









-
Chili pepper (Capsicum annuum L.), ranking third globally in vegetable production after tomato and potato, faces severe yield limitations due to plant diseases[1]. Phytophthora capsici, a highly dynamic and destructive oomycete pathogen, is a major cause of pepper phytophthora blight, one of the most devastating soil-borne diseases affecting pepper crops[2,3]. It was first identified as the causative agent of pepper blight in 1922 by Leonian[4]. Originally thought to be host-specific, P. capsici now infects a broad range of hosts, spanning at least 30 plant families, including vegetables and tropical fruits[3,5]. Notably, P. capsici primarily targets Solanaceae crops, especially members of the Capsicum genus, such as C. annuum (bell pepper, red chili, and chili pepper), C. pubescens, C. baccatum, C. chinense, and C. frutescens[5,6]. In the presence of water, sporangia release zoospores that migrate into the plant root system[7]. Flooding, heavy rainfall, and irrigation facilitate the movement of these propagules to aboveground plant parts[8]. Under favorable conditions, P. capsici can infect all parts of the pepper plant and spread rapidly, causing symptoms such as crown and root rot, greyish-brown water-soaked lesions on the leaves, and black lesions on the stems and fruits[2,9]. These infections often lead to plant wilting and death, significantly reducing pepper yield and quality[10]. Although global estimates of annual losses caused by P. capsici have not been precisely quantified, regional reports have highlighted its severe impact. For instance, outbreaks in Ontario, Canada resulted in 40%–60% yield losses in pepper (C. annuum) fields, whereas 80% of chili pepper fields in New Mexico were affected by P. capsici[11]. Furthermore, P. capsici can persist in the soil for many years, causing recurrent disease outbreaks if not effectively managed. Consequently, P. capsici remains a critical limiting factor in pepper production worldwide[5].
Traditionally, the management of plant diseases relies heavily on cultivation techniques, such as water management, crop rotation, and grafting[5]. However, with the growing need for sustainable disease control, the primary approach for managing pepper Phytophthora blight has shifted from conventional methods to synthetic chemical fungicides. These fungicides offer broad-spectrum activity against multiple pathogens, and provide rapid prevention and control[11,12]. Despite its efficiency in controlling Phytophthora blight, the genetic variability of P. capsici enables it to adapt rapidly to adverse conditions, thereby reducing the effectiveness of fungicides over time. Furthermore, the long-term application of fungicides has led to significant environmental concerns, including soil, water, and air pollution; toxicity to non-target organisms; emergence of fungicide-resistant pathogens; reduced biodiversity; and potential risks to food safety due to fungicide accumulation[13,14].
As antagonistic agents against plant pathogens, the application of crop-beneficial microorganisms that naturally inhabit plant tissues and the rhizosphere has been recognized as an environmentally friendly alternative to control plant diseases, while reducing the over-use of chemical fungicides[15,16]. The genus Bacillus is one of the most common antagonistic bacteria used as a plant biocontrol agent. Bacillus antagonists constitute > 50% of the total antagonistic consortium[17]. Bacillus antagonists suppress various diseases induced by soil-borne pathogens in crops and vegetables, such as maize root rot caused by Fusarium moniliforme[18], tomato early blight caused by Alternaria solani[19], and pepper bacterial spots caused by Xanthomonas euvesicatoria[1]. Among the multitudinous Bacillus antagonists, a diverse range of Bacillus amyloliquefaciens-derived biopesticides are the most recognized biocontrol agents[20]. Recent studies have confirmed that B. amyloliquefaciens possesses strong antipathogenic characteristics, especially against P. capsici through mechanisms such as the production of antimicrobial metabolites, including antibiotics and antimicrobial volatile organic compounds, competition for ecological niches and nutrients, and induction of systemic resistance (ISR) in plants[21]. For example, B. amyloliquefaciens strain IBFCBF-1 exhibits strong biocontrol activity against P. capsici, and promotes pepper plant growth by releasing volatile organic compounds that inhibit mycelial growth[22]. Ngo et al.[23] reported that the pepper rhizosphere strains B. amyloliquefaciens, B. velezensis, and B. methylotrophicus have high in vitro antagonistic activities and inhibit > 90% of the lesions caused by P. capsici in pepper shoots because of their high chitinase and protease activities. Synergistic interactions of the combined B. amyloliquefaciens strains Fy11 and Zy44 isolated from Ginkgo biloba leaves greatly suppressed pepper phytophthora blight owing to the direct antagonism of antipathogenic lipopeptides, and indirect inhibition through ISR induction[24]. In summary, B. amyloliquefaciens is a potential biocontrol agent against P. capsici, and the development of B. amyloliquefaciens with biological control functions against P. capsici is of great significance for the green production and sustainable development of pepper.
In the present study, we investigated the antagonistic bacterium JK2 and its potent in vitro inhibitory activity against P. capsici. Morphological characterization and molecular identification of P. capsici were conducted, and transcriptomic analysis was employed to elucidate the inhibitory mechanism of JK2 against P. capsici under in vitro conditions. Furthermore, the biocontrol efficacy of JK2 against phytophthora blight was evaluated through a greenhouse-based pot experiment. This study aimed to characterize the antagonistic strain JK2 and its inhibitory efficacy against P. capsici, elucidate its molecular mechanisms through transcriptomic analysis, evaluate JK2's biocontrol potential in pepper plants under P. capsici infection, and investigate JK2-mediated modulation of oxidative stress in host plants.
-
The bacterial strain JK2 was isolated from honey peach fruit, identified, and preserved in the Laboratory of Agro-Product Storage at the Chengdu Academy of Agriculture and Forestry Sciences (Chengdu, China). The fungal strain P. capsici and red chili seed cultivar 'Mexican pepper No. 1', which is susceptible to phytophthora blight, were provided by the Vegetable Research Laboratory of Sichuan Agricultural University, Chengdu, China.
In vitro antagonistic activity of JK2 against P. capsici
-
The activated Bacillus amyloliquefaciens JK2 strain was cultured in sterilized Luria-Bertani (LB) broth at 37 °C with orbital shaking (150 rpm) for 24 h. The bacterial suspension was subsequently diluted with sterile distilled water to achieve a final concentration of 1 × 108 CFU mL−1. Following the protocol of Sun et al.[25], 15 mL of sterilized V8 agar medium (cooled to 55 °C) was transferred to a sterile centrifuge tube, mixed with 100 μL of the prepared JK2 suspension, and poured into a sterile Petri dish. After solidification at room temperature, a 7 mm mycelial stipe of P. capsici was centrally placed on the medium. Control plates received no JK2 inoculation. All treatments were performed in triplicate and incubated at 28 °C for 7 d under controlled conditions[22]. Colony diameters were measured, and the inhibition ratio was calculated using the formula below[10]:
Inhibition ratio = (Control colony diameter − Treated colony diameter)/(Control colony diameter − 7 mm) × 100%
Morphological identification of the biocontrol agent JK2
-
The JK2 strain was streaked onto Luria-Bertani (LB) agar plates using a three-zone streaking method and incubated at 37 °C for 24 h. Colony morphological features, including size, shape, margin, surface texture, opacity, and pigmentation, were documented photographically following the taxonomic guidelines outlined in Bergey's Manual of Systematic Bacteriology[26]. Gram staining was performed using a commercial Gram stain kit (Hopebio Bio-Technology Co., Ltd, Qingdao, China), and cellular morphology was examined under an oil-immersion microscope (BX53, Olympus, Tokyo, Japan). Gram-positive cells exhibited dark purple to purple coloration, while Gram-negative cells appeared red to light pink. For endospore visualization, air-dried smears were stained with saturated malachite green solution (10 min), rinsed with distilled water, counterstained with safranin (30 s), and examined microscopically after final washing and drying.
