaiser/Izmir 108.2 ± 3.39a 29.4 ± 0.74a 3.68 ± 0.08b 0.040 ± 0.000a 0.033 ± 0.001a Suzuka/Izmir 107.8 ± 1.39a 28.4 ± 0.81a 3.81 ± 0.15b 0.040 ± 0.000a 0.032 ± 0.001a Izmir/Izmir 108.2 ± 2.37a 26.6 ± 1.36a 4.10 ± 0.21a 0.039 ± 0.000a 0.029 ± 0.000ab Saline Arnold/Izmir 88.2 ± 0.66b 20.4 ± 0.6b 4.34 ± 0.14a 0.039 ± 0.000a 0.023 ± 0.001b Dohkko/Izmir 85.2 ± 2.47b 22.0 ± 0.6b 3.89 ± 0.21ab 0.035 ± 0.001ab 0.025 ± 0.001b Emperador/Izmir 90.4 ± 1.91b 21.0 ± 1.1b 4.36 ± 0.27a 0.037 ± 0.001a 0.024 ± 0.000b He-man/Izmir 89.4 ± 2.24b 24.2 ± 1.2ab 3.74 ± 0.25ab 0.037 ± 0.000a 0.029 ± 0.001ab Kaiser/Izmir 96.6 ± 1.96b 25.8 ± 0.7a 3.75 ± 0.08ab 0.038 ± 0.000a 0.029 ± 0.001ab Suzuka/Izmir 90.0 ± 0.70b 27.2 ± 0.8a 3.32 ± 0.12b 0.038 ± 0.000a 0.031 ± 0.001a Izmir/Izmir 89.0 ± 3.36b 25.8 ± 1.0a 3.48 ± 0.23ab 0.032 ± 0.000b 0.029 ± 0.001ab Significance Rst/Sc 0.002 < 0.001 < 0.001 < 0.001 < 0.001 Salinity (S) < 0.001 < 0.001 0.040 < 0.001 0.004 Rst/Sc × S 0.175 0.501 0.187 0.129 0.271 * Salinity (0, 50 mM), Graft combinations (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir, Izmir/Izmir). PH Plant height, LN leaf number, IL internode length, RSER relative stem elongation rate, RLER relative new leaf emerging rate. Different letters (capital letters for main factors, small letters for Rst/Sc × S interactions) indicate significant differences within parameters (Holm-Sidak test, p < 0.05; mean ± SE); significant p-values of a two-way ANOVA are indicated in bold. Raised salinity has significantly reduced growth parameters; plant height (p < 0.001), leaf number (p < 0.001), internode length (p 0.04), relative stem elongation rate (p < 0.001), and relative new leaves emerging rate (p 0.004) to all variants (Table 1). Under such conditions, almost all hetero-graft combinations recorded higher RSER than the self-grafted variant. Kaiser/Izmir and Suzuka/Izmir recorded significantly higher RLER values than the self-grafted plants. Interestingly, hetero-grafted variants were able to sustain RSER despite the rise of salinity, whereas the self-grafted variant did not. In addition, despite the overall reduction in the number of leaves per plant, Suzuka/Izmir recorded the highest value of RLER (Table 1).
Under non-saline conditions, the self-grafted seedlings (Izmir/Izmir) showed a significantly higher root dry matter (DMroot) than the hetero-grafted variants. Suzuka/Izmir was the only exception. However, except for Emperador/Izmir, no differences were found between the self-grafted and hetero-grafted variants regarding the dry matter of shoots (DMshoot) (Table 2). Consequently, the self-grafted plants had a higher ratio of root to shoot dry matter (DMroot : DMshoot) than most of the hetero-grafted variants (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, and He-man/Izmir). The rest—Kaiser/Izmir and Suzuka/Izmir—were included in the same statistical group as Izmir/Izmir (Table 2). The rise of salinity in the irrigation water reduced DMroot and DMshoot to all variants, but the self-grafted variant, together with Suzuka/Izmir and Kaiser/Izmir, recorded the highest DMroot. Almost no differences were found between the self-grafted and hetero-grafted plants regarding DMshoot (Table 2).
Table 2. Plant dry matter parameters*.
