









-
The eyes serve as a crucial sensory organ for humans to acquire external information. The retina plays a significant role, acting as the sole source of visual information. Upon exposure to light stimuli, photoreceptor cells (PRs) in the retina undergo a series of photochemical changes and potential alterations. Horizontal cells (HCs) initially modulate the electrical signals, which then proceed to bipolar cells (BCs) for further processing. During this phase, modulation occurs from amacrine cells (ACs). Ultimately, retinal ganglion cells (RGCs) receive signals from the BCs, integrating them and converting them into neural impulses. These impulses travel through the axons of the RGCs to the lateral geniculate nucleus of the brain. After being relayed through the lateral geniculate nucleus, the signals reach the cerebral cortex, particularly the visual cortex, for final processing and interpretation of information. The PRs lack regenerative abilities. Damage to the retina often results in vision impairment or even irreversible blindness. This serves as a primary cause for retinal degenerative diseases (RDDs) such as age-related macular degeneration (AMD) and retinitis pigmentosa (RP).
With the global population growth and changes in age demographics, the epidemiological characteristics of RDDs exhibit significant variations across different countries and regions. Currently, AMD constitutes a substantial proportion of incurable blindness cases worldwide[1]. AMD is more prevalent in developed nations, with approximately 11.64% (around 18.34 million individuals) of the population aged over 40 in the United States affected by early AMD, while the prevalence of late AMD is about 0.94%[2]. RP is influenced by both genetic and environmental factors, displaying a notable familial inheritance pattern, with incidence rates varying by region; the global average incidence is approximately 1 in 4,000, with the onset typically occurring during adolescence[3,4]. Diabetic retinopathy (DR), a common complication among diabetic patients, is becoming increasingly prevalent alongside rising diabetes rates. As of 2020, over 3.9 million individuals globally were affected by DR, with higher concentrations observed in countries such as the United States, China, India, Brazil, and Saudi Arabia[5].
RDDs are chronic progressive conditions characterized by the gradual apoptosis of retinal cells, leading to a progressive decline in visual acuity for patients. This process not only severely impacts the daily lives of the affected individuals but may also trigger psychological issues such as anxiety and depression. However, these diseases exhibit high heterogeneity, with complex pathophysiological mechanisms and, in most cases, result in irreversible blindness[6,7]. The current therapeutic approaches can slow disease progression to some extent, but they remain unable to halt the continuous deterioration of vision until complete blindness occurs. Vision impairment significantly impacts individuals' economic, health, education, and daily lives. The rising number of people experiencing vision loss poses a serious public health burden for any nation. As early as 2020, vision impairment had already caused a global loss in productivity, with estimates putting this figure as high as
${\$} $ -
Organoids are three-dimensional (3D) structures that self-organize through interactions between cells and the extracellular matrix. They can originate from stem cells or adult tissues. Compared with two-dimensional (2D) cultures, organoids incorporate tissue architecture and exhibit some functional capabilities[9]. For over a decade, researchers have made significant efforts to identify and activate the crucial components that drive cellular differentiation into retinal neurons. They have developed 3D retinal organoids (ROs) enriched with natural retinal cell constituents. ROs derived from human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs) enhance our understanding of retinal development. These models support cell transplantation therapies, drug screening, and the development of disease models, offering promising prospects for treating RDDs.
In 2011, Prof. Sasai Yoshiki's group cultured bone marrow mesenchymal stem cells (mESCs) in suspension with Matrigel to obtain embryoid-like bodies (EBs) and induced the spontaneous formation of RPCs, which aggregated and invaginated in optic cup-like structures with a proximal–distal pattern, with the proximal portion differentiating into retinal pigment epithelium and the distal portion folding inward into an embryoid-like shape, exhibiting interdynamic nuclei and giving rise to layered neural retinal tissues[10]. Subsequently, they optimized the experimental protocol further. They initiated a SFEBq (serum-free floating culture of EB-like aggregates with quick reaggregation)[11] culture using hESCs that were reaggregated in low-cell-adhesion V-type 96-well plates, forming EBs. Under the influence of inducing agents, the EBs developed into optic cups. These were then maintained in the suspension culture, maturing into layered ROs. Although the optic cup structures were generated in both mESCs and hESCs, there were some interesting differences between them, with hESCs taking longer to culture and generating early optic cups that were much larger than those generated by mESCs and with thicker derived neural retinas[12].
The strategies for inducing ROs are continually improving and optimizing, bringing their structure closer to maturity. In addition to utilizing SFEBq technology for inducing cup-like structures in suspension cultures, several other methods have also successfully generated ROs. Zhong et al.[13] formed EBs using the suspension aggregation technique and hiPSCs, followed by attachment culture on Matrigel. In the fourth week, the outgrowing neural retina tissue was detached and continued in long-term culture under suspension conditions. By employing a similar scraping technique, Regent et al.[14] observed the formation of optic nerve vesicle (OV) structures during a 20- to 30-day cultivation period. Some methods first culture hESC or hiPSC aggregates in suspension to form cysts. After adhering, these cysts produce monolayer cell sheets, which spontaneously develop into ROs through enzymatic dissociation[15,16]. The neuroectoderm can be directly induced for directional differentiation in a flat system, eliminating the need for EB generation. Subsequently, mechanical enrichment of retina-like structures occurs, followed by suspension culture[17,18]. Overall, the development of ROs is a self-directed, prolonged process that typically begins at an early stage, featuring the formation of OV-containing RPCs.
RO technology has emerged as a crucial tool for studying retinal development, disease mechanisms, and drug screening (see Fig. 1). Before alternative therapies were developed, organoids were initially employed to create in vitro models of diseases. This approach aimed to enhance our understanding of disease mechanisms and evaluate potential treatments. Prof. Jin's research group utilized the Sendai virus to induce nonintegrating induced pluripotent stem cells (iPSCs) in the fibroblasts from patients. They aimed to develop patient-specific rod cell models to study RP caused by Rgo mutations. Rod cells derived from patients' iPSCs exhibited classic endoplasmic reticulum stress characteristics and replicated the disease phenotype in vitro[19]. Michael A. Dyer's research group differentiated iPSCs carrying genetic mutations associated with retinoblastoma (Rb) into ROs. They then transplanted these organoids into the eyes of mice, effectively creating a model for Rb[20]. Compared with traditional animal models and 2D cell culture techniques, human organoid systems offer models that closely mimic human physiological and pathological conditions in drug research while also reducing reliance on animals and lowering development costs. By establishing a late-stage retinoblastoma organoid model, researchers assessed the impact of drugs on cell cycle disruption and genotoxic effects. The combined treatmentof melphalan and topotecan effectively targeted tumor conecells within the organoids[21]. This organoid model offers a valuable opportunity for in vitro screening of drug accessibility and sensitivity.