Molecular identification of the biocontrol agent JK2
-
Genomic DNA was extracted from strain JK2 using a Rapid Bacterial Genomic DNA Isolation Kit (Tsingke Biotech Co., Ltd., Beijing, China) according to the manufacturer's instructions. The universal 16S rRNA primers 27F (5'-AGAGTTTGATCCTGGCTCAG-3') and 1492R (5'-TACGGTTACCTTGTTACGACTT-3') were amplified in a 50 μL PCR mixture containing: 5 μL of 10 × PCR buffer, 1 μL of DNA template, 4 μL of dNTP (2.5 mmol·L−1), 3 μL of MgCl2 (25 mmol·L−1), 1 μL of each primer, 0.5 μL of Taq polymerase, and 34.5 μL of ddH2O. Thermal cycling parameters included: initial denaturation at 94 °C for 5 min, 30 cycles of denaturation at 94 °C for 30 s, annealing at 53 °C for 30 s, extension at 72 °C for 1 min, and holding at 4 °C[22]. PCR amplicons were sequenced commercially (Beijing Tsingke Biotech Co. Ltd., Beijing, China), and sequences were aligned against the NCBI database using BLAST (
http://blast.ncbi.nlm.nih.gov/Blast.cgi ). Phylogenetic reconstruction was performed in MEGA 7.0 (www.megasoftware.net ) using the neighbor-joining algorithm with 1,000 bootstrap replicates.Gene expression in P. capsici
-
P. capsici mycelia were pre-cultured in 90 mL of 10% V8 medium (150 rpm, 28 °C, 24 h). Co-culture treatment (JK2) received 10 mL pre-prepared JK2 fermentation broth, while control (CK) received 10 mL sterile LB medium. Triplicate cultures were shaken (150 rpm, 28 °C, 48 h), filtered through sterile gauze, and washed thrice with sterile water. Mycelial pellets were blotted dry on filter paper, flash-frozen in liquid nitrogen, and stored at −80 °C. Total RNA was extracted using a Fungal RNA Mini Kit (Omega Bio-Tek, UT, USA) and sequenced on an Illumina HiSeq 2500 platform (Biomarker Technologies Co., Ltd., Beijing, China).
Raw sequencing reads were quality-filtered (Trimmomatic v0.39) and mapped to the P. capsici reference genome (NCBI accession: Phyca11_Mi) using HISAT2 (v2.1.0)[27]. Differentially expressed genes (DEGs) were identified using DESeq2[28] with thresholds of |log2 (fold change)| ≥ 2, p-value ≤ 0.05, and false discovery rate (FDR) < 0.01. Functional annotation of DEGs was performed via Gene Ontology (GO;
www.geneontology.org ) and Kyoto Encyclopedia of Genes and Genomes (KEGG;www.genome.jp/kegg ) enrichment analyses. Ten DEGs were validated by qRT-PCR using the 2−ΔΔCᴛ method with gene-specific primers (Supplementary Table S1).Evaluation of the effect of JK2 strain on pepper blight in vivo
-
JK2 inoculum was prepared by culturing in LB broth (37 °C, 150 rpm, 24 h) and adjusting to 1 × 108 CFU mL−1 (OD600 = 0.4) with sterile water[1]. P. capsici was incubated in 10% sterile V8 liquid culture medium (28 °C, 150 rpm, 4 d)[29], filtered through cheesecloth, and rinsed three times with sterile water to eliminate mycelial fragments. The spore suspension was adjusted to 1 × 105 CFU mL−1 with sterile distilled water using a hemocytometer[22].
Uniform four-leaf pepper seedlings were subjected to four treatments in a greenhouse. CK: 20 mL of LB liquidm edium (control); LJ: 20 mL of JK2 suspension; LP: 20 mL of LB medium + 10 mL of P. capsici spores (24 h post-LB); JP: 20 mL JK2 suspension + 10 mL P. capsici spore suspension (24 h post-JK2). Treatments were applied via root drenching (single application), with three biological replicates and 20 plants per replicate. Disease progression (incidence rate, disease index, and biocontrol efficacy) was monitored at 0, 3, 5, 7, 10, and 14 d post-inoculation using a 5-tier severity scale: Grade 0, plants were healthy with no illness; Grade 1, leaves exhibited slight wilting; Grade 2, stems and leaves slightly wilted with necrotic spots on the leaves; Grade 3, plants moderately wilted with necrotic spots on the leaves and stems; and Grade 4, plants died[30]. Disease index and biocontrol efficacy were calculated according to the following formulas[22,30]: Incidence rate (%) = (Number of diseased plants)/(Total number of investigated plants) × 100%; Disease index = ∑(Number of diseased plants in this grade × Diseased grade)/(Total number of investigated plants × Highest diseased grade) × 100%; Biocontrol efficacy (%) = (Disease index of LP − Disease index of JP)/Disease index of LP × 100%. The H2O2 and MDA contents, and SOD, POD, and CAT activities were determined using reagent kits (Solarbio Science & Technology Co., Ltd, Beijing, China), following the manufacturer's instructions.
Statistical analyses
-
Data variability was assessed using the coefficient of variation (C.V.), calculated as (SD/Mean) × 100%. Values from three independent biological replicates were presented as mean ± standard deviation (SD) alongside C.V. values. One-way ANOVA with variance decomposition was performed using IBM SPSS 21.0 Statistics software to evaluate between-group and within-group variances. Duncan's multiple comparison tests (p < 0.05) was applied to determine significant differences. All figures were drawn using Origin 2024 software.
-
The colony diameter of P. capsici cultured in a medium containing JK2 suspension was reduced to 14.90% of the control group, demonstrating 90.48% inhibition efficiency (Fig. 1a, b).
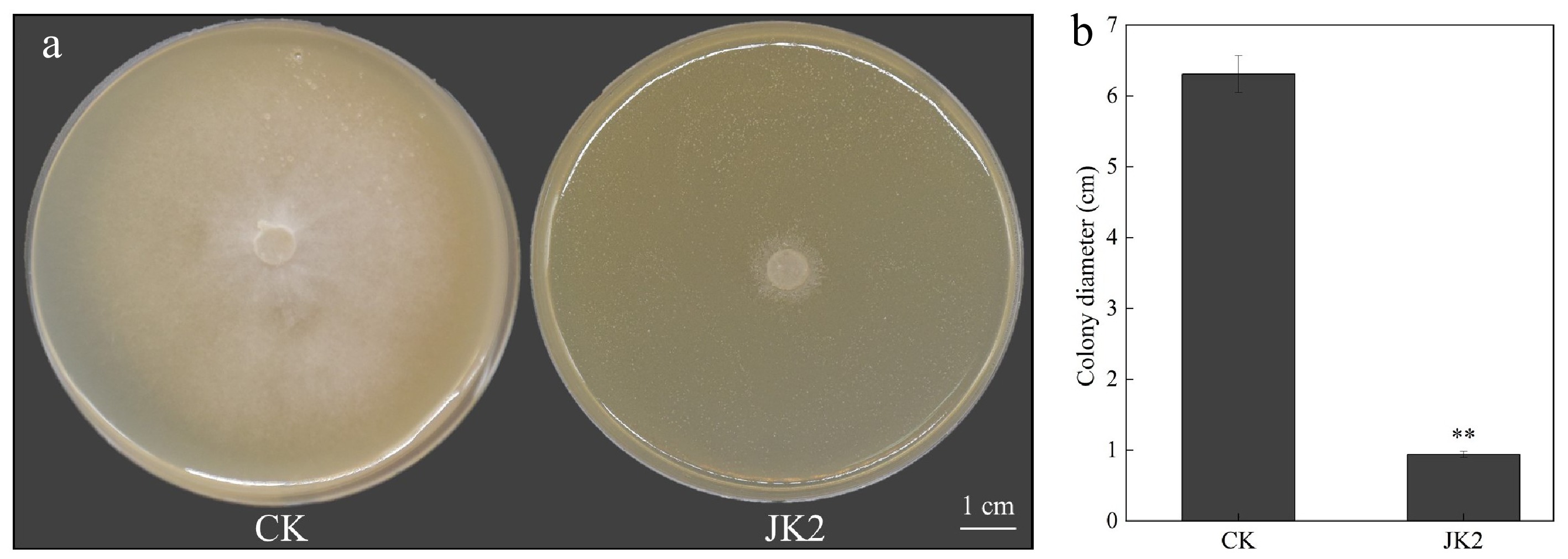
Figure 1.