Factors Dry matter (g) DMroot DMshoot DMroot : DMshoot Salinity (S) 0 mM 5.80 ± 0.28A 30.21 ± 0.59A 0.19 ± 0.00 50 mM 4.36 ± 0.17B 24.23 ± 0.44B 0.18 ± 0.00 Graft combinations (Rst/Sc) Arnold/Izmir 4.35 ± 0.26B 25.02 ± 1.49B 0.17 ± 0.00B Dohkko/Izmir 4.35 ± 0.14B 26.62 ± 1.58B 0.16 ± 0.00B Emperador/Izmir 4.17 ± 0.34B 30.41 ± 1.82A 0.14 ± 0.01B He-man/Izmir 4.85 ± 0.35AB 26.22 ± 1.43B 0.18 ± 0.00AB Kaiser/Izmir 5.42 ± 0.45AB 26.72 ± 1.12B 0.20 ± 0.00AB Suzuka/Izmir 6.62 ± 0.64A 29.67 ± 1.32A 0.22 ± 0.01A Izmir/Izmir 5.82 ± 0.54AB 26.12 ± 1.49B 0.22 ± 0.01A Graft combination performance under different salinity conditions (Rst/Sc × S) Non-saline Arnold/Izmir 4.90 ± 0.17bc 28.15 ± 1.06b 0.17 ± 0.01b Dohkko/Izmir 4.40 ± 0.23c 29.65 ± 1.47b 0.15 ± 0.01b Emperador/Izmir 4.90 ± 0.11bc 34.45 ± 0.43a 0.14 ± 0.00b He-man/Izmir 5.50 ± 0.23bc 29.30 ± 0.86b 0.18 ± 0.00b Kaiser/Izmir 6.15 ± 0.60b 29.10 ± 0.57b 0.21 ± 0.01a Suzuka/Izmir 7.80 ± 0.63a 32.50 ± 0.86ab 0.19 ± 0.01a Izmir/Izmir 7.00 ± 0.28ab 28.45 ± 1.70b 0.24 ± 0.00a Saline Arnold/Izmir 3.80 ± 0.11c 21.90 ± 0.57c 0.17 ± 0.00b Dohkko/Izmir 4.30 ± 0.23c 23.60 ± 1.09bc 0.18 ± 0.01b Emperador/Izmir 3.45 ± 0.20c 26.35 ± 0.37b 0.13 ± 0.00c He-man/Izmir 4.20 ± 0.40c 23.15 ± 0.14bc 0.18 ± 0.01b Kaiser/Izmir 4.70 ± 0.34bc 24.35 ± 0.54b 0.23 ± 0.00 a Suzuka/Izmir 5.40 ± 0.54b 26.85 ± 0.20b 0.20 ± 0.02ab Izmir/Izmir 4.65 ± 0.20bc 23.80 ± 1.67bc 0.19 ± 0.00b Significance Rst/Sc < 0.001 < 0.001 < 0.001 Salinity (S) < 0.001 < 0.001 0.067 Rst/Sc × S 0.053 0.643 0.062 * Salinity (0, 50 mM), Graft combinations (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir, Izmir/Izmir). DMroot dry matter weight of roots, DMshoot dry matter of shoots, DMroot : DMshoot the ratio of root dry matter with shoot dry matter. Different letters (capital letters for main factors, small letters for Rst/Sc × S interactions) indicate significant differences within parameters (Holm-Sidak test, p <0.05; mean ± SE); significant p-values of a two-way ANOVA are indicated in bold. Rootstock-scion combinations and salinity effects on plant's nutrient concentration
-
Large variations in nutrient concentrations were found among the grafted plants. Lower Ca concentrations, but significantly higher P concentrations were found in the self-grafted plants compared with hetero-grafted variants under non-saline conditions. No difference was found regarding Na concentration. Although the rise of salinity in the irrigation water multiplied shoot Na concentration several times, there was no difference between the self-grafted and hetero-grafted variants (Table 3). The increase of salinity in the irrigation water did not affect Ca concentration, but significantly reduced the shoot concentration of K and P. On the contrary, Mg concentration was significantly increased alongside an immense increase in Na concentration. Ca, K, Mg, and P concentrations were also significantly affected by Rst/Sc combinations. Higher Mg and P concentrations were found in the shoots of the self-grafted variant compared to both hetero-grafted ones. However, the hetero-grafted variants were able to maintain higher K (Suzuka/Izmir, Dohkko/Izmir) and Ca (Dohkko/Izmir) shoot concentrations under saline conditions.
Table 3. Plant macronutrient (Ca, K, Mg, Na, P, and S) concentration*.
