









-
Heat shock proteins (HSPs) are molecular chaperones important for responding to various biotic and abiotic stressors, ubiquitously present from higher organisms to bacteria[1]. Primarily, HSPs assist in proper protein folding, assembly, and refolding, maintaining protein conformation stability, and participating in numerous biological processes such as macromolecular assembly, polypeptide degradation, and transcription regulation[2−4]. They enhance cellular stress resistance and are closely associated with temperature adaptability[5]. Under stressful conditions like extreme temperatures and drought, HSPs modulate the organism's stress response, minimize cellular damage, promote repair, and facilitate rapid adaptation to the external environment[6].
Since the discovery of HSPs in 1962[7], the involvement of these proteins in processes including stress resistance, apoptosis, and inflammatory responses in living organisms has been the subject of extensive research[8−10]. The structure and function of HSPs are highly conserved, and the family of HSPs can be subdivided on the basis of molecular weight and structure, into the following subfamilies: HSP100, HSP90, HSP70, HSP60, HSP40, and the small HSPs (sHSP)[11,12]. Some studies have already focused on HSPs in mollusks[13−15]. In Octopus vulgaris, HSP90 is associated with somatic cell growth and shows higher transcription levels with increased growth rates[16]. HSP90 plays a pivotal role in the transduction of multiple hormone and growth factor receptors and is essential for cell viability under certain growth conditions[17,18]. Chen et al. found that HSP70 interfered with apoptosis caused by high expression of cytochrome c under salt stress in Crassostrea gigas[19]. Previous studies have shown that HSP70 is involved in endoplasmic reticulum stress-induced apoptosis and affects the expression of apoptosis regulatory proteins[20]. In addition, another study has evidenced a notable elevation in HSP40 expression levels in Venerupis philippinarum in response to vibrio challenge or heavy metal stress[21]. This finding substantiates the pivotal role of HSP40 in the immune response of this species. In the study of the HSP gene family in Pomacea canaliculata, each family formed monophyly on the phylogenetic tree, and more genes showed stronger transcription level under heat stress than under cold stress[13]. Cold stress may induce the expression of HSP70, which is helpful in maintaining the normal physiological function of cells at low temperatures[22]. In one study, the changes of gene expression of Ericerus pela in China under low temperatures were analyzed by WGCNA, among which HSPs were identified as hub genes, and HSPs may play an important role in cold adaptability[23]. These studies show that organisms can effectively cope with temperature stress and enhance their environment viability through the regulation of HSP gene expression[24].
The Chinese mysterysnail (Cipangopaludina chinensis) belongs to the phylum Mollusca, class Gastropoda, and the family Viviparidae. This species is one of the few commercially significant edible freshwater snails in the Chinese market, possessing substantial economic value. In recent years, an increasing number of farmers have elected to engage in C. chinensis breeding in captivity, to satisfy the substantial demand for these snails in the Chinese market. C. chinensis exhibits a wide adaptation, occurring in a range of aquatic habitats, including paddy fields, lakes, and rivers[25]. This species of snail allows its young to develop within the maternal body before oviposition[26]. This ovoviviparous reproduction plays a role in its reproductive strategy in harsh environments[27]. In addition, C. chinensis demonstrates remarkable resilience to a range of environmental stressors, including drought, heat, cold, and water pollution[28]. This adaptability contributes to its extensive distribution in freshwater ecosystems across China.
The identification of gene families facilitates an in-depth comprehension of the biological characteristics and evolutionary history of species[26,29]. At present, there is a paucity of research on gene families of viviparids. To enhance comprehension of the molecular mechanisms underpinning the extensive adaptation of viviparids, including C. chinensis, we have conducted a comprehensive genome-wide identification and characterization of the HSP gene family of C. chinensis. This study provides supplementary data to the existing body of research on HSPs in this species, offering a foundation for future investigations into functional gene studies, molecular breeding, and genetic engineering in this species.
-
The HSP gene family of C. chinensis (CchHSPs) was identified using genomic data from our laboratory that had been uploaded to the China National Center for Bioinformation (Project No. PRJCA032441).