Effect of strain JK2 on P. capsici in vitro. (a) Antifungal activity of JK2 against P. capsici in vitro. (b) Diameter of P. capsici after treatment with JK2 suspension. All experiments were repeated three times. Data represents means ± SD for three independent replicates. The statistical analyzes were performed using Duncan's multiple comparison (**, 0.001 < p < 0.01).
Morpho-molecular characterization of strain JK2
-
Colonies of JK2 exhibited creamy-white pigmentation (Fig. 2a) with smooth margins during early growth (24 h), transitioning to wrinkled textures in later stages (Fig. 2b). Gram staining confirmed its Gram-positive nature (purple coloration; Fig. 2c), while endospore staining revealed green-fluorescent spores (Fig. 2d). Phylogenetic analysis of the 1,432-bp 16S rDNA sequence demonstrated high similarity to B. amyloliquefaciens (NR 117946.1) (Fig. 2e).
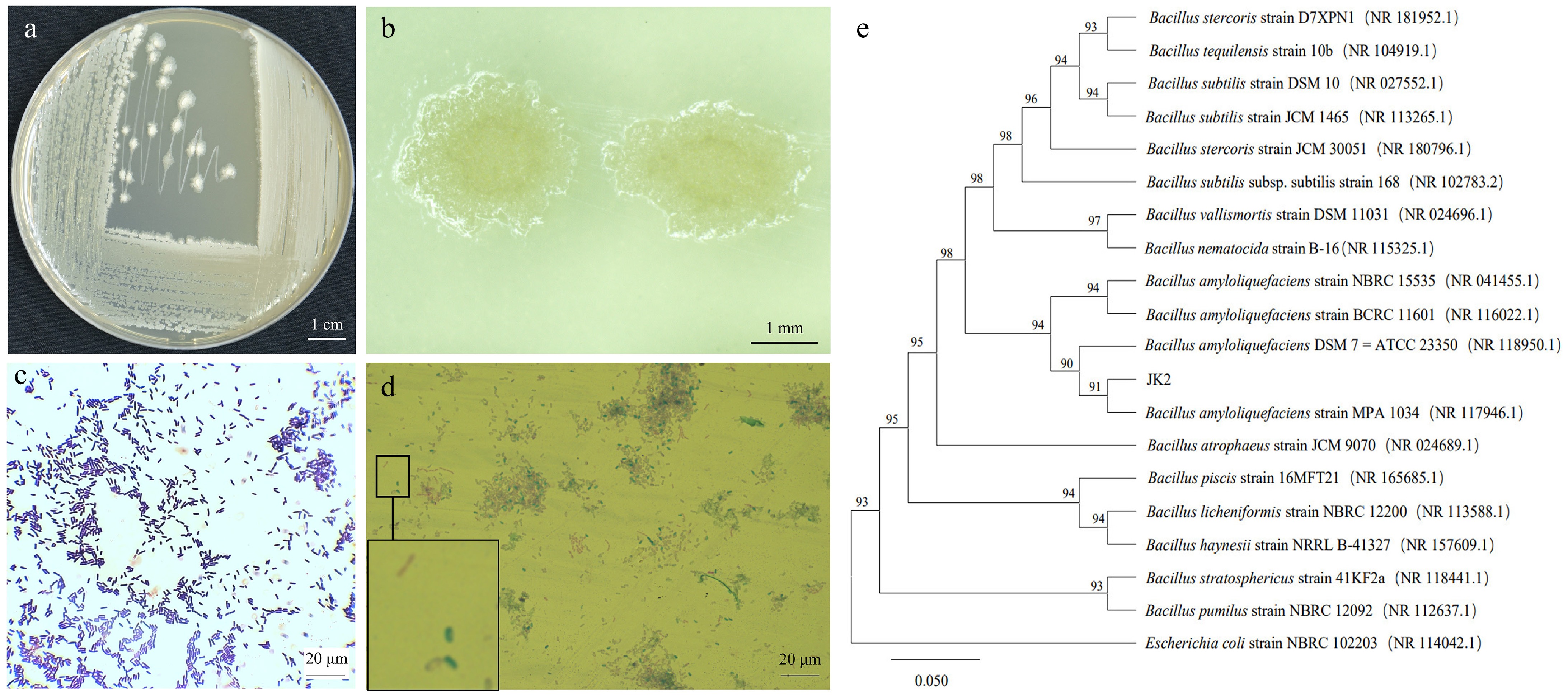
Figure 2.
Morphology and phylogenetic tree of antagonistic bacteria strain JK2. (a) Colony morphology of JK2 on LB medium. (b) Stereoscopic microscopic observation of JK2 single colony. (c) Microscopic observation of JK2 gram staining. (d) Microscopic observation of JK2 spore staining. (e) Phylogenetic tree of JK2 based on 16S rDNA sequence and closely related species constructed by the neighbor-joining method. The number at the nodes indicates the level of bootstrap support (%) based on 1,000 replicates. The scale bar at the bottom indicates the genetic distance. GenBank accession numbers analyzed here are shown in parentheses.
Global transcriptomic changes in P. capsici under JK2 stress
-
Comparative transcriptomics identified 2,464 DEGs (|log2FC| ≥ 2, FDR < 0.01), comprising 1,249 upregulated and 1,215 downregulated genes (Fig. 3a, b). Functional annotation of the top 20 upregulated DEGs revealed enrichment in: Carbohydrate transport/metabolism (25%, 5/20); Energy production/conversion (15%, 3/20); Lipid transport/metabolism (10%, 2/20); Amino acid transport/metabolism (10%, 2/20); and Inorganic ion transport (5%, 1/20). Notably, the top 20 downregulated DEGs included: Eukaryotic initiation factor 3 subunit (11.64-fold downregulation); Cutinase-encoding gene (10.75-fold downregulation); DNA replication/repair-associated gene; Four signal transduction genes (9.89–10.56-fold downregulation) (Supplementary Table S2).