Factors Rs/Sc combination Ca K Mg Na P Salinity (S) 0 mM 29.25 ± 0.77 23.57 ± 0.38A 8.93 ± 0.31B 1,189 ± 33B 4,602 ± 287A 50 mM 29.31 ± 0.77 21.35 ± 0.38B 9.73 ± 0.39A 4,011 ± 47A 4,560 ± 244B Graft combinations (Rst/Sc) Izmir/Izmir 29.25 ± 0.35B 22.07 ± 1.11B 10.12 ± 0.32A 2,598 ± 762 5,201 ± 139A Dohkko/Izmir 27.24 ± 0.24C 22.17 ± 0.16B 8.28 ± 0.17B 2,560 ± 835 4,708 ± 126B Suzuka/Izmir 31.33 ± 0.24A 23.14 ± 0.69A 9.58 ± 0.21A 2,643 ± 848 3,833 ± 22C Graft combination performance under different salinity conditions (Rst/Sc × S) Non-saline Izmir/Izmir 28.721 ± 0.07b 23.953 ± 0.48a 9.572 ± 0.02b 1,279 ± 37b 5,429 ± 8.35a Dohkko/Izmir 27.505 ± 0.48c 22.438 ± 0.09b 7.982 ± 0.03d 1,114 ± 29b 4,507 ± 91.0c Suzuka/Izmir 31.544 ± 0.52a 24.329 ± 0.01a 9.238 ± 0.08bc 1,174 ± 11b 3,869 ± 20.5d Saline Izmir/Izmir 29.791 ± 0.39b 20.186 ± 0.26c 10.678 ± 0.19a 3,917 ± 60.9a 4,974 ± 118.1b Dohkko/Izmir 26.992 ± 0.03c 21.910 ± 0.12b 8.590 ± 0.01c 4,006 ± 97.1a 4,909 ± 84.4b Suzuka/Izmir 31.122 ± 0.01a 21.959 ± 0.28b 9.936 ± 0.03b 4,112 ± 42.2a 3,798 ± 3.26d Significance Rst/Sc < 0.001 0.011 < 0.001 0.366 < 0.001 Salinity (S) 0.875 < 0.001 < 0.001 < 0.001 0.05 Rst/Sc × S 0.098 0.002 0.065 0.063 0.003 * Salinity (0, 50 mM), Graft combinations (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir, Izmir/Izmir). Different letters (capital letters for main factors, small letters for Rst/Sc × S interactions) indicate significant differences within parameters (Holm-Sidak test, p <0.05; mean ± SE); significant p-values of a two-way ANOVA are indicated in bold. Rootstock-scion combinations and salinity effects on yield
-
Hetero-grafting did not provide any increase in the yield versus the self-grafted variants. No difference in the number of fruits per plant between the self-grafted and hetero-grafted variants was observed under either non-saline or saline conditions. The average fruit size of the self-grafted variant was generally higher under non-saline conditions, but no differences existed under saline conditions (Table 4). There were cases of hetero-grafted variants (Armold/Izmir under non-saline conditions) with a lower yield than the self-grafted variant. No correlations were found between RSER and RLER with yield in non-saline conditions, but some mild positive correlations were found between RLER and yield under saline conditions (Fig. 1a, b).