According to the published HSP gene family of the P. canaliculata[13] as reference sequences, the protein sequence was compared with those of C. chinensis using Blastp software[30], with the parameter set to E-value ≤ 1e−9 to identify candidate protein sequences. The candidate HSP gene family was screened and confirmed using HMMER software[31] in combination with the specific domains of the HSP gene family (HSP90: PF00183; HSP70: PF00012; HSP60: PF00118; HSP40: PF00226; HSP20: PF00011; HSP10: PF00166).
The chromosomal location of CchHSPs was extracted from the genome annotation file of C. chinensis using TBtools v2.119 software[32]. The tandemly arrayed gene file obtained from intraspecific collinearity analysis was also verified using TBtools software.
Characterization prediction of CchHSPs
-
The candidate protein sequences were analyzed for conserved domains using the online SMART tool (
http://smart.embl-heidelberg.de ). Subsequently, the protein sequences of the identified HSP gene family were evaluated using the ProtParam tool on the ExPASy website (web.expasy.org/protparam ), which calculated their amino acid length (aa), theoretical isoelectric point (pI), and molecular weight (MW). The subcellular localization of these proteins was predicted using the CELLO online server (http://cello.life.nctu.edu.tw ).The intron, exon, and non-coding region structures were visualized using the GSDS2.0 online tool (
http://gsds.gao-lab.org ). The conserved motifs of the CchHSPs were predicted and analyzed using the MEME suite (http://meme-suite.org/tools/meme ). The maximum limit number of detected motifs was set to 20, with functional domain lengths ranging from 30 to 100 amino acid residues. The minimum number of occurrences for the functional domain was set to 10, while other parameters remained at default settings[33]. The domain structure of CchHSPs was predicted using NCBI's BatchCD-Search tool (www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ), and all domain sequences were extracted based on the predicted positions. The secondary structure of CchHSPs was predicted using the GOR4 online tool (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html ), and the 3D protein structures were modeled using the SWISS-MODEL interactive website (https://swissmodel.expasy.org/interactive ).To analyze the protein interaction network of CchHSPs, the protein sequences were uploaded to the STRING online database (
https://cn.string-db.org/ ), with P. canaliculata serving as the model. The maximum number of interactions displayed was set to 10 for the first layer, and 20 for the second layer.Phylogenetic analysis of the HSP gene family in gastropods
-
To further investigate the relationship of HSP gene families among C. chinensis and its relatives, we also used the uploaded genomic data in the National Center for Biotechnology Information (NCBI) database to identify the HSP gene families of Pomacea maculata (GCA_004794325.1), Biomphalaria glabrata (GCA_000457365.2), Achatina fulica (PRJNA511624), Lautoconus ventricosus (GCA_018398815.1), and Patella pellucida (GCA_917208275.1). In addition, the B. aeruginosa genome obtained from our laboratory was similarly identified for the HSP gene family. Afterward, the protein sequences of the HSP gene family of these seven snails were compared using the default parameters of the MEGA11 software[34]. A phylogenetic tree was constructed using the maximum likelihood (ML) method, with the Jones-Taylor-Thornton (JTT) model employed for ML algorithm analysis. Additionally, a separate ML phylogenetic tree was constructed specifically for C. chinensis. The resulting phylogenetic trees were enhanced using the tvBOT website[35].
Expression levels of CchHSPs in various tissues
-
Total RNA was individually extracted from the antenna, labial palp, foot muscle, testis, ovarian endometrium, kidney, gill, and mantle tissues of C. chinensis using an RNA extraction kit (TaKaRa Bio, Dalian, China). After verifying RNA integrity through agarose gel electrophoresis, total RNA concentration was measured with a micro-UV spectrophotometer (MaestroGen, Taiwan, China). Samples with an OD260/OD280 ratio between 1.8 and 2.0 were selected and sent for sequencing (Fraser Biotech, Wuhan, China). Transcriptome sequencing was performed using a HiSeq4000 sequencer (Illumina, CA, USA) with a read length of 150 PE. The transcriptome data were filtered, and quality controlled using Trimmomatic v0.38 software[36], RNA-seq data are aligned with the reference genome of C. chinensis from this paper using the default parameters of Hisat2 v2.1.0 software[37]. The data were then aligned with the reference genome, and gene expression levels for each tissue were calculated using featurecounts v2.0.1[38]. The expression levels of each member of the CchHSPs gene family in different tissues were integrated into a matrix. Data normalization and a tissue expression heat map of the CchHSPs gene family were generated using the pheatmap package in R Studio.