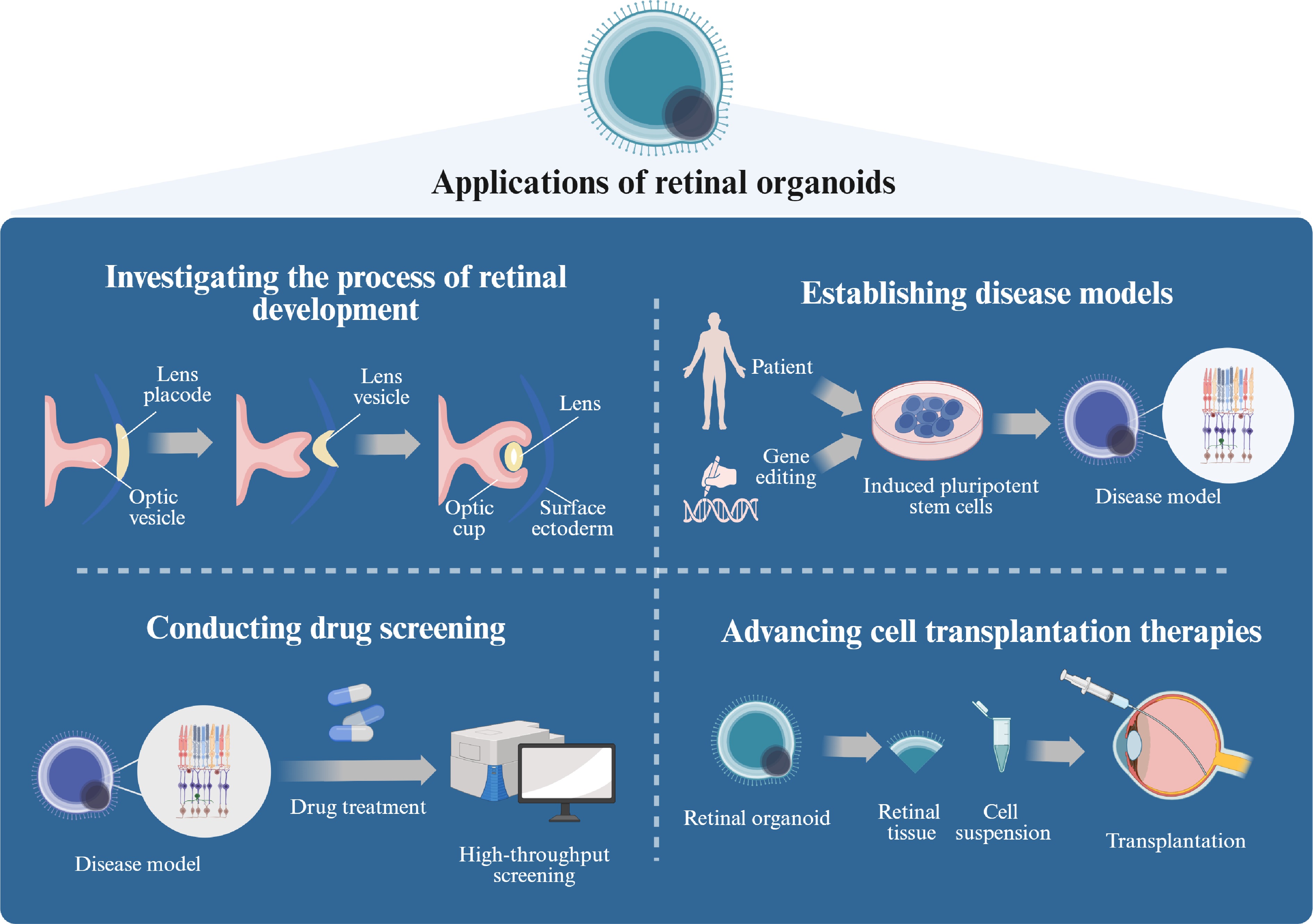
Figure 1.
The application of ROs (created with BioRender,
www.biorender.com ). Retinal organoids demonstrate significant potential across multiple research domains, including developmental biology, modeling disease pathophysiology, therapeutic discovery, and regenerative medicine applications. (1) These 3D in vitro models recapitulate the spatiotemporal progression of retinogenesis from embryonic progenitor commitment to stratified tissue maturation, enabling systematic investigation of molecular regulators and cellular dynamics during neuroretinal development. (2) Disease-specific models can be established through either derivation from patient-specific hiPSCs or CRISPR/Cas9-mediated genome editing, allowing precise replication of inherited retinal disorder phenotypes. (3) Their structural complexity and scalability serve as a high-throughput screening platform for pharmacological compounds, facilitating quantitative assessment of therapeutic efficacy and developmental toxicity. (4) From a translational perspective, retinal organoids represent a promising autologous cell source for PR replacement therapies targeting degenerative retinopathies. -
Current therapeutic strategies for RDDs primarily target early-stage pathologies through gene therapy, pharmacotherapy, and optogenetic modulation, with the principal objective of preserving the residual PRs. However, they show limited efficacy in cases with significant loss of PR in later stages[22]. The clinical and genetic heterogeneity of RDDs, coupled with an incomplete understanding of the apoptotic mechanisms underlying PR degeneration, poses significant challenges for targeted drug development. In contrast, cell-based therapies offer a dual therapeutic paradigm by replacing lost PRs and providing neuroprotective support, thereby emerging as a universalizable strategy across diverse RDD subtypes.
PR transplantation has been systematically investigated since the 1990s, when the initial evidence of donor PR integration into the host's outer nuclear layers (ONLs) was documented. Seminal studies established foundational principles. Cerro et al.[23] demonstrated that neonatal rat retinal cell transplantation in phototoxic retinopathy models resulted in mature rod morphogenesis and synaptic integration within 30 days. Gouras et al.[24] reported graft persistence for at least three months in Royal College of Surgeons (RCS) rats with complete endogenous PR degeneration. Kwan et al.[25] observed transplanted retinal progenitors differentiating into functional PRs exhibiting outer segment formation, RGCs, and BCs. In clinical studies, after transplantation of RPC layers and RPE, 7 out of 10 patients with retinal degeneration (RD) showed some degree of visual improvement and no rejection[26].
Transplanted cells can form visual networks with the host and secrete protective factors to sustain the remaining PRs' survival, further supporting the restoration of photoreceptive function[27]. Numerous research teams have reported that RNA, protein, or organelle exchange between the donor and host can rescue diseased PRs during their integration with retinal receptors, with transfer occurring solely in PR–PR and Müller glial cell (MGC)–PR interactions[28−34].
Despite research indicating that material exchange between PRs is species-dependent, positive outcomes from interspecies transplants suggest that restoring vision by reconstructing visual structures is feasible[35]. In the RCS rat, transplantation of retinal progenitor cells (RPCs) or retinal pigment epithelium (RPE) cells induced visual activity in a portion of the superior colliculus (SC) corresponding to the transplanted area[36]. Material exchange may be a mechanism involved in cell therapy but it is not necessarily the primary factor. Endogenous cell transplantation is feasible for compensating for the lack of PRs; however, in humans, healthy retinas must be obtained from mid-gestation fetuses, which raises a number of ethical, cell source, and supply issues that have hindered the development of related research. Consequently, there is a need to generate large numbers of transplantable PRs in vitro. Autologous cell transplantation represents a promising approach to compensate for the loss of PRs. However, current efforts to obtain healthy human retinal cells face ethical controversies, sourcing issues, and supply limitations, severely hindering the progression of related research.
