








-
Solanum lycopersicum L., commonly known as the tomato, is an herbaceous plant of the Solanaceae family and a modern, widely cultivated vegetable recognized for its red, sweet, and sour edible fruit[1]. Tomato fruits are low in calories and rich in the antioxidant lycopene, along with essential vitamins, and many minerals, providing humans with a rich source of nutrients that are of great nutritional value[2]. Due to its relatively simple growth and development process, short cycle, and fully sequenced genome, tomato is often used as a model system for studying the regulatory mechanisms of fruit development, ripening, and quality formation[3]. Tomato fruit is classified as climacteric, similar to apple, banana, peach, and kiwifruit. Climacteric fruits exhibit a marked increase in respiration rate and ethylene production during the ripening process[4−6]. These characteristics of climacteric fruits are frequently utilized in agricultural practices to regulate the post-ripening process and optimize storage conditions of fruits for various crops[7,8]. Ethylene is a fundamental phytohormone and it was identified as a key factor in the regulation of the ripening process of climacteric fruits by promoting color change, softening, and flavor development, thereby enhancing both edibility and economic value[6,9]. In addition to its role in climacteric fruit ripening, ethylene also plays a pivotal role in various plant growth and developmental processes[9]. ERFs are known to act at the downstream of the ethylene signal pathway and mediate the diverse functions of ethylene by directly regulating the expression of ethylene-responsive genes.
ERFs belong to the AP2/ERF (APETALA2/Ethylene Response Factor) family which is a crucial gene family in plants, playing an important role in the response to biotic and abiotic stresses, as well as in various aspects of plant growth and development. AP2/ERF transcription factors are characterized by the presence of one or two highly conserved AP2/ERF domains, which enable them to bind to specific DNA sequences. This binding influences the expression of downstream target genes, leading to various responses in the plant. AP2/ERF domains typically comprise about 60 to 70 highly conserved amino acid residues, which are crucial for binding to DNA cis-acting elements. Based on the number and similarity of AP2 domains, the AP2/ERF family can be divided into three subfamilies: AP2, ERF, and RAV (Related to ABI3/VP). The AP2 subfamily contains two duplicated AP2/ERF domains, whereas the ERF subfamily contains only a single AP2/ERF domain[10]. The ERF subfamily is further categorized into ERF and dehydration-responsive element binding (DREB) subgroups based on conserved amino acid residues within the AP2/ERF domain[11,12]. The differences in these amino acid residues lead to variations in promoter affinity and recognition specificity between ERF and DREB[13]. The RAV subfamily has a highly conserved AP2 domain, similar to the ERF subfamily, but also contains an additional B3 domain, contributing to its unique regulatory functions[14,15]. In different species, more detailed classifications of the ERF family have been established[11−17]. For instance, in rice and Arabidopsis thaliana, a distinct soloist subfamily has been identified[14]. This subfamily comprises ERF transcription factors with significant structural differences from other subfamilies while maintaining highly conserved nucleotide sequences across different species. Nevertheless, the functions and molecular mechanisms of the soloist subfamily remain largely unclear at present[18]. The ERF family is involved in the regulation of various physiological processes such as seed germination, leaf growth, root development, flowering, fruit development, and ripening[18−22]. Moreover, the ERF family also plays important roles in plant responses to both biotic and abiotic stresses, encompassing drought, high salinity, low temperature, pathogens, and other stress-related stimuli[18,23−26]. Several ERF genes have been identified as potential regulators of primary or secondary metabolic processes[27]. Members from the ERF family typically regulate physiological processes in plants by binding to specific cis-acting elements located within the promoter of genes associated with stress responses (DRE/C-repeat (CRT) cis-elements), or developmental pathways (GCC-box)[10]. Protein interaction analyses suggests that AP2/ERF proteins can interact with other regulatory proteins to coordinately regulate plant growth and development[28]. Overall, the ERF family plays an important role in plant development and stress response, and further elucidation of their functions and mechanisms is crucial for systematically understanding their contributions to plant physiology.
In tomato, 77 ERF family genes have been identified and divided into nine subfamilies (ERF.A-ERF.J)[29,30]. So far, various tomato ERF family genes have been reported to play important roles in various developmental processes and responses to biotic and abiotic stress (Fig. 1). In this review, we summarized the role and mode of action of the ERF family in tomato and discussed the potential use of these genes by molecular breeding in crop improvement.