Table 4. Yield (harvested yield, kg·var−1; fruit number; average fruit weight, g), and fruit quality (Brix; acidity) parameters*.
Factors Total yield (kg·plant−1) Fruit number Average fruit weight (g) Brix Acidity Salinity (S) 0 mM 3.75 ± 0.06A 35.02 ± 0.66 108.06 ± 2.35 5.38 ± 0.07B 0.91 ± 0.02B 50 mM 1.51 ± 0.03B 32.31 ± 0.59 47.07 ± 1.08 9.28 ± 0.11A 1.67 ± 0.04A Graft combinations (Rst/Sc) Arnold/Izmir 2.37 ± 0.37 32.0 ± 1.06 71.86 ± 9.46B 7.33 ± 0.74 1.34 ± 0.16A Dohkko/Izmir 2.50 ± 0.40 34.2 ± 1.41 70.82 ± 9.66B 7.53 ± 0.84 1.43 ± 0.17A Emperador/Izmir 2.64 ± 0.42 34.1 ± 1.33 77.26 ± 11.96AB 7.01 ± 0.72 1.30 ± 0.14AB He-man/Izmir 2.70 ± 0.36 34.6 ± 1.20 82.31 ± 10.36AB 7.12 ± 0.66 1.35 ± 0.18A Kaiser/Izmir 2.66 ± 0.34 34.1 ± 1.03 78.76 ± 10.56AB 7.60 ± 0.83 1.29 ± 0.17AB Suzuka/Izmir 2.66 ± 0.36 35.2 ± 1.63 75.05 ± 9.11AB 7.72 ± 0.74 1.27 ± 0.13AB Izmir/Izmir 2.88 ± 0.41 33.5 ± 1.05 86.91 ± 12.72A 7.03 ± 0.69 1.04 ± 0.09B Graft combination performance under different salinity conditions (Rst/Sc × S) Non-saline Arnold/Izmir 3.46 ± 0.18b 34.60 ± 0.81a 99.89 ± 3.16b 5.375 ± 0.19b 0.915 ± 0.04c Dohkko/Izmir 3.71 ± 0.09ab 37.40 ± 1.03a 99.37 ± 2.85b 5.375 ± 0.14b 1.008 ± 0.12c Emperador/Izmir 3.91 ± 0.12ab 35.00 ± 2.21a 112.7 ± 3.52ab 5.125 ± 0.22b 0.938 ± 0.07c He-man/Izmir 3.68 ± 0.31ab 34.00 ± 2.16a 109.3 ± 10.4ab 5.375 ± 0.08b 0.920 ± 0.09c Kaiser/Izmir 3.67 ± 0.12ab 33.80 ± 1.28a 109.4 ± 5.34ab 5.450 ± 0.27b 0.843 ± 0.03c Suzuka/Izmir 3.72 ± 0.19ab 37.20 ± 2.65a 101.12 ± 4.97b 5.800 ± 0.22b 0.94 ± 0.05bc Izmir/Izmir 4.11 ± 0.08a 33.20 ± 1.59a 124.59 ± 3.85a 5.225 ± 0.16b 0.818 ± 0.09c Saline Arnold/Izmir 1.28 ± 0.04c 29.40 ± 1.03b 43.82 ± 0.63c 9.300 ± 0.08a 1.773 ± 0.06a Dohkko/Izmir 1.29 ± 0.02c 31.00 ± 1.67b 42.27 ± 2.13c 9.700 ± 0.39a 1.867 ± 0.10a Emperador/Izmir 1.38 ± 0.06c 33.20 ± 1.65a 41.77 ± 1.58c 8.900 ± 0.23a 1.670 ± 0.06a He-man/Izmir 1.71 ± 0.07c 31.20 ± 0.91b 55.31 ± 3.12c 8.875 ± 0.11a 1.788 ± 0.15a Kaiser/Izmir 1.64 ± 0.03c 34.40 ± 1.77a 48.12 ± 2.03c 9.750 ± 0.31a 1.738 ± 0.07a Suzuka/Izmir 1.60 ± 0.03c 33.20 ± 1.74a 48.98 ± 3.04c 9.650 ± 0.31a 1.615 ± 0.08a Izmir/Izmir 1.65 ± 0.05c 33.80 ± 1.56a 49.23 ± 1.98c 8.850 ± 0.20a 1.277 ± 0.04b Significance Rst/Sc 0.983 0.544 0.003 0.989 0.003 Salinity (S) < 0.001 0.003 < 0.001 < 0.001 < 0.001 Rst/Sc × S 0.204 0.257 0.050 0.474 0.158 *Salinity (0, 50 mM), Graft combinations (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir, Izmir/Izmir). Different letters (capital letters for main factors, small letters for Rst/Sc × S interactions) indicate significant differences within parameters (Holm-Sidak test, p < 0.05; mean ± SE); significant p-values of a two-way ANOVA are indicated in bold. 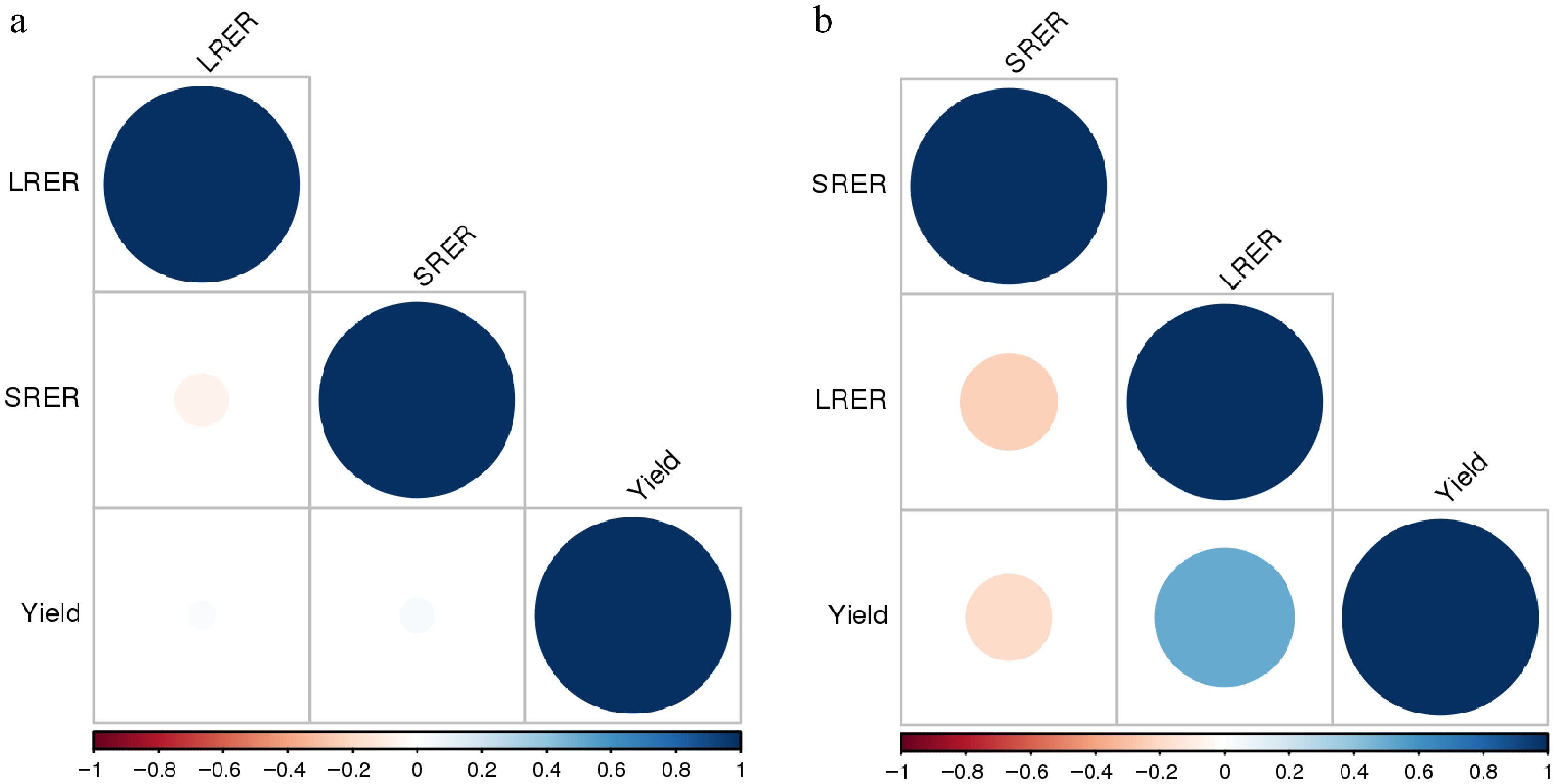