Human embryonic stem cells (hESCs)[37] and hiPSCs[38] offer potential for the generation of a vast supply of cellular resources that can be expanded in vitro in large numbers and have the capacity to differentiate into different cell types. In vitro differentiation techniques have demonstrated the capacity of hESCs and hiPSCs to undergo a stepwise differentiation process towards the neural retina[14,39−41]. The integration and survivability of retinal neurons derived from embryonic stem cells (ESCs)/iPSCs have been demonstrated in rodent, nonprimate, and other animal models[29,42]. Although stem cell therapy has been put into clinical research, the PR cells produced by this 2D differentiation still have certain gaps compared with in vivo differentiation, such as the absence of the outer segmental structure of mature PRs and the difficulty in forming the structure of the neural retinal epithelial cell layer[43]. To address these limitations, 3D retinal organoid systems enable spatiotemporal PR maturation with outer segment morphogenesis, stratified retinal lamination, and high-throughput production of transplantable PRs, positioning organoid-based therapy as a viable strategy for reconstructing visual circuitry in advanced RDDs with concurrent PR/BC loss.
-
For cases of retinal cell loss due to disease or injury, ROs present a potential source for cell replacement. Transplanted cells may integrate into the host's retinal tissue and restore visual function (see Fig. 2). The potential of RO transplantation therapy is significant, but to achieve widespread clinical application, challenges related to technology, ethics, and regulations must be addressed.
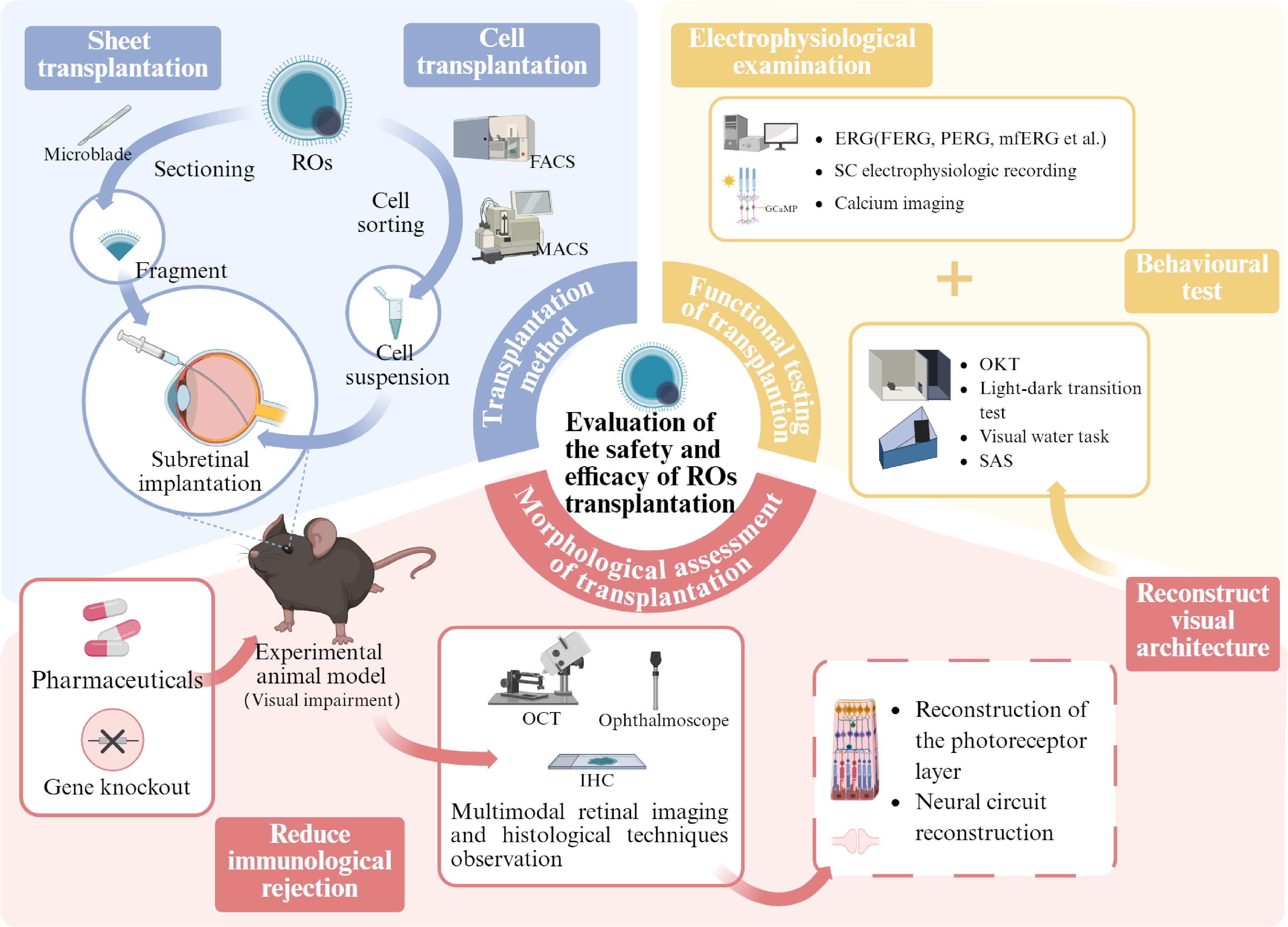
Figure 2.
Retinal organoids: evaluating the safety and efficacy of transplantation (created with BioRender,
www.biorender.com ). In order to advance retinal organoid transplantation therapies toward clinical application, their safety and efficacy must be thoroughly evaluated. Currently, there are two main transplantation methods: lamellar transplantation, in which the organoid is trimmed to a suitable size, and cell suspension transplantation, in which the target cells are isolated from the organoid. Researchers deliver these grafts to the subretinal cavity of the disease model animals using specific tools. In order to reduce immune rejection and increase the survival rate of transplanted cells, researchers have established immunodeficient animal models or used pharmacological treatment protocols. At the same time, multimodal retinal imaging and histologic techniques are used to monitor the contact between the transplanted cells and the host's retina. These assays are designed to assess whether transplantation treatments are successful in reestablishing visual structures, focusing on two key aspects: reconstruction of the PR layer and reconstruction of the neural circuit, i.e., the formation of synaptic connections. In addition, the combined assessment of behavioral tests and electrophysiology can further validate the effective reconstruction of visual structures and provide evidence of improved visual function. OCT, optical coherence tomography; IHC, immunohistochemistry; OKT, optokinetic tracking; SAS, shuttle avoidance behavioral experiment; FERG, flash electroretinogram; PERG, patern electroretinogram.Transplantation methods
-
RO transplantation falls into two categories: layered grafting and cell suspension grafting. The method of delivery significantly influences the polarization of the graft within the host. Layered grafting tends to be more complex, often requiring custom-designed delivery tools depending on the experimental conditions. Typically, researchers trim cultured 3D ROs into millimeter-sized slices using a microblade. They preserve these slices in ice-cold Hawk's balanced salt solution (HBSS) buffer. Using a glass micropipette with an approximate tip diameter of 500 μm or other implant instruments, surgeons deposit the graft into the retinal space of anesthetized animals through a small incision on the sclera or a posterior approach, all under the guidance of an operating microscope. In large animal models, given their anatomically elongated ocular axes and expansive retinal surface areas, intact retinal organoids are preferentially transplanted en bloc over dissected fragments. This surgical strategy preserves structural integrity while achieving the therapeutic PR cell densities required for functional retinal repair[44]. The complexity of surgical delivery and the diminutive size of ROs restrict the large-scale transplantation of RO sheets, which may impose certain limitations on the restoration of visual function.