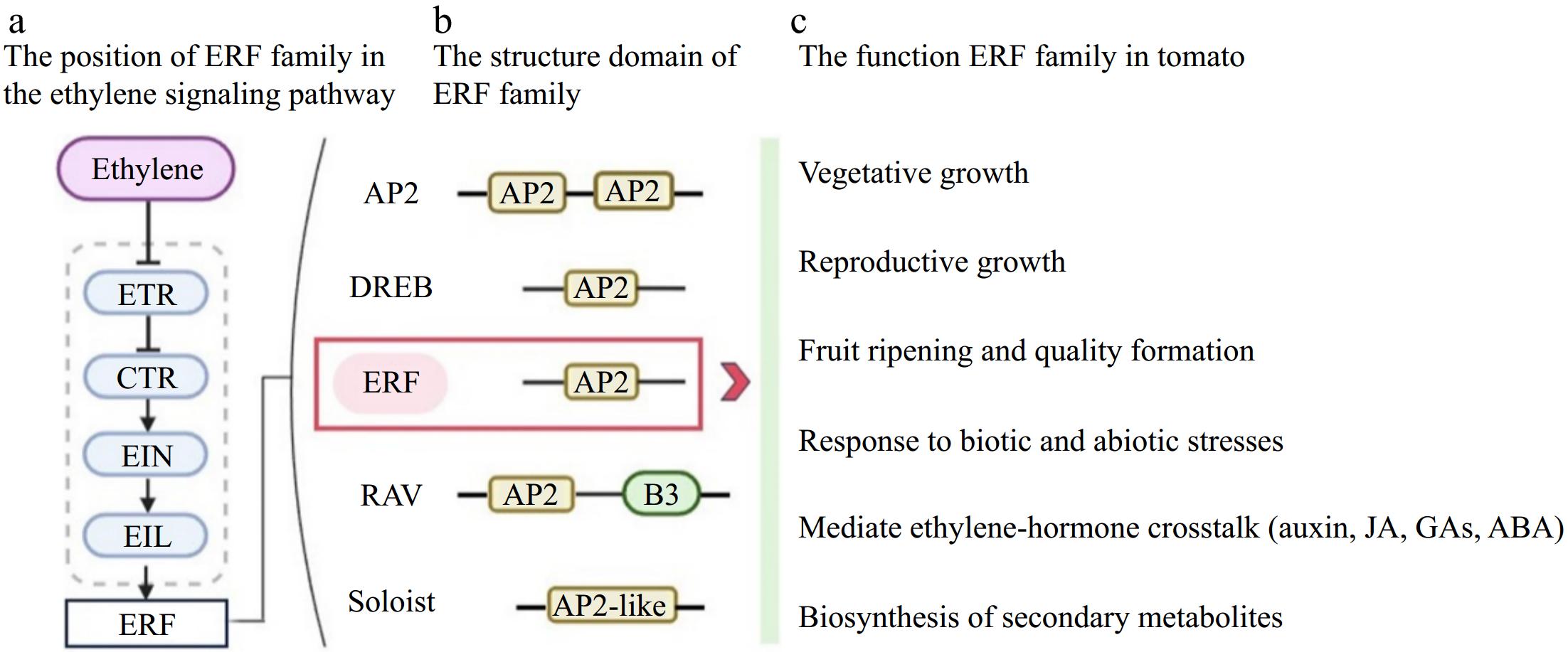
Figure 1.
The location of ERF in ethylene signal pathway and the roles of ERFs in various developmental process and response to biotic and abiotic stress. (a) Hierarchical positioning of ERF transcription factors within the ethylene signaling cascade. Simplified schematic illustrates the sequential activation of core pathway components (abbreviations defined below) culminating in ERF-mediated transcriptional regulation. (b) Phylogenetic classification and domain architecture of the AP2/ERF superfamily. Conserved AP2/DNA-binding domains are highlighted across subfamilies (DREB, ERF, RAV, Soloist), with structural variations denoted by color-coded motifs. (c) Multifunctional roles of ERFs in plant development and stress adaptation. Network analysis reveals ERF involvement in: Developmental transitions (vegetative/reproductive growth); Fruit ripening and quality trait modulation; Cross-talk with hormonal pathways (jasmonic acid, auxin, gibberellins, abscisic acid); Biotic/abiotic stress response coordination; Secondary metabolite biosynthesis. Abbreviations: AP2/ERF – APETALA2/Ethylene Response Factor; DREB – Dehydration-Responsive Element Binding; RAV – Related to ABI3/VP1; ETR – Ethylene Receptor; CTR – Constitutive Triple Response; EIN – Ethylene Insensitive 3; EIL – EIN3-Like; ERF – Ethylene Receptor Factor; JA – Jasmonic Acid; GA – Gibberellin; ABA – Abscisic Acid.