Figure 1.
(a) Yield-RSER-RLER correlograms of hetero-grafted and self-grafted tomato plants (cv. Izmir F1), under non-saline conditions. RSER (relative stem elongation rate), RLER (relative new leaf emerging rate). (b) Yield-RSER-RLER correlograms of hetero-grafted and self-grafted tomato plants (cv. Izmir F1), under saline conditions. RSER (relative stem elongation rate), RLER (relative new leaf emerging rate).
To all variants, the yield was drastically reduced due to the rise of the salinity level in the irrigation water (Table 4). Mostly, the yield reduction was a consequence of significantly reduced (more than halved) average fruit weight. However, a significant reduction in the number of fruits per plant due to raised salinity was observed in several hetero-grafted variants (Arnold/Izmir, Dohkko/Izmir, He-man/Izmir) (Table 4). No effects of grafting were found regarding soluble solids content (Brix) and acidity under non-saline conditions, whereas some variations were found regarding acidity under saline conditions. Under such conditions, the self-grafted variant (Izmir/Izmir) appeared with the lowest acidity level (1.277), significantly lower than the rest of the grafted variants (Table 4).
The principal component analyses (PCA) have shown that the most contributing factors to the variability were the plant height (PH, −8.97) and average fruit weight (−6.15). The rest of the factors were grouped but still split into two subgroups with either positive (RSER (2.52), RLER (2.53), DMroot : DMshoot (2.51), fruit acidity (2.36), yield (2.24), DMroot (1.93), Brix (1.6)), or negative (fruit number (−1.51), DMshoot (−0.7), LN (−0.43)) values (Table 5). Most of the variability was represented by the first principal component (PC1, 96%), and the next principal component (PC2) accounted for nearly 4%. The non-saline and saline variants were clearly distinguished (Fig. 2), whereas the plant traits most affected by raised salinity were stem length and average fresh fruit weight.
Table 5. The principal components scores of growth, yield, and fruit quality traits*.