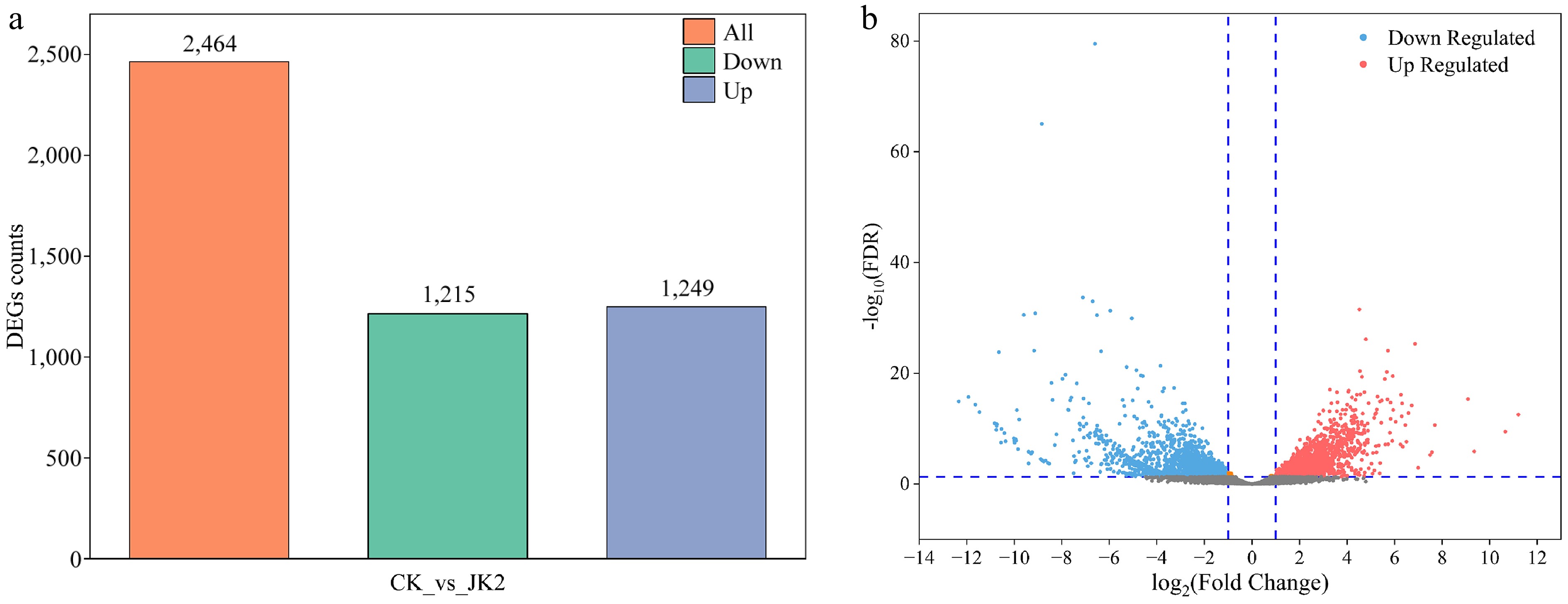
Figure 3.
Overview of transcriptome analysis of P. capsici after application of strain JK2. (a) Bar graph of up- and down-regulated genes from pairwise comparisons. (b) Volcano map of DEGs for CK vs JK2. The red dots represent up-regulated differential genes, the blue dots represent down-regulated differential genes, and the gray dots represent non-differentially expressed genes. CK: P. capsici cultured without strain JK2; JK2: P. capsici co-cultured with strain JK2.
Functional enrichment analysis of DEGs
-
GO functional classification includes three main branches: biological process, molecular function, and cellular component. GO enrichment analysis revealed that DEGs annotated to biological processes had the highest number and widest coverage, with 1,005 and 889 DEGs related to cellular and metabolic processes, respectively. DEGs related to cellular components were enriched only in the cellular anatomical, intracellular, and protein-containing complex pathways. In the molecular function category, 922 and 833 DEGs were associated with binding and catalytic activity, respectively (Fig. 4a). To speculate on the possible functions of the DEGs, the first five terms with small q values in biological processes, molecular functions, and cellular components were used for enrichment analysis. Among the biological processes, DEGs were enriched in ncRNA metabolic (14.30%), ncRNA processing (12.20%), ribosome biogenesis (11.34%), rRNA processing (9.82%), and rRNA metabolic (9.82%) pathways. In the cellular component category, the DEGs were enriched in the nucleolus (14.14%), ribonucleoprotein complex (12.46%), preribosome (7.63%), 90S preribosome (4.15%), and small-subunit processome (3.37%) pathways. In the molecular function category, the DEGs were mainly enriched in the RNA-binding (11.21%) pathway (Fig. 4b).
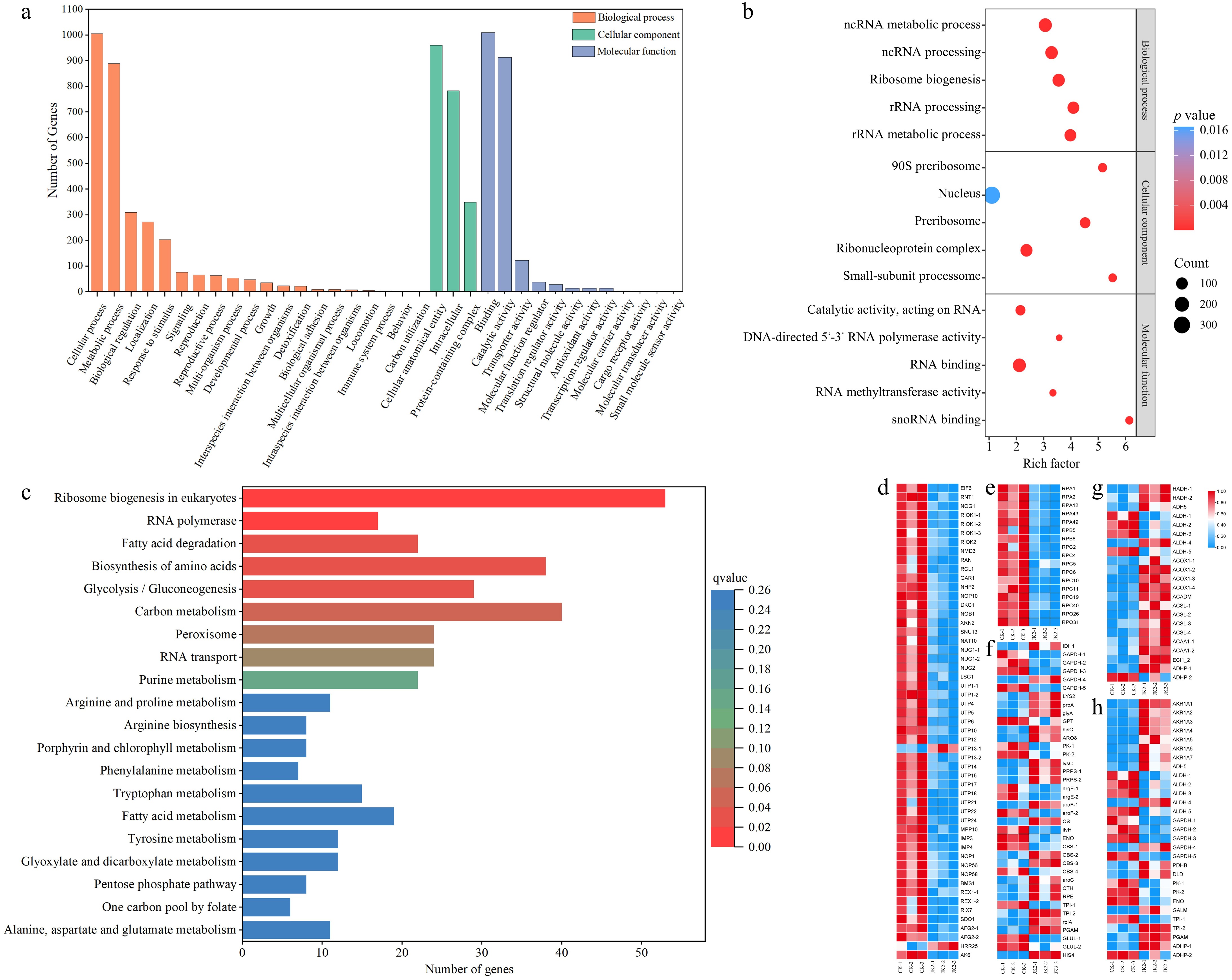
Figure 4.