-
The functions of ERF family members during the vegetative growth stage of tomato mainly include the regulation of hypocotyl elongation, root development, and overall plant growth. For instance, ERF transcription factor SlERF.J2 has been reported to act as a negative regulator of hypocotyl elongation and plant stature in tomato[31]. SlERF.J2 exerts its regulatory role by binding to the promoter region of Indole Acetic Acid protein IAA23, thereby suppressing its expression and subsequently influencing plant stature and hypocotyl elongation in tomato plants[31]. Similarly, the ERF.B subfamily gene SlERF.B3, has been reported to regulate root development and plant growth by integrating ethylene and auxin signaling pathways via modulating the expression of an auxin response factor SlIAA27[19]. Further study showed that SlERF.B3 promotes the expression of SlIAA27 by directly binding to specific sequences within the promoter region of SlIAA27, thereby initiating its expression and affecting plant sensitivity to auxin[19]. In addition, SlERF36 was identified as a regulator controlling the growth cycle of tomato plants[32]. Subsequent investigations revealed a correlation between these physiological changes and alterations in gibberellic acid (GA) levels[32]. More recently, it was reported that overexpression of SlERF.H5 or SlERF.H7 resulted in strong growth inhibition phenotypes[33]. It has been shown that SlERF.H5 and SlERF.H7 inhibit the expression of GA20ox1, a key gene involved in gibberellin synthesis, thereby affecting plant growth[33]. These findings illustrate the key role of ERFs in mediating plant development through interactions with other hormones such as auxin and gibberellins.
During the reproductive developmental process in tomato, ERF family members have been shown to play an important role in regulating flowering time and flower abscission Specifically, SlERF52 functions as a positive regulator of flower abscission in tomato[34]. SlERF52 is specifically expressed in the pedicel abscission zone (AZ), where it up-regulates the expression of genes associated with flower abscission and cell wall hydrolysis, such as cell-wall hydrolytic enzymes Cel and PG (Cel5, TAPG1, TAPG2, and TAPG4). This regulatory activity promotes the progression of flower abscission in tomato plants[34]. The EAR motif containing transcription factor SlERF36 was also reported to modulate the onset of flowering in tomato[32,35]. In addition, it was shown that overexpression of SlERF5 in tomato resulted in earlier flowering, an increase in the number of petals and the diameter of the flowers[36]. However, these alterations in reproductive growth did not influence changes in fruit ripening quality[36]. Despite these findings, there is limited research focusing on the regulation of ERF transcription factors during the reproductive growth stage, and detailed molecular mechanisms are not yet well elucidated. Addressing this knowledge gap represents a significant direction for future research on ERF transcription factors and their role in reproductive development.