Traits PC1 PC2 PC3 PC4 PC5 PC6 PC7 PC8 PC9 DMroot 1.93 0.06 0.02 0.05 −0.02 0.04 0.03 0 0 DMshoot −0.7 −0.17 0.04 −0.19 −0.08 0 0 0 0 DMroot : DMshoot 2.51 0.08 −0.03 0.02 0 0.01 −0.01 0 0 PH −8.97 −1.26 −0.08 0.07 −0.02 0.01 0 0 0 LN −0.43 −0.33 0.23 0.08 0 −0.01 −0.01 0 0 IL 2.05 0.01 −0.08 0 0 −0.01 0 0.01 0 RSER 2.52 0.08 −0.03 0.02 0 0.01 −0.01 0 0 RLER 2.53 0.08 −0.03 0.02 0 0.01 −0.01 0 0 Total yield 2.24 0.16 −0.03 0.01 0.01 0.02 −0.01 0 0 Fruit number −1.51 −0.5 0.03 −0.12 0.1 0.01 0.01 0 0 Avg. fruit weight −6.15 2.11 0 0.01 0.01 −0.01 0 0 0 Brix 1.6 −0.31 −0.01 0.03 0 −0.06 0.02 0 0 Acidity 2.36 0.01 −0.04 0.02 0 −0.01 0 −0.01 −0.01 * The self-grafted (Izmir/Izmir) and heterografted (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir) tomato variants under non-saline (0 mM NaCl) and saline (50 mM NaCl) conditions. 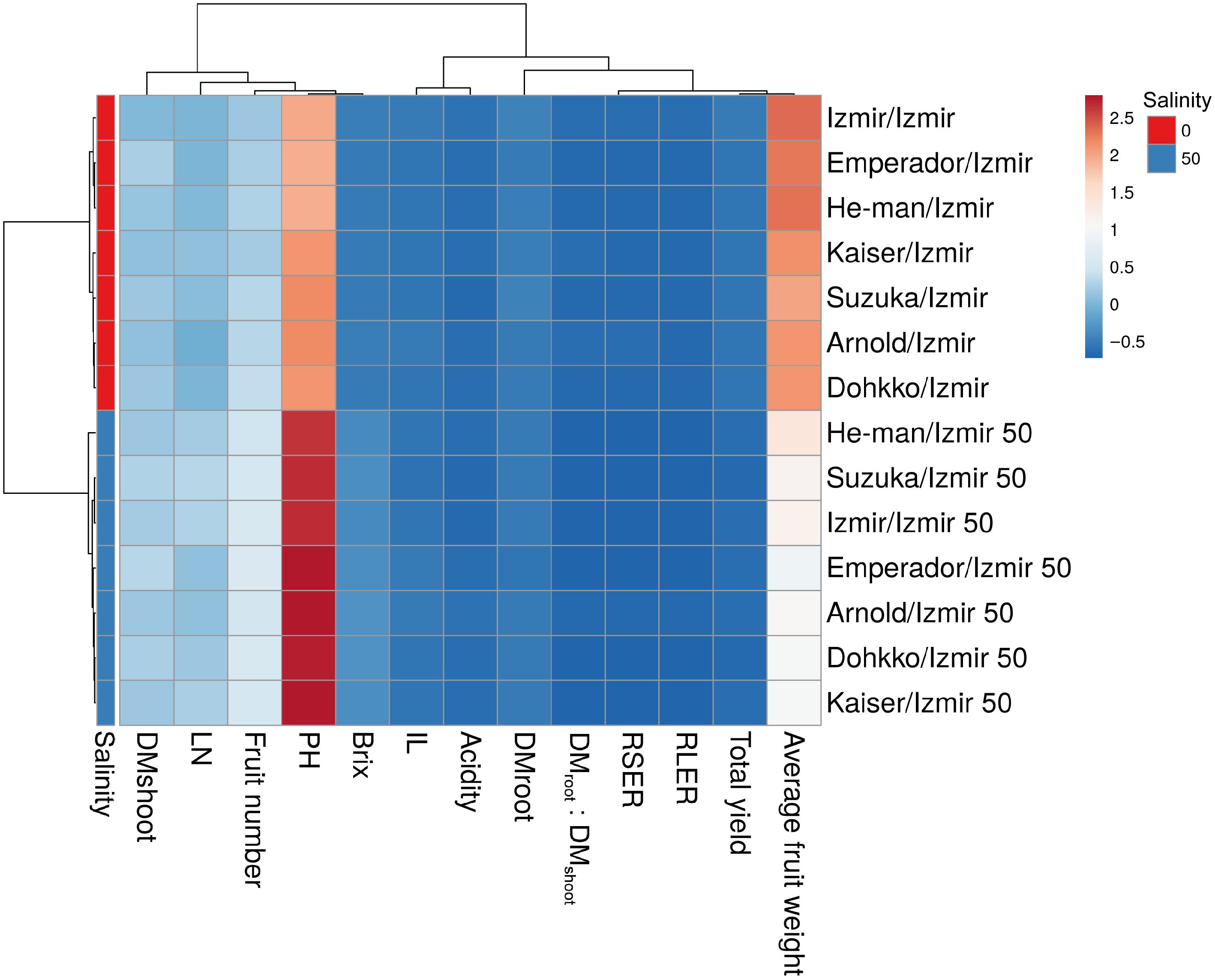
Figure 2.
The heatmap of growth, yield, and fruit quality traits of self-grafted (Izmir/Izmir) and heterografted tomato variants (Arnold/Izmir, Dohkko/Izmir, Emperador/Izmir, He-man/Izmir, Kaiser/Izmir, Suzuka/Izmir) under non-saline (0 mM NaCl) and saline (50 mM NaCl) conditions. Rows are centered; unit variance scaling is applied to rows. PH Plant height, LN leaf number, IL internode length, RSER relative stem elongation rate, RLER relative new leaf emerging rate, DMroot dry matter weight of roots, DMshoot dry matter of shoots, DMroot : DMshoot the ratio of root dry matter with shoot dry matter. Both rows and columns are clustered using correlation distance and average linkage. Fourteen rows, 13 columns.