GO functional classification, KEGG classification of DEGs for CK vs JK2, and Heatmap of the KEGG pathway. (a) Histogram of GO functional classification. The DEGs were assigned to three categories: biological process, cellular component, and molecular function. (b) GO functional classification map of the first five terms with small q value in biological process, molecular function, and cellular component. The color of the points reflects the p value, and the red indicates the more significant enrichment. The size of the dots represents the count of DEGs enriched. (c) Histogram of KEGG classification. (d) Heatmap of DEGs in ribosome biogenesis in eukaryotes pathway (ko03008). (e) Heatmap of DEGs in RNA polymerase pathway (ko03020). (f) Heatmap of DEGs in fatty acid degradation pathway (ko00071). (g) Heatmap of DEGs in amino acid biosynthesis pathway (ko01230). (h) The heatmap of DEGs in glycolysis/gluconeogenesis pathway (ko00010).
The top 20 pathways with low q values were visualized using KEGG enrichment analysis (Fig. 4c). The most prominent of which was ribosome biogenesis in eukaryotes (ko03008), which mapped to 9.45% of the annotated genes, including EIF 6, RNT1, NOG1, and 38 other downregulated genes (Fig. 4d). Seventeen genes annotated to the RNA polymerase pathway (ko03020) were all downregulated in the JK2 group, including five RPA, two RPB, eight RPC, and two RPO genes (Fig. 4e). Following the fatty acid degradation pathway (ko00071), 17 genes (HADH, ADH5, ACOX1, ACADM, ACSL, ACAA1, and ECI1) were upregulated and five genes (four ALDH and one ADHP) were downregulated (Fig. 4f). Additionally, 38 genes were enriched in amino acid biosynthesis (ko01230) (Fig. 4g). In addition, 29 genes were enriched in the glycolysis/gluconeogenesis (ko00010) pathway, and AKR1A, PDHB, DLD, GALM, TPI1-2, PGAM, and ADHP were significantly upregulated (Fig. 4h).
Combined with GO and KEGG enrichment analyses, the expression products of many DEGs were enriched in the categories of 'Genetic information processing' and 'Metabolism' pathways, indicating that strain JK2 inhibited the growth of P. capsici mainly by regulating nutrient metabolism and biological genetic information processing.
qRT-PCR validation of transcriptome data
-
Expression patterns of 10 DEGs (five upregulated, five downregulated) showed strong concordance between RNA-seq and qRT-PCR data, confirming transcriptome reliability (Fig. 5).
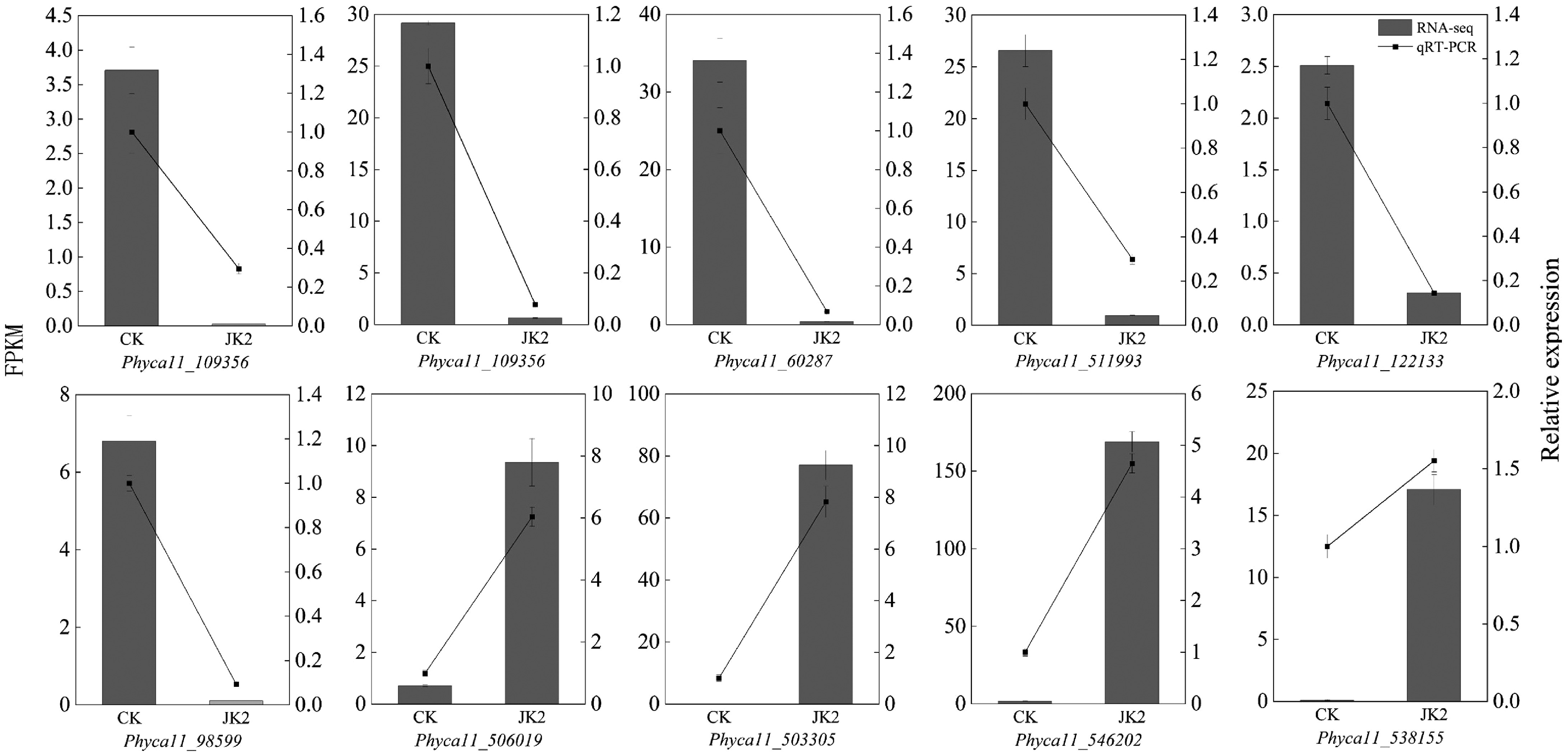
Figure 5.
RT-qPCR validation of DEGs in transcriptome sequencing. The bar graph represents the transcriptome data, and the line graph represents the RT-qPCR results.