-
ERF family members have been shown to play important roles in regulating fruit ripening and quality in tomato. Fruit ripening signifies a pivotal transition in the life cycle of plants, particularly in species with fleshy fruits such as tomatoes, garnering considerable attention and study from researchers[21,28]. Although the ripening process of fruits exhibits distinct characteristics and unique pathways across different plant species, common features include changes in fruit color, texture, aroma, sugar content, acidity, and other biochemical substances. During the ripening process, many ERF genes exhibit differential expression patterns, suggesting their involvement in the ripening process[31]. Among these, LeERF1 has been reported to positively mediate the ethylene response and regulate the fruit ripening process in tomato[28]. The expression of the PG gene, which is highly correlated with fruit softening was up-regulated in LeERF1 overexpression (LeERF1-sn) lines[28]. Conversely, SlERF.F12 was reported to act as a repressor in fruit ripening by interacting with the corepressor TPL2, facilitating the recruitment of HDA1 and HDA3 to form a co-repressor protein complex[21]. This complex binds to the promoters of ripening-related genes, such as 1-aminocyclopropane-1-carboxylate (ACC) synthase (ACS) ACS2, ACS4, PG2a, and Pectate Lyase (PL), thereby inhibiting their expression and delaying the ripening process[21]. In a study conducted in 2022, it was demonstrated that SlERF.D6 plays a significant role in tomato fruit ripening and steroidal glycoalkaloid metabolism[20]. More recently, SlERF.D6 was reported to act as a key ripening regulator by the formation of a transcriptional cascade with SlDEAR2 and SlTCP12[37]. Additionally, SlERF.E4 has been identified to bind to the promoter region of the β-D-N-acetylhexosaminidase (β-Hex) gene, where it acts as a positive transcriptional regulator. Molecular mechanisms revealed that SlERF.E4 promotes the expression of β-Hex during tomato fruit ripening, thereby facilitating fruit softening through conjunction with Ripening Inhibitor (RIN) and Abscisic Acid Stress Ripening (SlASR1)[22]. Using dominant repression technology (SRDX), S1ERF.B3 was shown to exert a significant effect on tomato fruit ripening[38]. Inhibition of SlERF.B3 activity by the SRDX fusion protein led to the downregulation of Phytoene Synthase PSY1 and Phytoene Desaturase PDS expression, while upregulated Lycopene Cyclase b-LCY1 and genes associated with ethylene production and ripening[38]. Similarly, the downregulation of SlERF6 expression resulted in elevated ethylene production, accompanied by increased expression of ACS2 and ACC oxidase (ACO) ACO1, suggesting that SlERF6 acts as a negative regulator in fruit ripening[39]. Furthermore, this study demonstrated that SlERF6 controls carotenoid biosynthesis by negatively regulating the expression of Heat Shock Protein 21 HSP21 and 1-Deoxy-D-xylulose-5-phosphate synthase DXS[39]. Three ethylene response factors Pti4, Pti5, and Pti6 also have been shown to function in fruit ripening[40]. Overexpression of these genes resulted in a shorter fruit ripening time by up-regulation of ripening-related genes such as ACS2 and ACS4[40]. In contrast, the expression of LeERF3b was downregulated during fruit ripening but significantly increased in the Never-ripe (Nr) mutant[24]. These findings suggest that LeERF3b may act as a negative regulator in fruit ripening[24]. A recent study revealed that SlERF.H5 and SlERF.H7 genes play vital roles in tomato fruit softening[33]. These ERFs can bind to the DRE cis-acting element in the promoter region of the Cellulose Synthase gene SlCESA3, thereby increasing the expression of SlCESA3 and leading to enhanced synthesis of cellulose in the fruits[33]. In addition, SlERF.G3-Like has recently been proven to be related to fruit ripening in tomato, with its expression directly regulated by the well-known fruit ripening regulator RIN[41]. Moreover, SlERF.G3-Like exerts a regulatory effect on several downstream genes involved in tomato fruit ripening and quality, including SlACS2, SlACO1, SlPL, PG2a, Chalcone synthase CHS1, Flavonol synthase FLS, and Flavonoid 3'-hydroxylase F3H[41]. Collectively, the functions of the ERF family in fruit ripening have been extensively investigated compared to their roles in vegetative and reproductive growth. Studies have demonstrated that the regulatory pathways governed by ERF family genes in fruit ripening are diverse, contributing to complex regulatory networks. The effects of ERF family genes on fruit ripening encompass various traits such as fruit color, texture, flavor substances, and secondary metabolites. However, fully comprehending the specific pathways through which ERF family genes regulate these tomato traits, as well as their intricate relationship with ethylene signaling remains a challenging endeavor. Future research endeavors are anticipated to make significant breakthroughs in elucidating these pathways addressing these fundamental questions and contributing to advancements in tomato breeding and fruit quality improvement.