While 2D PRs demonstrate limited translational potential for cell replacement therapy due to suboptimal differentiation efficiency and incomplete maturation[43], 3D retinal organoids offer distinct advantages. By recapitulating key aspects of the native retinal niche through precise cellular organization and microenvironmental signaling, these organotypic cultures facilitate robust PR lineage specification and terminal differentiation, thereby generating functionally competent PRs at therapeutic scales[45]. Cell suspension transplantation requires purification steps to eliminate unwanted heterogeneous and undifferentiated cells. First, 0.25% trypsin or papain is used to dissociate ROs. The dissociated cells are collected, then the researchersapply fluorescence-activated cell sorting (FACS) or magnetic-activated cell separation (MACS) to isolate target retinal neurons using specific markers. The cells are suspended in sterile HBSS with 0.05% DNase. After anesthetizing the animal, the researcher inserts the injection needle through the superior equatorial sclera under microscopy to inject the cells between the RPE and the neural retina. At this point, the suspension inoculation volume is approximately 200,000 cells. Considering the limited subretinal space, typically, 1–2 μL of the buffer is injected. Excessive cell inoculation leads to high viscosity, making it difficult to draw from the fine injection needle. Common specific cell surface markers, such as CD24, CD73, and CD133, can be used to isolate PRs. In a mixed retinal cell population, CD24/CD73 indicates effective separation of the highly transplantable post-mitotic PR population[46]. However, the biosafety profiles and chemical stability of these sorting markers remain to be systematically validated, necessitating the development of more robust PR isolation methodologies with improved specificity[47].
For cell transplantation, early disease stages show that donor cell suspensions are effective due to their ability to integrate and support the existing structures of the remaining host PRs. In the Cpfl1 murine model characterized by cone-specific dysfunction with preserved rod PR viability, human retinal organoid (hRO)-derived PRs demonstrated functional integration and synaptic connectivity with the host's retinal circuitry[48]. Lamba et al.[49] demonstrated that transplantation of hESC-derived PRs into the subretinal space of neonatal Crx–/– mice resulted in donor cell differentiation into functional PRs and restoration of electroretinographic (ERG) responses within 2–3 weeks. However, the degenerative retinal microenvironment during end-stage retinal degeneration, marked by extensive PR and RPE cell death, significantly compromises the donor cells' survival. In Rd9 murine retinas, integrated rod precursor survival rates exhibited a time-dependent decline post-transplantation[50]. These findings underscore two critical limitations of PR suspension transplantation using RO-derived cells: (1) restricted patient eligibility criteria and (2) transient therapeutic efficacy, as standalone cell replacement fails to arrest the progression of advanced degenerative pathology. Although late-stage disease models including RCS rats[51], Pde6brd1 mice[52], and Rd1 mice[53] exhibited transient visual improvement (1–4 months post-transplantation), longitudinal data confirming sustained therapeutic benefits remain absent. Furthermore, gravitational sedimentation adversely affects transplanted cell polarity, impeding uniform spatial distribution within target regions[54].
PR integration critically depends on structural guidance from host's retinal laminae[55,56]. Lamellar retinal transplantation preserves the organoid's cytoarchitecture, leveraging intrinsic cell–cell interactions to support PRs' maturation and signal transduction, thus emerging as a superior strategy for cell replacement therapy[57]. Human pluripotent stem cell (hPSC)-derived retinal laminae exhibit low immunogenicity[58] and demonstrate prolonged survival in end-stage degenerative retinas, with transplanted PRs developing ultrastructurally intact outer segments (OSs) and functional phototransduction capacity[59,60]. While endogenous bipolar cells within grafts may impede synaptic connectivity between the donor and host PRs[61], CRISPR/Cas9-mediated bipolar cell depletion enhances synaptogenesis and the fidelity of the RGC response[62,63].
Survival of grafts in the host eye
Selecting the optimal grafting stage
-
The culture of retinal organoids recapitulates the sequential developmental stages of in vivo retinal morphogenesis[64,65]. Firstly, during eye field specification, pluripotent stem cells (PSCs) differentiate into eye field-committed progenitor cells. Subsequently, these progenitors evaginate to form optic vesicles dominated by optic vesicle epithelial cells. Following this, the optic vesicle undergoes invagination to establish a bilayered optic cup structure: the inner layer proliferates to generate the neuroepithelium containing multipotent retinal progenitor cells (RPCs), while the outer layer differentiates into the RPE. During neurogenesis, RPCs progressively differentiate into RGCs, PR precursors, horizontal cells, and amacrine cells. Thereafter, the PR precursors mature into functional PRs, concurrent with the differentiation of bipolar cells and Müller glial cells. Finally, synaptic connections between PRs and RGCs are established, endowing the organoids with light responsiveness and electrophysiological functionality. The stage-specific cellular composition and cytoarchitecture critically determine post-transplantation integration outcomes.