Expression levels of CchHSPs under temperature stress
-
Ninety healthy, uniformly sized female C. chinensis (26.42 ± 4.75 g) were selected from Yancheng, China, and placed uniformly and randomly into nine 10 L buckets in the laboratory for three days of acclimatization. Afterward, the water temperature of three buckets was set at 0 °C to subject the snails to cold stress, three buckets were set at 30 °C to subject the snails to heat stress, and the other three buckets were kept at room temperature (24 °C) to serve as a control group. After three days, three snails were randomly sampled from each bucket. Kidney, foot muscle, and gill tissues obtained from each snail were used for subsequent total RNA extraction.
Total RNA was also extracted using an RNA extraction kit (TaKaRa Bio, Dalian, China). The cDNA was synthesized using HiScriptIIqRT SuperMixII (Vazyme Biotech, Nanjing, China), and the primers for Real-Time Polymerase Chain Reaction (qPCR) of the target genes were designed using Primer Premier 5.0 software[39]. The primers used are shown in Supplementary Table S1. ChamQ SYBR Color qPCR Master Mix (Vazyme Biotech, Nanjing, China) and QuantStudio3 (Thermo Fisher Scientific, USA) were used for qPCR experiments. The program was conducted as follows: 2 min at 95 °C, 40 cycles (each cycle was for 15 s at 95 °C, 20 s at 56 °C, and 30 s at 72 °C), 30 s at 95 °C, 1 min at 60 °C, and 10 s at 95 °C. Statistical analysis of qPCR results was performed using GraphPad Prism 10 software. Significance levels between groups were analysed using one-way ANOVA and Tukey's post hoc test.
-
Through whole-genome scanning, a total of 36 CchHSPs were identified in the genome of C. chinensis, including six subfamilies: CchHSP10 (1), CchHSP20 (8), CchHSP40 (11), CchHSP60 (1), CchHSP70 (12), and CchHSP90 (3) (Supplementary Tables S2−S3). In C. chinensis, chromosome localization analysis revealed that the 36 CchHSPs were distributed across nine chromosomes. Specifically, CchHSP70s were found on Chr1, Chr2, Chr3, Chr5, Chr6, Chr7, and Chr8; CchHSP10 on Chr1; CchHSP20s appeared on both Chr4 and Chr9, CchHSP40s were present on Chr1, Chr2, Chr3, Chr4, Chr6, Chr7, and Chr8, CchHSP60 was identified on Chr1; while CchHSP90s resided on Chr3 and Chr6. Moreover, we identified four clusters formed by tandemly repeated genes: CchHSP20-01 to CchHSP20-05 on Chr4, CchHSP70-01 and CchHSP70-02 on Chr1, CchHSP70-05 and CchHSP70-06 on Chr2, and CchHSP70-10 and CchHSP70-11 on Chr7 (Fig. 1). The structure of genes within each cluster exhibited high similarity, and the intergenic distances between genes in each cluster were less than 10 kb (Supplementary Table S4).