Control effect of strain JK2 on pepper phytophthora blight caused by P. capsici
-
The incidence rate and disease index of plants in LP and JP increased exponentially with the days of inoculation with P. capsici; however, those of JP were significantly lower than those of LP at each stage. The incidence rate of LP reached 100% on the 14th day, while the incidence rates of JP were 72.33 ± 8.04% (C.V. = 11.12%), and 83.48 ± 7.25% (C.V. = 8.68%) on the 14th and 18th d, respectively, which were significantly lower than those of LP (Fig. 6a). The disease index of JP (0.55 ± 0.02) was 29.49% lower than LP (0.78 ± 0.06; p < 0.05) (Fig. 6b). JK2 showed high biocontrol efficacy in the early and middle stages, which decreased in later periods. Before the 7th d, the control effect was > 50% (Fig. 6c). During the entire growth period, the pepper plants in CK and LJ grew normally, whereas those in LJ grew better than those in CK. After 5 d of JK2 inoculation, most plants in the LP showed symptoms of disease with wilted leaves. On the 10th d, nearly half of the pepper plants died in the LP, with many leaves turning yellow, whereas minimal plants died in the JP. On the 18th d, all plants in LP died and dried, and most plants in JP died; however, the leaves did not dry, and a small number of plants survived in JP (Fig. 6d). These results indicated that strain JK2 was a potential biocontrol agent for P. capsici.
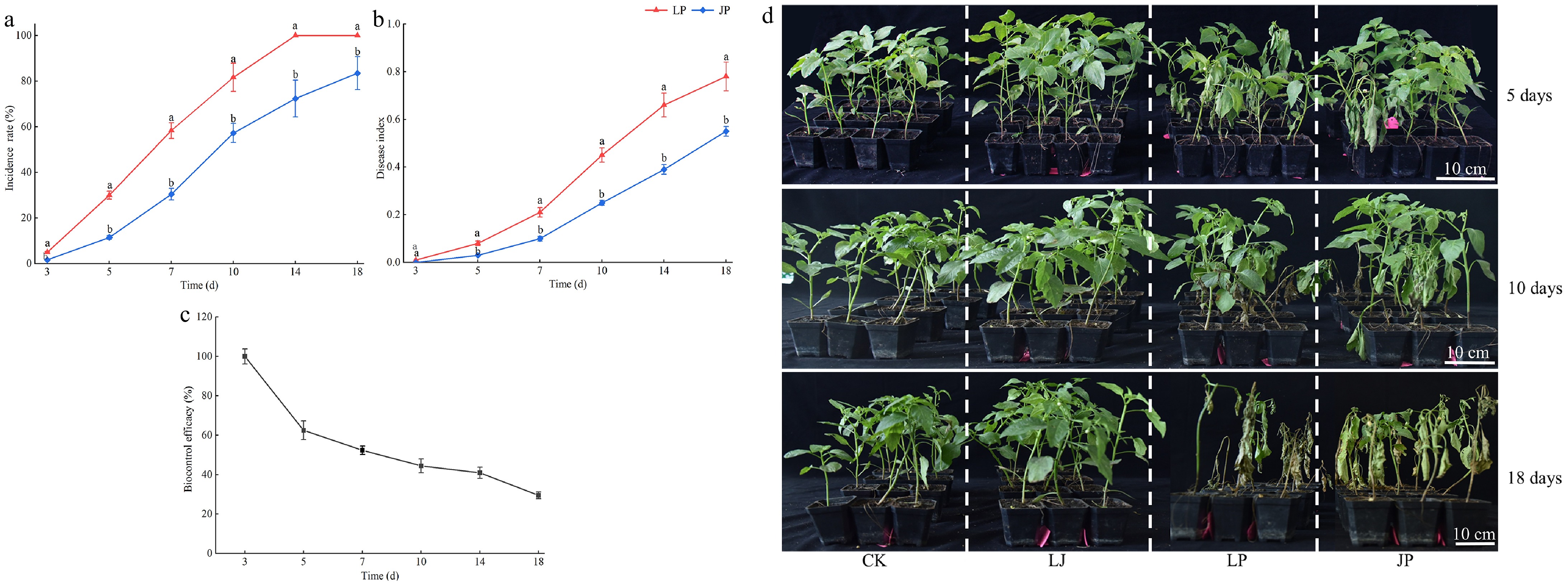
Figure 6.
Control activity of JK2 against P. capsici on pepper seedlings. Twenty mL of JK2 suspension was poured into the roots of the pepper plants, 10 mL P. capsici spore suspension was inoculated 24 h later. (a) Incidence rate of JK2 treated peppers at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (b) Disease index of JK2 treated peppers at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (c) Biocontrol efficacy of JK2 treated peppers at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (d) Disease symptoms on pepper seedlings at 5, 10, and 18 d after inoculation with P. capsici. Data represents means ± SD for three independent replicates. Different lowercase letters in the line chart at the same time indicates significant difference by Duncan's multiple comparison (p < 0.05). CK: Inoculation with 20 mL of LB liquid culture medium; LJ: Inoculation with 20 mL of JK2 suspension; LP: Inoculation with 20 mL of LB liquid culture medium for 24 h, followed by inoculation with 10 mL P. capsici spore suspension; JP: Inoculation with 20 mL JK2 spore suspension for 24 h, followed by inoculation with 10 mL P. capsici spore suspension.
Effect of strain JK2 on H2O2 and MDA contents of pepper plants
-
Under different treatments, H2O2 and MDA contents in pepper leaves gradually increased with growth. CK and LJ without inoculation with P. capsici showed a slow increase in H2O2 and MDA content, and no significant difference was observed between them at each stage (Fig. 7a, b). Inoculation with P. capsici significantly promoted the production of H2O2 and MDA accumulation. H2O2 and MDA contents in LP and JP increased sharply with the extension of inoculation time after day 3; however, H2O2 and MDA contents at each stage were significantly lower than those in LP. At 14 d, the H2O2 (22.57 ± 1.84 nmol·g−1, C.V. = 6.76%), and MDA contents (2.15 ± 0.06 μmol·g−1, C.V. = 2.79%) of JP decreased by 22.66% (p < 0.05) and 12.95% (p < 0.05), respectively, compared with those of LP (Fig. 7a, b). This indicated that the addition of strain JK2 effectively reduced the accumulation of reactive oxygen species (ROS), and the degree of membrane lipid peroxidation induced by P. capsici.