RPCs, though pluripotent and theoretically capable of replacing degenerated cells, exhibit limited integration and differentiation efficiency in practice[66]. Animal studies suggest their therapeutic effects may primarily arise from neuroprotective mechanisms rather than direct cell replacement[67]. However, neuroprotection alone cannot sufficiently restore the visual circuitry in patients with advanced retinal degeneration, where large-scale integration-competent PRs are required. A number of studies have also shown that late graft integration is not satisfactory[68]. Although early grafts and late grafts exhibit a similar integration efficiency with the inner nuclear layer (INL) of the host, they do so in different manners. Early- and late-stage grafts demonstrate comparable integration rates with the host's INL, but through distinct mechanisms: immature grafts retain developmental plasticity to form complete INL/ONL structures integrated with the host's INL, whereas mature grafts fail to undergo structural reorganization[61]. In human RO transplantation studies, Shirai et al.[69] systematically engrafted organoids at different differentiation timepoints into murine hosts, identifying days 50–60 as the optimal window for PRs' survival and synaptic connectivity with the host's bipolar cells. Subsequent studies[62,70] using SFEBq-derived organoids corroborated these findings. Stage-specific analyses via immunostaining and quantitative real-time polymerase chain reaction (qRT-PCR) revealed PAX6+/ChX10+ RPC populations, emergent Brn3+ RGCs, and apically localized Crx+ PR precursors with progressive mRNA upregulation during this critical phase[70]. Research on the isolation and transplantation of PR precursors indicates that fully mature PRs may exhibit greater fragility than PR precursors, rendering them unsuitable for transplantation[71]. Notably, PR precursor cells (PRPCs) isolated from retinal organoids exhibiting progenitor-stage Crx expression demonstrate enhanced synaptic integration competency compared with terminally differentiated counterparts[35]. In conclusion, the current data suggest that retinal organoids in the early differentiation stage have the best PR survival and integration efficiency, and the optimal transplantation stage of ROs varies from culture protocol to culture protocol due to the different different differentiation times required for the different protocols, which can be analyzed and screened for ROs in each period of time by immunostaining, qRT-PCR, and flow cytometry.
Researchers have encountered challenges in precisely determining the optimal time for implantation during the cultivation of human ROs due to their extended growth period. To ensure that the developmental status of the organoids aligns with the physiological characteristics of human retinas, researchers must enhance their cultivation strategies to improve physiological similarity. This involves achieving precise correspondence in both the temporal and spatial dimensions with the human retina's development. Consequently, ROs should be able to produce a large quantity of PR precursor cells in vitro and mature progressively according to normal developmental pathways. This optimized cultivation environment is critical for identifying the appropriate transplantation timing, ensuring that the transplanted ROs can achieve maximal functional recovery.
Reducing immune rejection
-
The immune-privileged status of the retina is mediated by the blood–retinal barrier (BRB), comprising retinal capillary endothelial cells and RPE cells. This dual-layered system restricts the infiltration of circulating immune cells and macromolecules into retinal parenchyma, thereby attenuating immunological rejection − a critical advantage for transplantation[72]. The subretinal space is 'fully immunoprivileged' and the neuroretinal tissue with blood vessels is 'partially immunoprivileged'[73,74]. Surgically preserved BRB integrity enables the survival of hESC–RPC grafts without systemic immunosuppression[29]. Conversely, intraoperative mechanical disruption triggers acute T cell/macrophage infiltration, precipitating early-phase PR graft loss[75]. Large-animal hESC–retina transplantation introduces additional complexity: inevitable vascular trauma during vitrectomy induces immunocyte clustering[76]. Thus, clinical translation necessitates the standardization of procedures by vitreoretinal surgeons with specialized expertise in minimizing iatrogenic RPE/choroidal injury. Occelli et al.[44] established optimized feline-specific delivery protocols, providing a roadmap for scaling retinal organoid transplantation to human applications.
However, the degenerative retina inherently harbors a proinflammatory milieu. Activated microglia, peripherally derived macrophages, RPE-secreted cytokines, and neutrophil extracellular traps collectively impair organoids' integration efficiency[77−80]. T cells dominate the post-transplant alloimmune responses[81−83]. Current immunomodulatory strategies include: (1) host immune ablation, (2) pharmacological regulation (see Table 1), and (3) graft immunogenicity reduction[84]. Although rodent models of immunodeficient visual impairment have been established[70], it is pharmacological immunomodulation that is more applicable, given the clinical translation. In animal experiments, researchers have used a variety of immunosuppressive agents to reduce the inflammatory response, including traditional cyclosporin A and anti-CD28 antibodies, and many transplant protocols require the drugs to be initiated one week prior to surgery in order to facilitate a stable grafting environment for the retinal organoid. Drug evaluation should include not only efficacy and safety, but should also consider the developmental and metabolic effects on ROs. To develop a graft tolerance treatment strategy for mycophenolic acid (MPA) versus tacrolimus (TAC), Sims et al.[83] used retinal organoids for preoperative exposure to serum- relevant concentrations of the drug, and assessed the drug's pharmacodynamics postoperatively to ensure low-risk dosing. Immunotherapy is necessary from the point of view of helping the reconstruction of visual function structures. Considering the side effects of drug dosage, the specific dosing regimen can be flexibly arranged by detecting the degree of inflammation in the patient's retinal microenvironment, e.g., topical administration in patients with low inflammation, and combined systemic medication in patients with high inflammation. In addition, is it possible to consider repairing the damaged BRB in conjunction with organoid transplantation, e.g., via drugs containing heparin-modified hyaluronic acid hydrogels with topical slow-release vascular endothelial growth factor (VEGF) receptor inhibitors (e.g., abciximab, norethindrone[85] and dexamethasone[86]) to stabilize endothelial cells[87].
Table 1. Exploration of dosing regimens to reduce immune rejection in retina-like organ transplantation.
Therapeutic agents Efficacy Ref. Dexamethasone, rapamycin Survival and differentiation of organoids were only successful under dexamethasone treatment. Rapamycin treatment showed no efficacy in preventing immune rejection. [86] Prednisolone, cyclosporine Combination therapy resulted in higher donor cell survival rates and reduced immune responses. [76] Prednisolone, cyclosporin A, and mycophenolate mofetil Combined drugs suppressed rejection, supported long-term donor cell survival, and prevented rapid donor cell loss in untreated hosts post-transplantation. [54] MPA, TAC Immunosuppressants enhanced PRs' survival and functional performance. [83] Autologous iPSC–retinal transplantation does not require one to consider immune rejection of the graft, but generating a single batch of autologous cells requires significant monetary and time costs and is not suitable for widespread clinical use[73]. To reduce the immune rejection associated with allogeneic transplantation, a variety of cyclic guanosine monophosphate (cGMP)-grade high-quality hPSCs can be stocked, allowing the transplanted cell lines to be genotypically matched to most patients[84]. It is also possible to reduce the immunogenicity of hPSC–retina transplants. Although retinal-like organs do not pass on antigens to the host, donors knocking out histocompatibility complex antigens can delay the infiltration of inflammatory cells[88].
Investigation of graft functionality
Reconstruction of visual structure
-
The main objectives of retinal organoid transplantation for the treatment of RDDs are to repair the retinal structure as well as to restore retinal function by replacing damaged PR cells, reconstructing the retinal neural network, and restoring retinal function. Reconstruction of the visual structure is a prerequisite for functional recovery, which consists of two main aspects: (1) reconstruction of the PR layer, i.e., migration of donor PRs to the correct position in the host retina, and (2) reconstruction of the neural circuit, i.e., synaptic connections between donor PRs and the host's bipolar cells[45]. By using technologies such as fundus cameras, optical coherence tomography (OCT), immunohistochemistry (IHC), confocal scanning laser ophthalmoscopy (CSLO), and fluorescence in situ hybridization (FISH), researchers can accurately monitor the viability state of donor cells and their distribution within tissues (see Table 2). These techniques exhibit distinct operational characteristics in imaging mechanisms, resolution thresholds, technical complexity, and clinical translatability. Fluorescence adaptive optic SLO (FAOSLO), with single-cell resolution, allows noninvasive tracking of fluorescently labeled donor cells' survival, migration, and morphological maturation in vivo. For nonfluorescently labeled donor cells and related clinical applications, histological studies at multiple time points are suboptimal. However, the limited resolution and range of observation of a single imaging technique not only prevent a comprehensive assessment of graft integration but have also misidentified mononuclear inflammatory infiltrates as grafts[89]. Integrated multimodal approaches combining OCT, cSLO, and molecular profiling currently represent the gold standard for assessing graft integration and immune rejection[54,90].