Interesting conclusions can be drawn from the heatmap chart regarding the variability of different rootstock-scion combinations under both non-saline and saline conditions. The non-grafted (Izmir/Izmir) variant was the most distinguished under non-saline conditions. Closely followed were Emperador/Izmir and He-man/Izmir. The remaining variants (Arnold/Izmir, Dohkko/Izmir, Kaiser/Izmir, Suzuka/Izmir) were grouped. On the contrary, He-man/Izmir was the most distinguished variant under saline conditions. Closely followed were Suzuka/Izmir and Izmir/Izmir. All remaining variants were grouped (Fig. 2).
-
It is commonly reported that grafting enhances growth and produces more vigorous plants[23,26]. Although often seen as an advantage of grafted plants, this might be a concern for greenhouse tomatoes. The vegetative plants increase the chances of foliar fungi diseases[32], induce assimilate partitioning imbalances between fruits and vegetative organs in the plants[33], and delay fruit setting and maturity[34]. Therefore, the rootstocks that produce less vigorous, compact plants fit better into a greenhouse environment[27] and might potentially allocate more carbohydrates toward fruit formation[27,35]. No significant differences between the self-grafted and hetero-grafted plants was found regarding stem height. Some of the rootstocks were able to shorten the length of plant internodes which satisfies the demand for compact plants in greenhouse production. No difference exists between the self-grafted and hetero-grafted plants regarding dry matter of shoots (DMshoot). Conversely, the self-grafted plants had a higher dry matter of roots (DMroot) than most of the grafted variants which grants them the highest DMroot : DMshoot ratio. Previous reports confirm that the reduced growth of the root system due to grafting is compensated by an enhancement in root enzyme activity[36].
Although plant response to salinity is highly variable[37], commonly, salinity reduces plant growth[19] by inhibiting both cell division and elongation[38]. Yet, little is known about salt-induced changes in shoot growth rate[4]. Interestingly, the study found significant differences between self-grafted and hetero-grafted plants regarding relative stem elongation rate (RSER) and relative new leaf emerging rate (RLER). The enhanced shoot growth and the increased ability of certain rootstock-scion combinations to produce new leaves indicate that they are more capable of alleviating the salinity stress by improving the osmotic tolerance of grafted plants[37]. Maintaining an appropriate stem growth rate was considered essential to achieving a good yield in cucumber plants[39]. In contrast, the study did not find any significant correlation between relative stem elongation rate, or relative new leaf emerging rate with tomato yield in non-saline conditions, but good correlations were found between the relative new leaf emerging rate and yield under saline conditions.
Grafting is often considered a tool to increase yield in several vegetable crops[17,22,40]. On the contrary, the rootstocks involved in this experiment were not able to increase tomato yield in the condition of reduced root volume, either in non-saline or saline conditions. No advantage of grafted variants was found versus the non-grafted plants, either regarding the number of harvested fruits per plant, or the average fruit weight. In contrast, in a previous publication of Djidonou et al.[40] an increase in the yield of grafted tomatoes was supported by a higher number of larger fruits. Still, similar to us, di Gioia et al.[25] did not report any significant change in yield components of a heirloom tomato variety grafted onto two interspecific (S. lycopersicum × S. habrochaites) rootstocks. More recently, Parisi et al.[41] reported an increase in harvested yield by only three out of seven rootstocks used in grafting with an Italian tomato landrace, and Gong et al.[42] found varying results depending on specific rootstock-scion combinations and planting season. Considering a large number of different, even contradictory, evidence, the study concludes that the wide range of rootstock-scion combinations leads to mixed results. Huge differences regarding specific production systems and the interactions of specific genotypes with production systems make the performance of grafted tomato plants even more complicated[43].
The higher yield in grafted plants is often linked with a more vigorous rootstock root system, its enhanced root hydraulic conductance, and nutrient uptake[44]. However, if the growing system can maintain a constant concentration of nutrient elements at the root surface, the restricted root system is no longer a limiting factor[11]. Under these circumstances, frequent fertigation fulfills plants' requirements through the continuous replenishment of nutrients in the vicinity of the root interface[45]. Indeed, despite the differences between self-grafted and hetero-grafted plants, shoot concentrations of Ca, K, Mg, and P to all variants varied within the range of respective recommended optimum concentrations[46]. We discussed in detail the differences between the self-grafted and heterografted variants regarding the root and shoot nutrient concentrations in a previous publication[30]. Even under salinity conditions where the shoot concentration of some elements (K, Mg, P) falls, it remained within the recommended range of optimum concentration. Therefore, improved nutrient uptake in several rootstocks was not enough to distinguish the grafted and non-grafted plants regarding the harvested yield. To increase the efficiency of grafted plants under such production systems, further studies regarding the appropriate management of limited root systems to ensure adequate nutrition, optimum irrigation frequency, proper root zone temperature, and oxygenation level are needed[13].