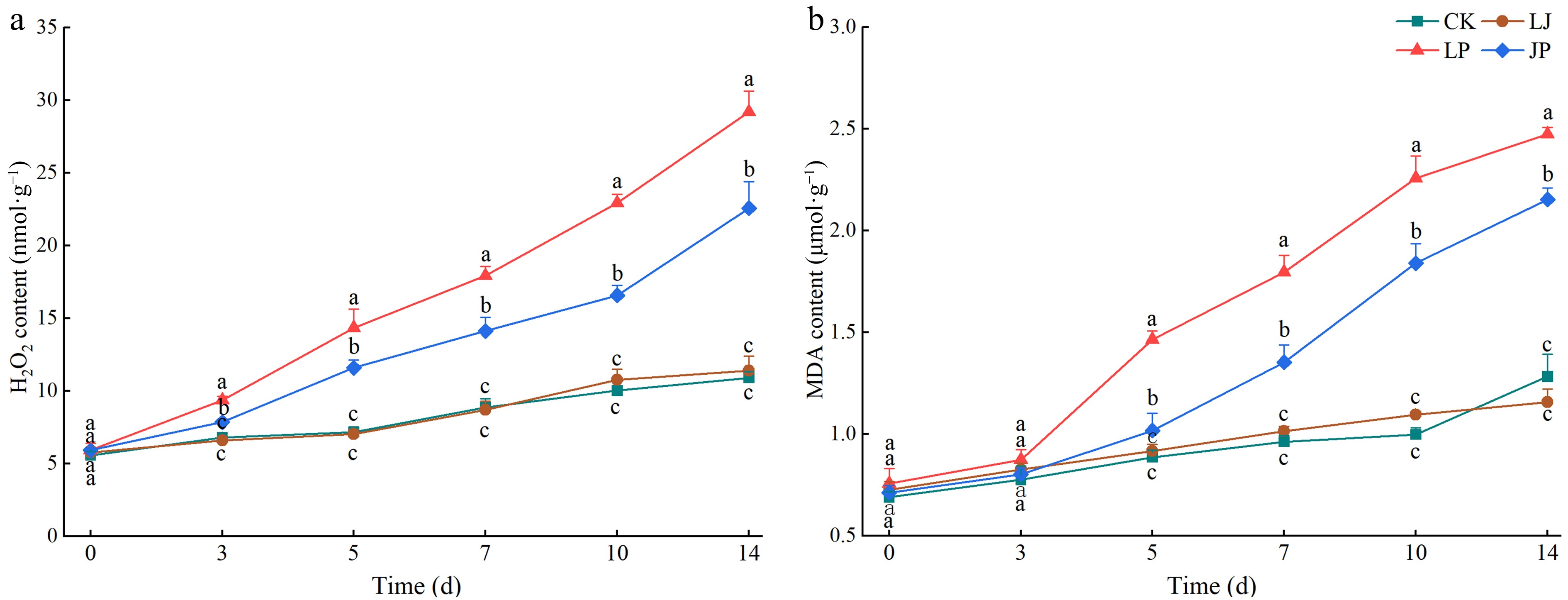
Figure 7.
Effect of JK2 on H2O2 and MDA content in pepper leaves inoculated with P. capsici. (a) H2O2 content at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (b) MDA content at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. Data represents means ± SD for three independent replicates. Different lowercase letters in the line chart at the same time indicates significant differences across treatments by Duncan's multiple comparison (p < 0.05).
Effect of strain JK2 on antioxidant enzyme activity of pepper plants
-
The activities of SOD, POD, and CAT in the CK and LJ plants without P. capsici inoculation increased slowly with pepper growth. No significant differences were observed in the activities of SOD and POD between CK and LJ (Fig. 8a−c). The SOD, POD, and CAT activities in LP and JP inoculated with P. capsici first increased and then decreased. During the early stages of infection with P. capsici, SOD, POD, and CAT activities in LP leaves rapidly increased and peaked on the 5th, 7th, and 5th d, respectively. Subsequently, owing to disease outbreaks and plant death, antioxidant enzyme activities declined rapidly and continuously. On the 14th day, as most plants in LP had died, SOD (65.67 ± 4.76 U·g−1, C.V. = 7.25%), POD (7.64 ± 0.04 U·g−1, C.V. = 0.52%), and CAT (151.68±10.26 U·g−1, C.V. = 6.76%) activities were 73.37% (p < 0.05), 61.59% (p < 0.05), and 73.59% (p < 0.05) lower than those in LJ, respectively (Fig. 8a−c). The addition of JK2 delayed the peak antioxidant enzyme activity by 2–3 d, and increased the time and ability of antioxidant enzymes to clear ROS. Although the enzymatic activity of JP decreased in the later period of P. capsici infection, it was significantly higher than that of LP during the same period. Specifically, on the 14th d, SOD activity in JP (14.77 ± 1.04 U·g−1, C.V. = 7.04%) was 159.39% higher than in LP (65.67 ± 4.76 U·g−1, C.V. = 7.25%). Similarly, POD (JP: 14.77 ± 1.04 U·g−1, C.V. = 7.04%; LP: 7.64 ± 0.04 U·g−1, C.V. = 0.52%) and CAT (JP: 878.52 ± 69.42 U·g−1, C.V. = 7.90%; LP: 151.68 ± 10.26 U·g−1, C.V. = 6.76%) activities showed significant increases (p < 0.05) (Fig. 8a−c).
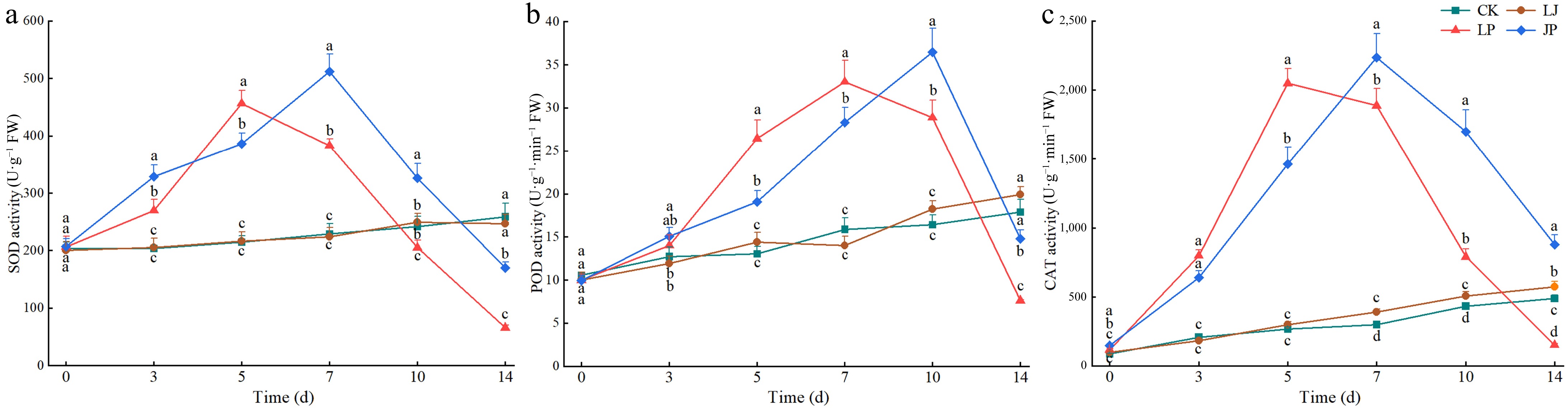
Figure 8.
Effect of antioxidant enzyme activities in pepper leaves inoculated with P. capsici. (a) SOD activity at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (b) POD activity at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. (c) CAT activity at 3, 5, 7, 10, 14, and 18 d after inoculation with P. capsici. Data represents means ± SD for three independent replicates. Different lowercase letters in the line chart at the same time indicates significant differences across treatments by Duncan's multiple comparison (p < 0.05).
-
The escalating demand for sustainable alternatives to chemical pesticides has driven significant interest in microbial biocontrol agents[31]. Among these, Bacillus spp. emerge as predominant antagonists, with B. amyloliquefaciens being extensively validated for its broad-spectrum pathogen suppression[16,32,33]. This bacterium competitively colonizes phytopathogen niches, secreting surfactins to enhance biofilm formation and resource competition[21,34]. Beyond lipopeptides and cyclic surfactins, B. amyloliquefaciens produces lytic enzymes (e.g., amylases, proteases) that directly degrade pathogen structures[35]. In the present study, the antagonistic bacterial strain JK2 was identified as Bacillus amyloliquefaciens, with an in vitro inhibition efficiency of 90.48% against P. capsici, consistent with reported efficacies of B. amyloliquefacienss strains PsL[34], and IBFCBF-1[22].