Table 2. Common methods and uses for detecting graft status in RO transplantation studies.
Technique Application Can it be used
in the clinic?Fundus camera Enables in vivo real-time monitoring of graft viability, spatial distribution, and host retinal structural changes. Yes OCT Provides high-resolution 3D tomographic retinal imaging for graft tracking, retinal thickness quantification, and detection of immune-mediated complications (e.g., macular edema). Yes CSLO Generates 2D en face retinal images for dynamic morphological assessment of transplant regions, resolving fine structures (e.g., PRs' outer segments). Yes Adaptive optic SLO (AOSLO) Achieves cellular-level resolution for in vivo visualization of RPE/PR morphology, structural alignment, and spatial orientation. Yes FAOSLO Combines AOSLO's resolution with molecular specificity, enabling noninvasive longitudinal tracking of fluorescently labeled donor cells without terminal histology. Yes TEM Validates ultrastructural reconstitution ex vivo, cone inner/outer segment formation, and synaptic ribbon connectivity with the host's bipolar cells. Yes IHC Postmortem analysis of cellular stratification, differentiation status (recoverin+/rhodopsin+), and synaptic connectivity (ribbon synapse quantification). No FISH Post-transplantation mapping of donor cells' distribution and survival kinetics in fixed retinal sections. No TEM, transmission electron microscopy; FISH, fluorescence in situ hybridization. The donor cells' positioning and functional integration depend on coordinated biological mechanisms. As previously discussed, surgical methodology (lamellar vs. suspension grafts), the developmental stage of transplanted cells, the host's retinal microenvironment, and immunomodulatory strategies collectively determine the outcomes of integration. Preretinal transplantation is superior to subretinal transplantation for cell integration in the target area of transplantation[29]. In addition to these influences, the extracellular matrix (ECM)[91], the outer segmental membrane (OLM)[92] and the glial barrier[93] are the main biological barriers known to interfere with this process, and the chances of correct graft positioning can be improved by optimizing the ECM's composition[94], disrupting the OLM's structure[54,95] and inhibiting glial proliferation[96]. Scaffold-based technology for cell delivery can guide graft implantation into specific anatomical sites and may further increase the chances of the graft's success[45,97]. In silico techniques can also be used to screen and identify ligands that promote the spontaneous migration of transplanted cells. By delivering these ligands in vivo, transplanted cells can be directed to migrate to the right niche[98].
Under normal physiological conditions, ROs exhibit a strictly laminar structure in which PR precursor cells have spontaneously formed a stratified layer. When retinal slices are transplanted into the retinal gap, these cells do not need to migrate further to form the PR layer, but their axons still need to migrate to establish the correct neural connections[73]. Neuronal information exchange primarily relies on chemical synapses. In histological terms, synaptic connectivity and the generation of functional structures are fundamental elements for the transplanted cells to achieve functional replacement. After immunosuppression, the establishment of initial axonal connections and initial synaptic connections between hESC–retinal tissues of grafts and receptor cat retinas was robust and rapid[76]. Santos-Ferreira et al.[99] considered the appropriate supplementation of brain-derived neurotrophic factor (BDNF) and chondroitinase ABC or the use of BDNF or Down syndrome cell adhesion molecules to induce dendritic growth and arborization to promote synaptogenesis. Lee et al.[100] developed a biodegradable microstructured ultrathin scaffold. This scaffold supports the correct polarization of hESC-derived retinal PRs within the host's eye. It also enhances the synaptic connectivity between PRs and the host. Currently, researchers use IHC methods to label human donor cells with the corresponding synaptic markers, and there is a lack of techniques that allow direct visualization of the synaptic connections between transplanted PR cells and the host's bipolar cells in vivo[101]. This state of affairs poses new challenges for future development of imaging technology, and there is an urgent need to develop technologies that enable the detection of synaptic-level connectivity to enhance our scientific understanding of functional integration in the post-transplantation retina and to improve clinical validation.
Functional tests
-
Enhancement of the host's visual function post-transplantation requires further validation through behavioral and electrophysiological experiments (see Table 3). To enable functional PRs to operate, they must undergo isomerization and oxidation to recycle visual pigments. This process involves enzymatic steps within the visual cycle, converting all-trans retinol into 11-cis retinol. In RPE cells, all-trans retinol is first converted to 11-cis retinol, which is then further oxidized to 11-cis retinol[102]. If the transplanted tissue does not connect with the host's RPE cells, the production of 11-cis-retinol will be hindered, thus affecting the generation of light responses. In this scenario, regularly supplementing the transplanted animals with 9-cis-retinyl acetate can enhance the PRs' light response, thereby supporting their performance in behavioral tests[74]. In addition, bioadhesives can be used to make co-grafts from ROs and RPE slices to support the normalization of the phototransduction cycle and the healthy survival of PRs[103].
Table 3. Functional tests used in RO transplantation studies.