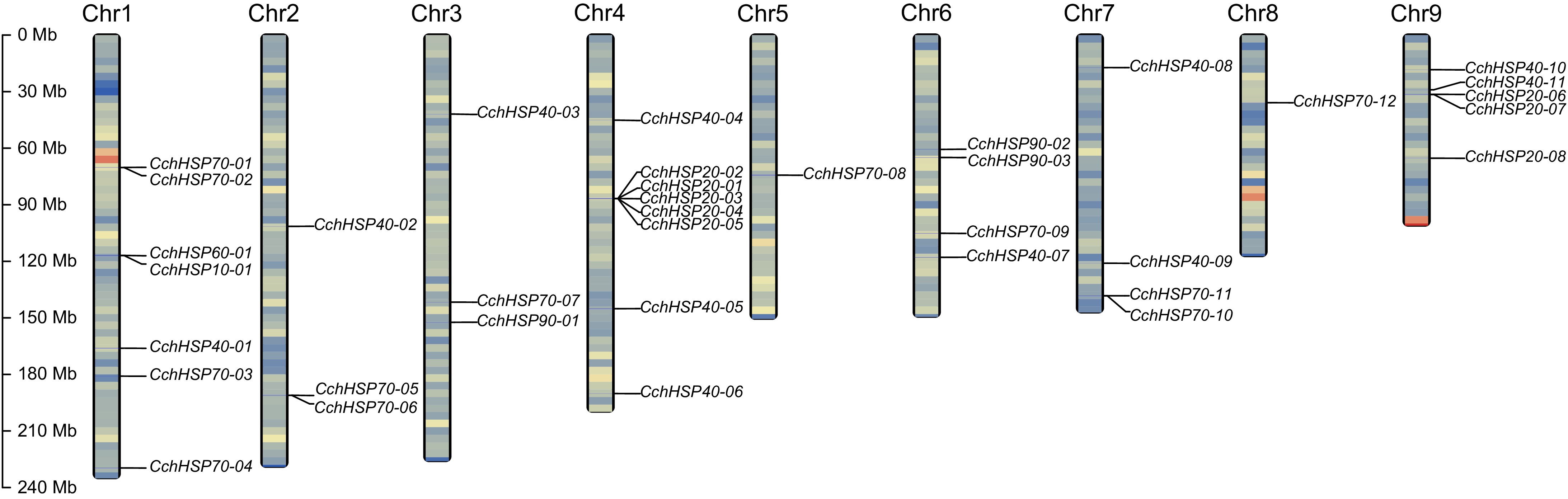
Figure 1.
The chromosomal locations of the identified CchHSPs. The chromosome gene density is expressed by the gradient color of blue-red from low to high.
Physicochemical properties of CchHSPs
-
The CchHSPs vary from 102 to 1,102 aa with the CchHSP70s being the longest and the CchHSP10s the shortest. Generally, an increase in the classification within the HSP gene family correlates with an increase in protein sequence length. The molecular weights (MWs) of CchHSPs range from 11.23 to 124.69 kDa. It is evident from the data that both the protein sequence lengths and molecular weights of CchHSP10s and CchHSP20s are relatively small, while those of CchHSP70s and CchHSP90s are significantly larger. The theoretical isoelectric points (pI) of CchHSPs range from 4.82 to 9.21. Among these, 29 members exhibit a pI between 4.82 and 6.99 (acidic), while seven members have a pI ranging from 7.61 to 9.21 (alkaline) (Table 1).
Table 1. Characteristics summary of HSP genes in C. chinensis.
Gene name Size
(aa)MW
(Da)pI Instability
indexAliphatic
indexGrand average of
hydropathicity (GRAVY)Chromosome
no.Subcellular
localizationCchHSP10-01 102 11,232.16 7.95 35.20 95.53 −0.110 Chr1 Cytoplasm CchHSP20-01 165 18,552.21 5.83 32.45 80.36 −0.445 Chr4 Cytoplasm CchHSP20-02 260 29,741.03 7.61 44.95 76.85 −0.596 Chr4 Cytoplasm CchHSP20-03 168 19,473.30 5.83 41.55 72.38 −0.667 Chr4 Cytoplasm CchHSP20-04 171 19,507.20 5.61 44.42 71.11 −0.629 Chr4 Cytoplasm CchHSP20-05 167 19,123.81 5.75 43.94 74.67 −0.656 Chr4 Cytoplasm CchHSP20-06 432 47,398.99 5.94 58.01 62.75 −0.694 Chr9 Nucleus CchHSP20-07 199 23,147.35 5.37 33.67 64.17 −0.658 Chr9 Cytoplasm CchHSP20-08 646 73,972.96 6.63 57.22 73.19 −0.784 Chr9 Nucleus CchHSP40-01 235 25,939.71 6.99 56.68 46.89 −0.697 Chr1 Nucleus CchHSP40-02 318 35,945.69 8.57 54.98 79.53 −0.823 Chr2 Nucleus CchHSP40-03 354 38,868.21 8.93 33.87 64.72 −0.631 Chr3 Cytoplasm CchHSP40-04 317 35,413.31 6.72 36.37 79.62 −0.494 Chr4 Cytoplasm CchHSP40-05 412 45,624.63 5.89 35.02 66.00 −0.662 Chr4 Nucleus CchHSP40-06 404 44,884.34 6.40 41.65 74.83 −0.591 Chr4 Nucleus CchHSP40-07 493 53,964.07 8.95 44.23 70.43 −0.484 Chr6 Mitochondrion CchHSP40-08 368 42,963.14 8.74 62.88 64.16 −0.986 Chr7 Nucleus CchHSP40-09 356 40,248.48 5.47 41.35 72.19 −0.619 Chr7 Cytoplasmic CchHSP40-10 339 40,677.82 9.21 52.62 80.24 −0.696 Chr9 Nucleus CchHSP40-11 220 24,065.09 5.46 46.26 62.59 −0.540 