-
The ERF family, known for its responsiveness to ethylene signaling, is also recognized for its crucial role in mediating plant responses to both biotic and abiotic stresses[42]. ERF family members are involved in the regulation of disease and insect resistance in tomato when confronted under biotic stress conditions[43]. Studies have shown that the expression of some ERF transcription factors is induced by pathogens and insect infestations, thereby initiating defense responses in tomato[44]. It was shown that the expression of ERF2 gene in tomato plants was upregulated in response to Stemphylium lycopersici infection[45]. Silencing ERF2 led to a susceptible phenotype, reduced hypersensitivity response, and alterations in physiological activities such as reactive oxygen species (ROS) production[45]. Furthermore, ERF2 confers resistance to the invasion by S. lycopersici by playing a key role in multiple SA, JA, and ROS signaling pathways[45]. Several members of the ERF family, particularly the B3 subfamily, including SlERF.A1, SlERF.B4, SlERF.C3, and SlERF.A3, play a critical role in conferring resistance to Botrytis cinerea in tomato[46]. Silencing these genes has been shown to increase susceptibility to pathogens and diminish pathogen-induced jasmonic acid/ethylene-mediated signaling responses[46]. This downregulation consequently affected the expression of defense genes involved in the immune responses of plants[46]. In another study, overexpression of ERF68 induced spontaneous cell death and activated multiple defense pathways, including salicylic acid (SA), ethylene (ET), jasmonic acid (JA), and hypersensitive response in tomato[44]. In addition, Pti4, Pti5, and Pti6 genes, previously identified as key players in plant defense responses, share a conserved DNA binding domain characteristic of Ethylene-responsive element binding proteins (EREBPs)[47,48]. This domain facilitates interactions with Pto kinases, thereby regulating the expression of downstream pathogenesis-related (PR) genes[48]. In the absence of this binding domain, Pti4, Pti5, and Pti6 are unable to interact with Pto kinases, rendering tomato plants more susceptible to disease[47,48]. The expression of Tomato Stress Responsive Factor TSRF1 was up-regulated in response to ethylene, salicylic acid, or tomato Ralstonia solanacearum infection[49]. Overexpression of TSRF1 constitutively activates the expression of PR genes in tomato, thereby enhancing tomato resistance to Ralstonia solanacearum infection[49]. Furthermore, recent studies have highlighted the significant role of the SlERF.C1 in Botrytis cinerea response[50]. SlERF.C1 interacts with the mitogen‐activated protein kinase SlMPK8, thereby enhancing the regulation of SlERF.C1 to the downstream PR genes by phosphorylating SlERF.C1[50].
In response to abiotic stresses such as drought and salt stress, ERF family members are involved in regulating adaptative responses and stress tolerance mechanisms in tomato[51]. It has been shown that the expression of various ERF transcription factors can be induced by abiotic stresses such as high salinity, drought, and low temperature[51]. ERF transcription factors play crucial roles in modulating growth and developmental processes in tomato, ultimately enhancing the plant´s ability to tolerate and adapt to adverse environmental conditions[52]. Notably, SlERF5 exhibited significant upregulation in response to various abiotic stresses such as high salt, drought, flooding, and low temperature, suggesting that SlERF5 plays a pivotal role in the response to abiotic stresses in tomato[52]. Overexpression of the SlERF5 resulted in increased resistance to abiotic stresses such as drought and salt stress in transgenic plants[52]. Similarly, the ethylene response factor ERF.C1 also named TERF1 or JERF2 gene, which responds to ethylene signaling, exhibits increased tolerance to salt stress when overexpressed during seed germination and vegetative growth stages[25]. These findings suggest that TERF1 acts as a positive regulator in the plant's response to salt stress[25]. Overexpression of JERF3 in transgenic plants enhances plant tolerance to salt stress, demonstrating JERF3's role as a key regulator in mediating salt stress responses[53]. In addition, the JERF1 gene demonstrates versatility in binding not only to the GCC-box motif but also to the DRE-box[54]. This gene responds not only to ethylene signaling but also to stressors such as salt stress, abscisic acid, and methyl jasmonate[54]. Through these interactions, JERF1 activates the expression of PR genes containing the GCC-box, thereby enhancing tomato plant tolerance to salt stress[54]. These findings provide further evidence supporting the pivotal role of ERF family genes in diverse stress response.