In both the self-grafted and hetero-grafted plants, yield was drastically reduced by increased salinity. A slight reduction in fruit number was noticed, but the real factor that led to severe yield reduction was the drop in average fruit weight by more than half. No difference was found between the self-grafted and hetero-grafted variants regarding soluble solids content (SSC) and fruit acidity under non-saline conditions. In line with the study, Gong et al.[47] concluded that scion cultivars have a much more pronounced impact on fruit quality attributes than rootstocks. On the contrary, SSC and acidity were significantly increased by the rise of salinity in the irrigation water. Under such conditions, no differences were found between the self-grafted and hetero-grafted variants regarding SSC, but a distinctive difference was found regarding fruit acidity. The self-grafted plants showed a significantly lower acidity level than each of the hetero-grafted variants. As such, the results conform with a previous report by Turhan et al.[48] who reported that grafting tomatoes with interspecific rootstocks (S. lycopersicum × S. habrochaites) increased titratable acidity relative to the non-grafted controls. Contrary to the study, Djidonou et al.[40,49] and Parisi et al.[41] concluded that either beefsteak or cherry tomatoes grafted with interspecific rootstocks did not affect fruit SSC, acidity, and SSC/TA ratio. A clear answer regarding this issue is further complicated by the fact that rootstocks with similar genetic backgrounds could exert different impacts on tomato fruit sensory attributes depending on the scion genotype and growing conditions[48].
Overall, the results of this experiment prove that there is not always a clear distinction between the non-grafted and grafted tomato plants under root-restricted conditions. Only carefully selected rootstock-scion combinations might make a distinction. Therefore, the effects of grafting itself and the role of specific rootstock-scion combinations on tomato growth, yield, and the fruit quality remain an open question for further elaboration.
-
Grafted plants demonstrated improved shoot growth and new leaf emerging rate, indicating enhanced capabilities to alleviate the salinity stress. Good correlations were found between the new leaf emerging rate and yield under saline conditions. Considering the low variability found between the self-grafted and grafted tomato variants under root-restricted volume conditions, only carefully selected rootstock-scion combinations might make a distinction. The study's result suggest that He-man can be considered an appropriate rootstock for root-restricted grafted tomatoes under salinity conditions.
-
The authors confirm their contribution to the paper as follows: study conception and design, analysis and interpretation of results, and draft manuscript preparation: Balliu A; data collection: Pellumbi G; analysis, and interpretation of results: Sallaku G. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
We are grateful to AFC/GIZ for the support in conducting the field experiment, and to Marcel Hirsch (BOKU University) for performing ICP-OES analyses.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The tomato rootstocks involved in the experiment and their main characteristics.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Balliu A, Pellumbi G, Sallaku G. 2025. Do rootstock-scion combinations and irrigation water salinity affect the growth and yield of grafted tomatoes under root-restricted conditions? Technology in Horticulture 5: e026 doi: 10.48130/tihort-0025-0022 

Do rootstock-scion combinations and irrigation water salinity affect the growth and yield of grafted tomatoes under root-restricted conditions?
- Received: 25 June 2024
- Revised: 01 April 2025
- Accepted: 30 April 2025
- Published online: 03 July 2025
Abstract: The experiment evaluated the impact of several commercial rootstocks on the growth and yield of a selected tomato cultivar under restricted volume conditions. The experiment included six different rootstocks and a single tomato cultivar (Solanum lycopersicum cv. Izmir). The respective rootstock-scion combination and self-grafted tomato plants were tested in a randomized complete block design. The plants, transplanted into 5 L plastic pots, were irrigated daily with equal amounts of a nutrient solution and were subject to two different levels of salinity, 0- or 50-mM NaCl solution, respectively. The relative stem elongation rate and new leaf emergence rate were calculated from day 1 to day 42 after transplanting, and nutrient concentrations of mature leaves were analyzed by the end of this period. Fruits were harvested at full maturity, and Brix (°Bx) and fruit acidity were measured using a digital refractometer. While the Na concentration of mature leaves significantly increased, the rise of salinity in the nutrient solution significantly reduced the plants' relative growth rate, K and P concentration of mature leaves, and the yield of tomato plants. Although the grafted plants demonstrated improved shoot growth and increased new leaf emergence rate, indicating enhanced capabilities to alleviate salinity stress, most of the rootstocks did not contribute to any increase in tomato yield. Since only carefully selected rootstock-scion combinations might contribute to higher yields and better quality, the introduction of new rootstocks should be followed by continuous testing of their compatibility with specific, well-recognized tomato cultivars under diverse growing conditions.