Chr9 Extracellular space CchHSP60-01 573 61,096.40 5.69 28.52 98.05 −0.093 Chr1 Mitochondrion CchHSP70-01 296 32,945.25 4.82 21.95 84.63 −0.626 Chr1 Endoplasmic reticulum CchHSP70-02 659 72,885.36 5.02 29.26 86.07 −0.494 Chr1 Endoplasmic reticulum CchHSP70-03 431 47,342.62 5.97 30.02 105.36 0.060 Chr1 Cytoplasm CchHSP70-04 657 71,936.88 6.63 36.74 82.68 −0.461 Chr1 Mitochondrion CchHSP70-05 640 70,385.24 5.70 36.86 78.81 −0.499 Chr2 Cytoplasm CchHSP70-06 640 70,440.41 5.86 37.46 78.81 −0.499 Chr2 Cytoplasm CchHSP70-07 518 55,978.35 6.18 38.34 89.34 −0.141 Chr3 Cytoplasm CchHSP70-08 1,102 124,693.00 5.20 37.68 72.29 −0.854 Chr5 Nucleus CchHSP70-09 855 95,997.61 5.24 44.11 70.95 −0.635 Chr6 Cytoplasm CchHSP70-10 637 70,158.17 5.77 33.55 79.62 −0.496 Chr7 Cytoplasm CchHSP70-11 637 70,158.21 5.77 33.39 79.92 −0.491 Chr7 Cytoplasm CchHSP70-12 649 71,456.75 5.55 37.75 79.52 −0.479 Chr8 Cytoplasm CchHSP90-01 792 90,870.62 4.91 36.57 81.96 −0.630 Chr3 Endoplasmic reticulum CchHSP90-02 730 83,874.10 4.93 42.35 79.07 −0.686 Chr6 Cytoplasm CchHSP90-03 730 83,274.87 6.40 42.29 88.59 −0.409 Chr6 Cytoplasm MW, molecular weight; pI, isoelectric point. The instability index of CchHSPs varies from 28.52 to 62.88, while the aliphatic index ranges from 46.89 to 105.36. GRAVY analysis indicates that all members, except CchHSP70-03, are hydrophilic proteins characterized by a GRAVY value of less than zero. Predictions regarding subcellular localization reveal that 20 CchHSPs were predominantly located in the cytoplasm, nine in the nucleus, three in the mitochondrion, three in the endoplasmic reticulum, and one in the extracellular space. Specifically, CchHSP10s and CchHSP20s are primarily localized within the cytoplasm; CchHSP40s are mainly found in the nucleus; CchHSP60s reside within mitochondria; whereas both CchHSP70s and CchHSP90s predominantly occur in the cytoplasm. Consequently, it can be inferred that CchHSPs primarily execute their functions within these specific subcellular compartments.
Gene structure and protein characterization of CchHSPs
-
The results of gene structure prediction are shown in Fig. 2a. To investigate the diversity of protein structures of CchHSPs, we predicted their conserved motifs, which resulted in a total of 20 different motifs being identified (Supplementary Fig. S1). Among these, CchHSP70s exhibited the highest number of conserved motifs, while CchHSP10s displayed the fewest (Fig. 2b).