-
Functioning as the principal ethylene-responsive transcription factors, ERFs likely play a pivotal role as signaling hubs that facilitate the integration of ethylene pathways with other phytohormone signaling networks. Interactions between ethylene and auxin have long been reported at both the physiological and molecular levels, and in recent years more evidence supports the cross-talk between the two hormones[26,55]. Ethylene-responsive gene SlERF.B3 has been shown to integrate ethylene and auxin signaling via regulation of the expression of the auxin signaling component SlIAA27, suggesting that SlERF.B3 may act as a key mediator linking ethylene and auxin signaling in tomato[19]. Similarly, SlERF.D7 amalgamates auxin and ethylene signaling pathways via regulating the transcript accumulation of SlARF2 paralogs in tomato, thereby orchestrating tomato fruit ripening processes[56]. It was shown that SlERF.J2 suppresses IAA23 expression and modulates auxin and GA homeostasis, thereby regulating hypocotyl elongation and plant height in tomato[31]. Furthermore, in addition to auxin, ethylene signaling also interacts with other plant hormone pathways, such as those regulated by Jasmonic acid (JA), abscisic acid (ABA), and gibberellins (GAs). Jasmonate-inducible ERF transcription factors have been shown to act as key regulators that integrate JA signaling to activate steroidal glycoalkaloid biosynthesis in tomato, potentially mediating crosstalk between ethylene and JA pathways in plant defense responses[57]. Ethylene-induced ERF15 and ERF16 act as master transcriptional activators of JA biosynthetic genes, orchestrating the rapid jasmonate burst in tomato during herbivory, with ERF16 further amplified by a JA-MYC2 feedback loop[58]. Moreover, SlERF.B8 plays a positive role in tomato cold tolerance by triggering jasmonate biosynthesis and acts as a crucial node integrating ethylene and JA signaling pathways[59]. Additionally, recent studies have demonstrated that ERFs can mediate the crosstalk between ethylene and other plant hormones such as GAs and ABA to affect plant growth and development[60−62]. SlERF.H6 mediates the interplay between ethylene and gibberellin signaling to regulate the SGA biosynthesis in tomato[63]. SlERF36 accelerates growth transitions and reduces the plant life cycle by regulating GA levels and responses[32]. Ethylene response factor Pti4 is involved in the regulation of fruit ripening, seed germination, and responses to drought and Botrytis cinerea infection through adjustments to ABA metabolism and signaling[64]. Tomato ERF84 can be markedly induced by ABA, and its overexpressing plants confer hypersensitivity to ABA at the germination stage[65]. Similarly, ERF15 positively regulates ABA-mediated cold tolerance in tomato plants by activating CBF1 and WRKY6 expression[66]. These studies supported the important role of ERFs in mediating the crosstalk between different plant hormones. Despite the growing number of studies showing the phytohormone crosstalk mediated by ERFs, the underlying mechanisms still require further investigation.
-
As a key transcription factor family, ERFs also play a vital role in regulating secondary metabolite biosynthesis in tomato. In addition to the well-known functions in controlling the biosynthesis of carotenoids[67], ERFs have been shown to act as key regulators in the modulation of the biosynthesis of flavonoids in tomato. For example, ethylene response factor SlERF.G3-Like was reported to positively promote the accumulation of flavonoids in tomato fruits by directly regulating the expression of key structure genes involved in flavonoid biosynthesis[41]. Except for the important roles in the biosynthesis of carotenoids and flavonoids, ERFs are involved in steroidal glycoalkaloids (SGAs) metabolism. GAME9, an ERF family member, was reported to modulate SGA metabolism by regulating the expression of GAME17[68]. More recently, a distal enhancer GAME Enhancer 1 (GE1) was shown to recruit GAME9-MYC2 complex to form a chromatin loop, thereby regulating the metabolism of steroidal glycoalkaloids (SGAs) in tomato[69]. In addition, metabolome-wide association analysis identified SlERF.H6 as a critical regulator in controlling SGA metabolism in tomato. SlERF.H6 binds to the GCC-box present in the promoter regions of multiple GAME genes, negatively regulating their expression, and thus SGA biosynthesis[63]. Similarly, SlERF.D6 has been identified as a dual regulator, influencing both fruit ripening and SGA metabolism. SlERF.D6 suppresses the transcription of GAME12, thereby negatively regulating tomatidine synthesis[20]. ERFs are also shown to be involved in the regulation of phenolic biosynthesis in tomato. Ethylene response factor SlERF7 was reported to promote UV-C-induced phenolic biosynthesis in postharvest tomato fruit by binding to the promoter of SlPAL5[70]. Moreover, two ERF genes ERF1 and ERF4 were found to be differentially expressed in high-phenolic tomato introgression line IL7-3[71]. Further studies demonstrated that ERF1 acts as a key regulator of phenolic transport into the vacuole, while ERF4 acts as an auxiliary transcription factor in modulating phenolic metabolism[71]. These findings collectively highlight the multifaceted roles of ERF transcription factors in orchestrating secondary metabolite biosynthesis in tomato, with implications for fruit development, coloration, and nutritional quality.