Name Content Advantage Disadvantage Behavioral tests OKT The ability of an animal to produce a normal optokinetic nystagmus (OKN) in response to a rotating streak or moving light spot stimulation indicates basic visual perception. Reflects spatial visualization abilities without training Behavioral variables interfere, limited resolution, inability to distinguish between cell types SAS The animals were induced to actively avoid the light area by light stimulation and electrical or noise stimulation to determine whether their visual function was restored. Rapid, noninvasive, applicable to a wide range of animal models Dependent on the animal's learning ability, interfered with by other senses, unable to differentiate between cell types VGS Training animals to respond oculomotorly to specific visual stimuli to assess their advanced visual processing ability and the integrity of the retina–optic nerve–brain pathway. Evaluate advanced visual processing, quantifiable, clinically relevant Relies on complex training and high-precision equipment Visual acuity testing By training an animal to actively avoid a certain unfavorable environment when a specific visual stimulus is presented, it indirectly reflects its visual perception and behavioral integration abilities. Rapid, noninvasive, applicable to a wide range of animal models Dependent on the animal's learning ability, interfered with by other senses, unable to differentiate between cell types Electrophysiological tests ERG Recording the overall electrical response of the retina to light stimulation by means of electrodes. Noninvasive, easy to operate, reflective of overall functionality Unable to provide localized or single-cell information, limited sensitivity multifocal ERG (mERG) Similar to ERG, it has the ability to differentiate the functional status of different areas of the retina through multipoint stimulation and mathematical modeling. Noninvasive, easy to follow, reflects overall function Complex equipment, high signal interference, high training requirements SC electrophysiology By implanting electrodes in the region of the SC in an animal model and recording the electrical activity of neurons, it can be determined whether retinal signals could be perceived by the superior colliculus after transplantation. High spatial and temporal resolution, direct reflection of visual signaling Highly invasive, complex operation, difficult signal resolution MEA Retinal tissue was placed on the MEA chip, and its electrical activity in response to light stimulation was recorded. High spatial and temporal resolution, can record the synchronized activity of multiple cells Applicable to ex vivo models only Calcium imaging Indirect reflection of neuronal activity by detecting changes inthe intracellular calcium ion concentration in isolated retinal slices. High spatial resolution, simultaneous recording of multiple neurons, ability to distinguish cell types Signal delay, data processing complexity, only for ex vivo Noninvasive and readily accessible assessments represent advantages in behavioral evaluations. However, certain tests exhibit heightened sensitivity to dysfunctions in the primary visual pathways. These require extensive animal training and repeated trials. The testing conditions of the subjects and the adherence to operational protocols by personnel can also impact the stability of the experimental outcomes. Common visual behavioral experiments include optokinetic tracking (OKT), the light–dark transition test, and the visual water task. The advantage of OKT lies in its requirement of no training for mice. It tests the mice's reflexive response to a vertically rotating grating, wherein their heads move toward the direction of the grating's rotation[104]. Although OKT is one of the most representative behavioral assessments, Mandai et al.[105] found that some mice exhibit immobility during OKT, potentially compromising the accuracy of the results. To address this issue, researchers shifted to the shuttle avoidance behavioral experiment (SAS). In this experiment, mice are trained to actively move from one compartment to another upon detecting light, thereby avoiding an impending shock. This testing method can also be tailored by adjusting the intensity and interval of the shocks to accommodate the varying response characteristics of different mice, thereby enhancing the sensitivity and reliability of the assessment.
The measurements of visually guided saccades (VGS) closely relate to improvements in visual fields and graft positioning. In a study using rhesus monkeys as a model for retinal implant research, researchers conducted long-term visual function monitoring through VGS[59]. The monkeys faced a color LED monitor, focusing on a central fixation point. Subsequently, a target appeared at a random location on the monitor. If the monkeys accurately directed their saccade to the target, it counted as a correct response. In many instances, despite multifocal electroretinography (mERG) failing to yield the expected ideal results, the subjects' behavioral tests indicated positive changes in visual function. In such cases, behavioral assessments provided crucial supplementary information to the ambiguous outcomes of electrophysiological tests.
In electrophysiological assessments, the spatial distribution of rescued PRs is confined to the transplantation area. Consequently, traditional full-field electroretinography (ERG) may fail to accurately identify visual responses triggered exclusively by the transplant[106]. The results of ERG are most relevant when there is extensive cell integration between the donor and the host. Significant differences exist between the electrophysiological outcomes of allogeneic and xenogeneic transplants. Under similar conditions, multi-electrode array (MEA) recordings demonstrate pronounced mERG and RGC responses following mouse iPSC (miPSC) retinal transplants[70]. Conversely, hESC retinal implants in mice show no mERG response and only yield RGC reactions, which exhibit lower amplitude compared with miPSC-derived retinas. This discrepancy likely results from synaptic binding affected by immune rejection. Multiple factors may influence the intensity and response time of the light-induced retinal ganglion cell responses caused by transplanted cells. These factors include the length of outer segment (OS), the efficiency of phototransduction, synaptic transmission, and the physiological characteristics of intermediate neurons in the degenerated retina[53]. Given the uncertainty surrounding the ERG effect, one may consider monitoring light response signals using the potential of the superior colliculus (SC). The SC is a fundamental visual brain region that receives incoming information from the visual cortex. It serves as one of the termination sites for retinal ganglion cell axons. In rats with normal vision, a flash can easily evoke a response across the entire SC region. After transplanting hESC-derived retinal patches into RCS rats, researchers observed SC responses rather than ERG responses[107]. Only a limited area of the SC showed reactions to light stimuli, correlating with the regions of the host's retina that contained the transplant. This finding confirms that SC recordings are more sensitive than full-field ERG measurements. The heat map of the response on the SC shows a shape similar to the position of the graft in the host's retina, which indicates a point-to-point projection of the graft from the retina to the SC[101].
The patch-clamp technique serves as a precise method for assessing synaptic function between grafts and hosts. However, due to its operational complexity, it proves unsuitable for testing large cell populations. Retinal activity must transmit through retinal synapses. Calcium imaging technology plays a critical role in studying the activity and function of ROs. Measuring calcium ion dynamics in the presynaptic area can indicate synaptic transmission activities triggered by synaptic vesicle release. This approach allows for the detection of transplanted neurons responding to neural inputs. He et al.[108] used a two-photon microscope to detect GCaMP5 RPCs' migration to the receptor retina, forming synaptic structures in donor–host retinal grafts. By employing (S)-3,4-dicarboxyphenylglycine (DCPG) to activate the metabolic glutamate receptor mGluR8 at the presynaptic terminals of transplanted PRs, a characteristic reduction in intracellular calcium ions is observed, thereby confirming the functional activity of the transplant[109].
In studies of RO transplantation for the treatment of RDDs, behavioral and electrophysiological tests are key tools for assessing therapeutic efficacy, functional integration, and visual recovery. They verify the recovery of retinal function after transplantation at the levels of animal behavioral responses and cellular/tissue electrical activity, respectively. Behavioral tests reveal the overall functional state of the retina through a macro-level analysis of behavioral responses. In contrast, electrophysiological tests supplement this information by monitoring neural activity at a micro-level. Multimodal assessment methods are an indispensable systematic functional validation strategy in RO transplantation research[110,111]. By combining retinal imaging techniques, behavioral tests, and electrophysiological tests, it is possible to comprehensively assess whether the transplanted cells are alive, electrically active, forming functional connections, and affecting the output of visual behavior, from low-level perception to high-level behavioral responses. In order to more accurately assess the effect of visual function recovery in patients, a combined high-throughput behavioral–electrophysiological system needs to be developed in the future to achieve automated, multimodal functional assessment. Meanwhile, in vivo electrophysiological techniques, such as flexible electrode patches or micro-implanted electrodes[112], should be vigorously promoted to enable long-term recordings of patients in the waking state.