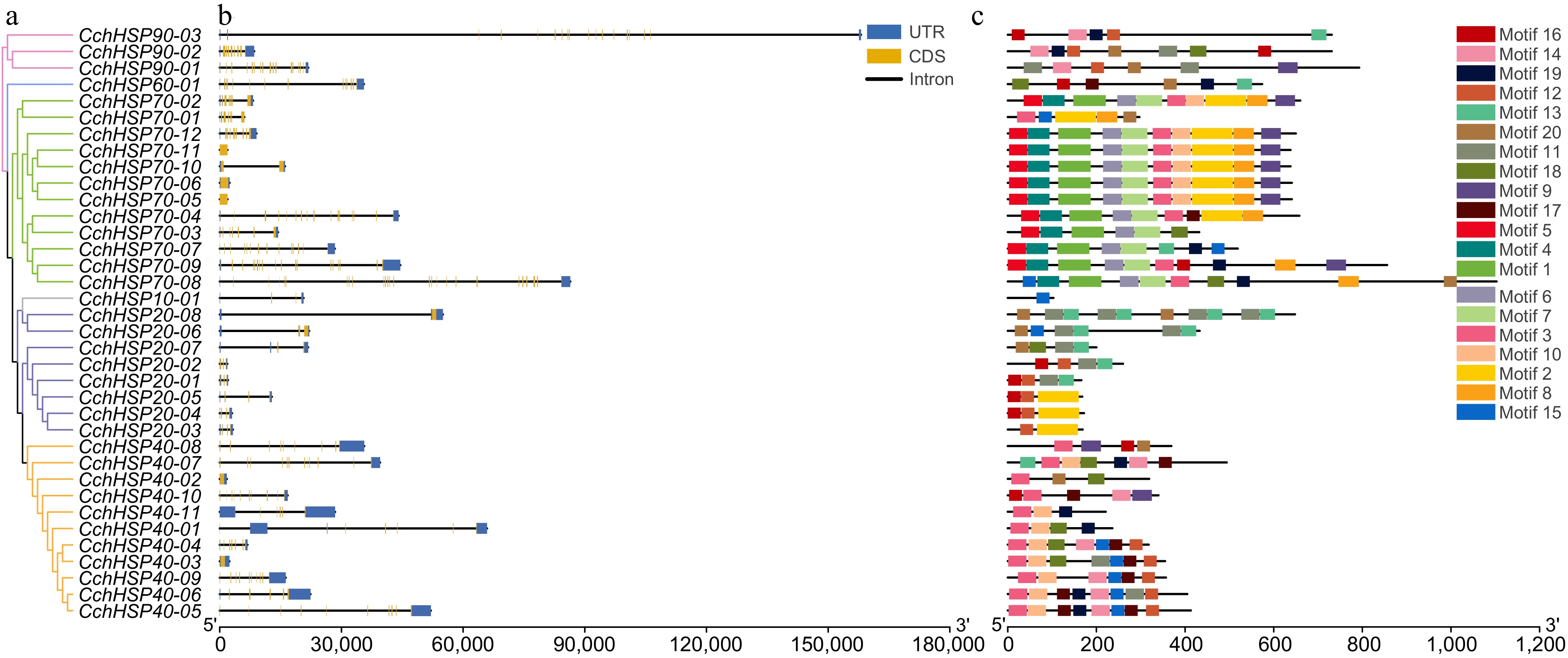
Figure 2.
(a) Phylogenetic relationships of the CchHSPs. (b) Gene structures of the CchHSPs. (c) Conserved motifs of the CchHSPs.
The prediction results for the domain of CchHSPs are illustrated in Fig. 3. Two proteins encoded by CchHSP70 possess domains related to the nucleotide-binding domain (NBD). The HSP40 gene family includes characteristic sequences encode for the DnaJ domain, as well as members of the DnaJ superfamily, RRM_DNAJC17, DUF1977, CbpA, PRPK10767 superfamily, and PTZ00037 superfamily domains. Specifically, CchHSP40-02, CchHSP40-03, CchHSP40-04, CchHSP40-07, CchHSP40-08, CchHSP40-09, and CchHSP40-10 all contain the DnaJ domain, which is more commonly referred to as the J domain. The proteins encoded by CchHSP60 were characterized by a distinct GroEL domain; CchHSP10 had a cpn10 structural domain; CchHSP20 had one or two metazoan_ACD structural domains.