-
The ERF family of transcription factors has emerged as a pivotal player in tomato breeding, offering immense potential for improving agronomic traits, fruit quality, and stress resilience. Their multifaceted roles in regulating vegetative growth, reproductive development, fruit ripening, and stress responses underscore their significance as candidates for crop enhancement (Table 1). These studies offer significant insights into the regulatory mechanisms that govern tomato growth and development offering valuable references for improving both tomato yield and quality. However, despite these successes, several limitations persist. The functional redundancy and pleiotropic effects of ERF genes often complicate their precise manipulation, as altering one ERF member may inadvertently affect multiple physiological processes. Additionally, the intricate crosstalk between ERFs and other hormonal pathways, such as auxin, gibberellins, and jasmonates, remains poorly understood, limiting our ability to predict outcomes of genetic modifications. Current research also lacks comprehensive field studies to validate the performance of ERF-engineered tomatoes under real-world conditions, leaving gaps between laboratory findings and practical applications. Looking ahead, future research should prioritize unraveling the complex regulatory networks governed by ERFs, particularly their interactions with other signaling pathways, to enable more precise and predictable genetic interventions. Advances in CRISPR-based genome editing and systems biology approaches could facilitate the targeted modulation of ERF genes while minimizing unintended effects. Moreover, integrating ERF manipulation with traditional breeding techniques may accelerate the development of novel tomato cultivars with optimized traits. Another promising direction is the exploration of ERF homologs across diverse tomato wild relatives, which could uncover untapped genetic variation for stress adaptation and fruit quality, thereby further developing new tomato varieties with optimized plant structure, high quality, and enhanced resistance.
Table 1. Summary of the role of ethylene response factors (ERFs) in tomato.
Gene identifiers Names New names Function Ref. Solyc02g090790 ERF.J2 SlERF.J2 Plant growth [31] Solyc05g052030 ERF.B3 SlERF.B3 Root development [19] Solyc10g006130 SlERF36 SlERF.F1 Plant growth [32] Solyc03g117130 SlERF52 Flower abscission [34] Solyc10g006130 SlERF36 SlERF.F1 Flowering time [35] Solyc03g093560 SlERF5 SlERF.B2 Flowering time [36] Solyc05g050830 SlERF.H5 SlERF.H5 Plant growth [33] Solyc06g066540 SlERF.H7 SlERF.H7 Plant growth [33] Solyc06g065820 LeERF1 SlERF.H1 Fruit ripening [28] Solyc02g077840 SLERF.F12 Fruit ripening [21] Solyc04g071770 ERF.D6 SlERF.D6 Fruit ripening [20] Solyc01g065980 ERF.E4 SlERF.E4 Fruit ripening [22] Solyc05g052030 ERF.B3 SlERF.B3 fruit ripening and
ethylene response[38] Solyc10g065980 ERF6 SlERF.E4 Fruit ripening and
carotenoid synthesis[39] Solyc05g052050 Pit4 SlERF.A3 Fruit ripening [40] Solyc02g077370 Pit5 SlERF.C6 Fruit ripening [40] Solyc06g082590 Pit6 SlERF.G2 Fruit ripening [40] Solyc10g009110 LeERF3b SlERF.F5 Fruit ripening [24] Solyc05g050830 SlERF.H5 SlERF.H5 Fruit firmness [33] Solyc06g066540 SlERF.H7 SlERF.H7 Fruit firmness [33] Solyc02g077790 SlERF.G3-like Fruit ripening [41] Solyc02g077370 ERF2 SlERF.C6 Disease resistance [45] Solyc08g078180 SlERF.A1 SlERF.A1 Disease resistance [46] Solyc03g093540 SlERF.B4 SlERF.B4 Disease resistance [46] Solyc09g066360 SlERF.C3 SlERF.C3 Disease resistance [46] Solyc05g052050 SlERF.A3 SlERF.A3 Disease resistance [46] Solyc08g078180 