Biofilm formation, a key resistance mechanism under biotic stress[36], comprises protein-rich matrices with minor carbohydrate, lipid, and nucleic acid components[37]. During biofilm formation, many macromolecular levels are typically higher in biofilms than in planktonic cells[38]. Transcriptomic analysis revealed JK2-induced enrichment of P. capsici DEGs in fatty acid degradation, amino acid biosynthesis, and central carbon metabolism, partially aligning with Colletotrichum gloeosporioides metabolic responses to Bacillus stress[39]. However, JK2 uniquely upregulated amino acid (arginine, proline, phenylalanine) synthesis and metabolism. Besides, contrasting with B. amyloliquefaciens JDF3's suppression of nucleotide sugar metabolism in Phytophthora sojae[40], in the present study, the DEGs related to carbohydrate, amino acid, lipid, and inorganic ion transport and metabolism were upregulated. We hypothesized that JK2-triggered metabolic dysregulation compromises membrane integrity, forcing P. capsici to overinvest in biofilm stabilization.
RNA polymerases, molecular motor enzymes that are responsible for the synthesis of RNA from DNA templates, facilitate transcription and control gene expression in all living organisms[41]. During transcription, RNA polymerases synthesize a variety of RNA molecules in the nucleus, including single polycistronic rRNA precursors for ribosomal subunit synthesis, and mRNA precursors for protein synthesis[42]. Given their critical roles, damage to RNA polymerases can disrupt essential cellular processes, resulting in impaired cell growth and development, and even death[43]. In addition, the ribosome is a core molecular machine that translates messenger RNA (mRNA) to protein, which plays a key role in gene expression in all organisms. Thus, ribosomes determine protein synthesis in all cells[44]. In the present study, the significant downregulation of RNA polymerase (e.g., RPB1, RPC1) and ribosome biogenesis genes (e.g., EIF3, NOG1) in P. capsici under JK2 stress aligns with prior reports where Bacillus spp. disrupted pathogen transcription and translation[40,45]. Unlike B. velezensis FZB42, which targets pectate lyases[45], JK2 uniquely suppresses ribosomal pathways, suggesting strain-specific modes of action. Furthermore, the upregulation of fatty acid degradation genes (ACOX1, ECI1) contrasts with findings in Colletotrichum gloeosporioides, where metabolic shifts favored pathogen survival[39]. This divergence implies that JK2's metabolic interference may overwhelm pathogen adaptation, a hypothesis supported by the rapid decline in P. capsici viability. These findings challenge the assumption that metabolic upregulation universally aids pathogen resistance, highlighting JK2's efficacy in destabilizing critical biosynthetic pathways.
Recent studies have identified multiple microbial agents demonstrating potent in vitro suppression of P. capsici, including Streptomyces albus[10], B. licheniformis[46], and B. amyloliquefaciens[22]. Our pot experiments revealed that strain JK2 achieved 62.50% and 52.38% disease control efficacy at 5 and 7 d post-inoculation, respectively, compared to untreated controls. This temporal efficacy profile indicates JK2 exerts early-intermediate stage suppression, effectively delaying symptom onset—a critical advantage for field applications. Comparatively, B. amyloliquefaciens JDF3 demonstrated 70.7% control against soybean Phytophthora blight[40], while strain PsL exhibited > 90% suppression in soil-infested sweet pepper systems[34], suggesting niche-specific performance variations among Bacillus biocontrol strains.
Pathogen invasion disrupts cellular redox equilibrium through excessive ROS generation, leading to membrane lipid peroxidation and tissue damage[47]. Plant countermeasures involve enzymatic ROS-scavenging systems, notably superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT), which detoxify H2O2 into water and oxygen[48−50]. The biocontrol effects of B. amyloliquefaciens antagonistic strains occur partly through the induction of resistant ISR in the host plant, including regulating the expression of defense genes, reducing the accumulation of ROS, and promoting the antioxidant system in host plants[21,51]. In the present study, the inoculation of pepper plants with JK2 delayed the accumulation of H2O2 and MDA, and the content at all time points was significantly lower than that in the treatment with only P. capsici. Strain JK2 also increased the levels of antioxidant oxidases SOD, POD, and CAT in the middle and later stages of P. capsici infection. These findings align with B. amyloliquefaciens L-1 induced POD and CAT activation, as well as MDA declined accumulation in Botryosphaeria berengeriana infected pears[52], reinforcing the genus' conserved role in ROS management. The dual mechanism, direct pathogen suppression coupled with host antioxidant priming, positions JK2 as a multifaceted biocontrol agent. Future field validations should assess its translational potential under agronomic conditions.
-
In this study, we evaluated the inhibitory effects of B. amyloliquefaciens JK2 on the growth of P. capsici and further assessed its biocontrol potential against Phytophthora blight in pepper plants. Bacillus amyloliquefaciens JK2 achieves 90.48% inhibition of P. capsici by disrupting ribosomal biogenesis (ko03008) and RNA polymerase activity (ko03020). JK2 treatment reduces pepper blight incidence by 27.67% and delays disease progression via ROS scavenging (SOD, POD, CAT enhancement). JK2's dual action-direct pathogen suppression and host immunity priming-provides a sustainable alternative to chemical fungicides.
This work was supported by the Chengdu Science and Technology Project (2023-YF09-00086-SN), and Sichuan Province Science and Technology Planning Project (2023JDRC0044, 2023JDRC0111).
-
The authors confirm contribution to the paper as follows: study conception and design: Xie Y, Liu J; data collection: Gu F, Lu Q, Wu Z, Guo Q, Zhu J; analysis and interpretation of results: Xie Y, Zhang Y, Yu X; draft manuscript preparation: Xie Y, Zhang Y, Chen M; review and editing, supervision: Tang Y, Liu J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primer sequences of qRT-PCR.
- Supplementary Table S2 The top 20 upregulated and downregulated DEGs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xie Y, Zhang Y, Gu F, Lu Q, Yu X, et al. 2025. Bacillus amyloliquefaciens JK2 suppresses Phytophthora capsici via genetic information disruption and ROS modulation in pepper plants. Technology in Horticulture 5: e028 doi: 10.48130/tihort-0025-0023 

Bacillus amyloliquefaciens JK2 suppresses Phytophthora capsici via genetic information disruption and ROS modulation in pepper plants
- Received: 11 February 2025
- Revised: 13 May 2025
- Accepted: 30 May 2025
- Published online: 04 August 2025
Abstract: Phytophthora capsici, a devastating soil-borne oomycete pathogen, poses a significant threat to global pepper production. Biocontrol using antagonistic bacteria offers a sustainable alternative to chemical fungicides. In this study, Bacillus amyloliquefaciens JK2, isolated from peach, exhibited 90.48% in vitro inhibition against P. capsici. Transcriptomic analysis identified 2,464 differentially expressed genes (DEGs) in P. capsici under JK2 co-culture, with downregulation in ribosome biogenesis and RNA polymerase pathways, and upregulation in fatty acid degradation and amino acid biosynthesis. These findings suggest that JK2 disrupts RNA synthesis and protein translation, thereby inhibiting the growth of pathogens. Pot experiments demonstrated that JK2 reduced disease incidence by 27.67%, and delayed hydrogen peroxide (H2O2) and malondialdehyde (MDA) accumulation, while enhancing superoxide dismutase (SOD, 159.39%), peroxidase (POD, 93.37%), and catalase (CAT, 479.19%) activities in pepper plants. Our results highlight JK2's dual role in directly suppressing P. capsici and priming host antioxidant defenses, positioning it as a promising biocontrol agent for sustainable agriculture.