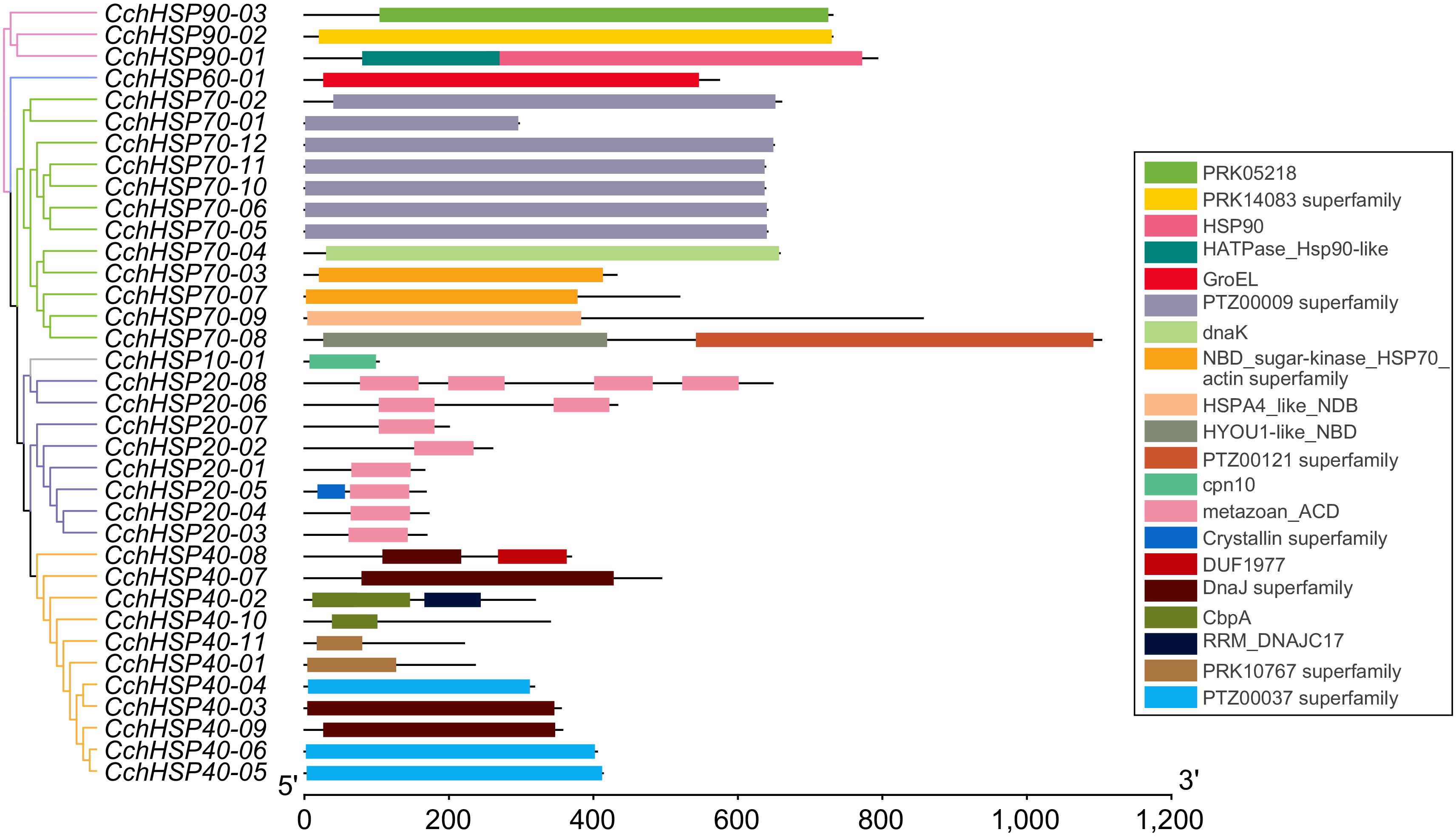
Figure 3.