ERF68 SlERF.A1 Disease resistance [44] Solyc05g052050 Pit4 SlERF.A3 Disease resistance [47,48] Solyc02g077370 Pit5 SlERF.C6 Disease resistance [47,48] Solyc06g082590 Pit6 SlERF.G2 Disease response [47,48] Solyc09g089930 TSRF1 SlERF.C4 Disease resistance [49] Solyc05g051200 SlERF.C1 SlERF.C1 Disease resistance [50] Solyc03g093560 SlERF5 SlERF.B2 Drought and salt tolerance [52] Solyc05g051200 TERF1/JERF2 SlERF.C1 Salt tolerance [25] Solyc03g123500 JERF3 SlERF.E3 Salt tolerance [53] Solyc06g063070 JERF1 SlERF.E2 Salt tolerance [54] Solyc05g052030 ERF.B3 SlERF.B3 Ethylene-Auxin crosstalk [38] Solyc03g118190 SIERF.D7 SIERF.D7 Ethylene-Auxin crosstalk [56] Solyc02g090790 SlERF.J2 SlERF.J2 Ethylene-Auxin crosstalk [31] Solyc06g054630 ERF15 Ethylene-ABA crosstalk [66] Solyc01g090320 SlERF.B8 SlERF.B8 Ethylene-ABA crosstalk [59] Solyc03g120840 SlERF.H6 SlERF.H16 Ethylene-ABA crosstalk [63] Solyc10g006130 SlERF36 SlERF.F1 Ethylene-ABA crosstalk [32] Solyc05g052050 Pit4 SlERF.A3 Ethylene-ABA crosstalk [64] Solyc04g071770 SlERF84 Sl-ERF.D6 Ethylene-ABA crosstalk [65] Solyc02g077790 SlERF.G3-like Flavonoid synthesis [41] Solyc01g090340 GAME9 SlERF.B9 SGA synthesis [68] Solyc03g120840 SlERF.H6 SlERF.H6 SGA synthesis [63] Solyc04g071770 SlERF.D6 SlERF.D6 SGA synthesis [20] Solyc03g123500 SlERF7 SlERF.E3 Phenolic compounds synthesis [70] Solyc05g051200 SlERF1 SlERF.C1 Phenolic compounds synthesis [71] Solyc07g053740 SlERF4 SlERF.F4 Phenolic compounds synthesis [71] The gene identifiers can be found at https://solgenomics.net/ , the Name of ERF genes are derived from the cited literature, the new names refer to the nomenclature in the study by Chen et al.[31]. This work was supported in part by the Institutional Research Funding of Sichuan University (2022SCUNL105), and the Applied Basic Research Category of Science and Technology Program of Sichuan Province (2021YFQ0071 and 2022YFSY0059-1).
-
The authors confirm contribution to the paper as follows: study conception and design: Liu M; draft manuscript preparation: Yu S, Wang H, Garcia-Caparros P, Liu M. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu S, Wang H, Garcia-Caparros P, Liu M. 2025. Revisiting the functions of ethylene response factors (ERFs) in tomato. Plant Hormones 1: e008 doi: 10.48130/ph-0025-0008 

Revisiting the functions of ethylene response factors (ERFs) in tomato
- Received: 06 February 2025
- Revised: 01 April 2025
- Accepted: 08 April 2025
- Published online: 28 April 2025
Abstract: Ethylene is a pivotal phytohormone that regulates a wide array of biological processes in plants, including growth, development, fruit ripening, and responses to environmental stresses. Ethylene-response factors (ERFs), members of the (APETALA2/ERF) superfamily, act as key downstream regulators in the ethylene signaling pathway, through modulating the expression of ethylene-responsive genes. Extensive studies have demonstrated that ERFs are involved in multiple aspects of plant development such as seed germination, plant growth, flowering, fruit development, and ripening, as well as responses to both biotic and abiotic stresses. This review systematically examines the roles and molecular mechanisms of ERFs in various developmental processes in tomato (Solanum lycopersicum L.). Furthermore, we discuss the potential future research directions and the possible applications by manipulation of ERFs based on their important roles in plant growth, fruit ripening, and stress response.
-
Key words:
- Ethylene /
- Ethylene response factors /
- Ethylene signaling pathway /
- Plant growth /
- Fruit ripening /
- Stress response