-
The Kobe City Eye Hospital in Japan conducted the first human application of RO sheets derived from allogeneic iPSCs (jRCTa050200027)[113]. This trial involved transplanting RO sheets into patients with advanced RP. Over a two-year period, the grafts did not exhibit adverse events; however, no significant functional improvement was observed. This clinical study on ROs derived from iPSCs has preliminarily demonstrated the safety of the treatment. However, due to the limited number of participants − only two patients − along with the variability in the severity of visual impairment and functional responses, these findings are insufficient for broader application. Writing protocols on the establishment of standardization of RO transplantation therapies is a key component in moving the therapy from laboratory research to clinical application. Standardization not only helps to ensure reproducible and comparable transplantation results but also meets the basic requirements for regulatory approval, ethical review, and clinical translation. Through our review and discussion of transplantation methods, screening of the period of graft differentiation, the immune response during transplantation, and recovery of visual structure and function after transplantation, we believe that a standardized system for RO transplantation therapy should cover the three processes pre-surgery, surgery, and post-surgery.
(1) Pre-surgical planning: the patient's disease status should be assessed, the human leukocyte antigen-matched cell source should be selected, the appropriate transplantation method should be chosen, the number of cells or organoids to be transplanted should be pre-designed, and appropriate immunosuppression should be carried out in advance to create a suitable environment for the survival of the grafts. This process should control the quality of organoids, clarify the cell culture conditions and the number of passages, and identify the quality of the transplanted batches of organoids in terms of size, cell purity, cell type, etc.[74]. We need to improve the method of organoid transportation to ensure that the organoid samples are active from the laboratory to the operating room[101].
(2) Standardized surgery: Strict surgical procedures should be developed to clarify the target area for transplantation, standardize transplantation techniques, and avoid mechanical damage to the retina as much as possible.
(3) Postoperative management: We must develop a pharmacological intervention program, choosing between topical or systemic administration, to help integrate the graft into the patient's retina. Moreover, we need to develop a recovery schedule that combines retinal imaging with behavioral and electrophysiological methods at multiple time points to monitor the patient's visual structure and function over time, with an emphasis on integrating and grading multimodal data.
In RO transplantation studies, ideal visual function restoration should be demonstrated as follows. Behaviorally, the animal can perceive light stimuli, perform visual navigation tasks, and restore color and detail vision; electrophysiologically, the ERG waveform is normal, the ganglion cell firing pattern is restored, and signaling efficiency is good; structurally, the grafted cells are localized in the outer nuclear layer and the outer ganglion is structurally intact and laminar with the host's bipolar cells. In terms of structural integration, the transplanted cells are localized in the outer nuclear layer, the outer segmental structure is intact, and they form a laminar arrangement with the host's bipolar cells. The research into RO transplantation in animal models has yielded promising results; however, significant challenges remain before its clinical application, including the following.
(1) Despite ongoing improvements in differentiation protocols for ROs, the developmental timeline remains lengthy, with suboptimal production efficiency and reproducibility. There is a pressing need for more straightforward, stable, and rapid differentiation methods. Difficulties in isolating 'good quality' ROs on a large scale and standardizing the size and shape of ROs are also limiting factors, but it is expected that advances in biomanufacturing and bioproduct development will soon be able to address these barriers[74].
(2) In the commercialization and clinicalization of retina-like organ transplantation therapy, cost control is a key factor in determining its large-scale dissemination and clinical application. Although this therapy has shown promising potential in animal models, cell culture, transplantation surgery, functional evaluation, and regulatory approval all involve high technical and resource investments[114].
(3) Although RO transplantation therapies have great clinical potential, they also raise many ethical issues, including cell source and informed consent, the ethical challenges of clinical trials, long-term follow-up and potential risks, patient acceptance, and societal impacts[115].
These issues need to be fully emphasized and properly addressed in the course of technological development to ensure that retina-based organ transplantation technology can bring the greatest benefits to patients and society in compliance with the ethical and legal framework.
-
Stem cell-derived RO technology offers unprecedented opportunities for the study of the human retina and the development of treatment strategies for RDDs. Various evidence suggests that human ROs can replicate the natural physiological features of the retina in vitro and ultimately develop the capacity to respond to light stimuli. This remarkable capability underscores the potential of these models. They serve as a foundation for the advancement of transplant therapies. Moreover, they can simulate diseases and facilitate drug development. A simplified and rapid cell differentiation process, strong reproducibility, and biocompatibility are essential. Suitable surgical equipment and techniques must exist. Additionally, the ability to survive and mature in vivo over an extended period is critical. The restoration of connections with the existing retinal pathways is also vital, as is the recovery of lost visual functions. These elements represent the necessary criteria for the transplantation of ROs in treating RDD. It is encouraging that iPSC-derived retinas have achieved new advancements in differentiation and implantation under xeno-free/cGMP conditions[17,116]. This marks a crucial step toward clinical application. Preliminary quality control strategies and short-term preservation methods for RO sheet transplantation are under continuous development to support clinical applications[60]. The process of ROs' preparation and transplantation can be significantly optimized through the application of tissue engineering, microfluidics, and biomaterials[117]. Future studies need to address the issues of cell integration, functional recovery, and long-term stability to facilitate the clinical translation of retinal organoid transplantation technology.
-
Not applicable.
This study was funded by the National Key Research and Development Program of China (No. 2022YFF1202900), Tianjin Municipal Science and Technology Program (No. 22JCZDJC00260, No. 22JCQNJC00980), and the Science Research Fundation of Aier Eye Hospital Group (No. Aiyanxiang-LCERI-002, No. AF2211D04).
-
The authors confirm their contributions to the paper as follows: draft manuscript preparation: Ye S, Hua X; study design: Ye S, Li H; manuscript revision and study supervision: Zhang W, Chen L; preparing the figures and tables: Sun Q, Mi B. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ye S, Zhang W, Sun Q, Mi B, Li H, et al. 2025. Evaluation of the safety and efficacy of retinal organoid transplantation: a comprehensive consideration of morphological, functional, and transplant techniques. Visual Neuroscience 42: e015 doi: 10.48130/vns-0025-0013 

Evaluation of the safety and efficacy of retinal organoid transplantation: a comprehensive consideration of morphological, functional, and transplant techniques
- Received: 20 February 2025
- Revised: 01 May 2025
- Accepted: 29 May 2025
- Published online: 21 August 2025
Abstract: Retinal degenerative diseases refer to a group of disorders characterized by the gradual degradation of retinal tissue structure and function. With their incidence on the rise, there is an urgent need to develop new therapeutic strategies. Retinal organoids, as an emerging technology with promising potential for the treatment of retinal degenerative diseases, play a crucial role in constructing a disease model and drug screening. Here, we review recent developments in retinal organoid research, focusing particularly on their application in transplantation methodologies. We aimed to optimize the implantation process of retinal organoids while qualitatively and quantitatively assessing the degree of visual function recovery. This evaluation intended to determine their potential and effectiveness in treating retinal diseases, thereby advancing the clinical application of organoid transplantation technologies. Although animal experiments have demonstrated that retinal organoids hold significant potential in improving retinal structure and function, there remains a lack of sufficient clinical evidence to support their application efficacy in humans. Therefore, future studies should further optimize implantation protocols for retinal organoids and refine the associated evaluation strategies, with the goal of integrating more clinical data. This will, in turn, provide a more robust evidence base for the application of retinal organoids in human subjects.