The protein domain analysis of the CchHSPs.
The secondary structure of CchHSPs primarily comprises alpha helices, extended strands, and random coils. Among these, random coil amino acids represent the highest proportion (35.10% to 64.81%), followed by alpha helix amino acids (9.94% to 52.20%), and extended strand amino acids, which account for the lowest proportion (10.87% to 27.45%) (Table 2). Three-dimensional structure predictions indicate that the genes in sister branches of the same subfamily exhibit similar protein structures (Supplementary Fig. S2).
Table 2. Summary of two-dimensional structures of CchHSPs proteins.
Gene name Sequence length Alpha helix (Hh) Extended strand (Ee) Random coil (Cc) CchHSP10-01 102 28/27.45% 28/27.45% 46/45.10% CchHSP20-01 165 32/19.39% 31/18.79% 102/61.82% CchHSP20-02 260 45/17.31% 58/22.31% 157/60.38% CchHSP20-03 168 19/11.31% 41/24.40% 108/64.29% CchHSP20-04 171 17/9.94% 45/26.32% 109/63.74% CchHSP20-05 167 39/23.35% 32/19.16% 96/57.49% CchHSP20-06 432 92/21.30% 60/13.89% 280/64.81% CchHSP20-07 199 42/21.11% 32/16.08% 125/62.81% CchHSP20-08 646 204/31.58% 131/20.28% 311/48.14% CchHSP40-01 235 71/30.21% 41/17.45% 123/52.34% CchHSP40-02 318 166/52.20% 36/11.32% 116/36.48% CchHSP40-03 354 85/24.01% 73/20.62% 196/55.37% CchHSP40-04 317 70/22.08% 72/22.71% 175/55.21% CchHSP40-05 412 75/18.20% 90/21.84% 247/59.95% CchHSP40-06 404 108/26.73% 77/19.06% 219/54.21% CchHSP40-07 493 114/23.12% 96/19.47% 283/57.40% CchHSP40-08 368 137/37.23% 40/10.87% 191/51.90% CchHSP40-09 356 111/31.18% 44/12.36% 201/56.46% CchHSP40-10 339 148/43.66% 72/21.24% 119/35.10% CchHSP40-11 220 46/20.91% 37/16.82% 137/62.27% CchHSP60-01 573 298/52.01% 70/12.22% 205/35.78% CchHSP70-01 296 100/33.78% 58/19.59% 138/46.62% CchHSP70-02 659 237/35.96% 128/19.42% 294/44.61% CchHSP70-03 431 190/44.08% 73/16.94% 168/38.98% CchHSP70-04 657 294/44.75% 101/15.37% 262/39.88% CchHSP70-05 640 254/39.69% 118/18.44% 268/41.88% CchHSP70-06 640 254/39.69% 117/18.28% 269/42.03% CchHSP70-07 518 210/40.54% 84/16.22% 224/43.24% CchHSP70-08 1102 534/48.46% 133/12.07% 435/39.47% CchHSP70-09 855 346/40.47% 105/12.28% 404/47.25% CchHSP70-10 637 257/40.35% 114/17.90% 266/41.76% CchHSP70-11 637 255/40.03% 114/17.90% 268/42.07% CchHSP70-12 649 272/41.91% 104/16.02% 273/42.06% CchHSP90-01 792 395/49.87% 112/14.14% 285/35.98% CchHSP90-02 730 363/49.73% 91/12.47% 276/37.81% CchHSP90-03 611 279/45.66% 113/18.49% 219/35.84% Phylogenetic analysis of HSPs at both inter- and intra-specific levels
-
The phylogenetic tree of HSPs in C. chinensis was constructed using the maximum likelihood (ML) method (Fig. 4a). The tree is divided into three major branches, with each subfamily gene clustering together separately. Among them, CchHSP90s, CchHSP60, and CchHSP70 are closely related, whereas the CchHSP10s, CchHSP20, and CchHSP40 families, being relatively large, occupy different branches.
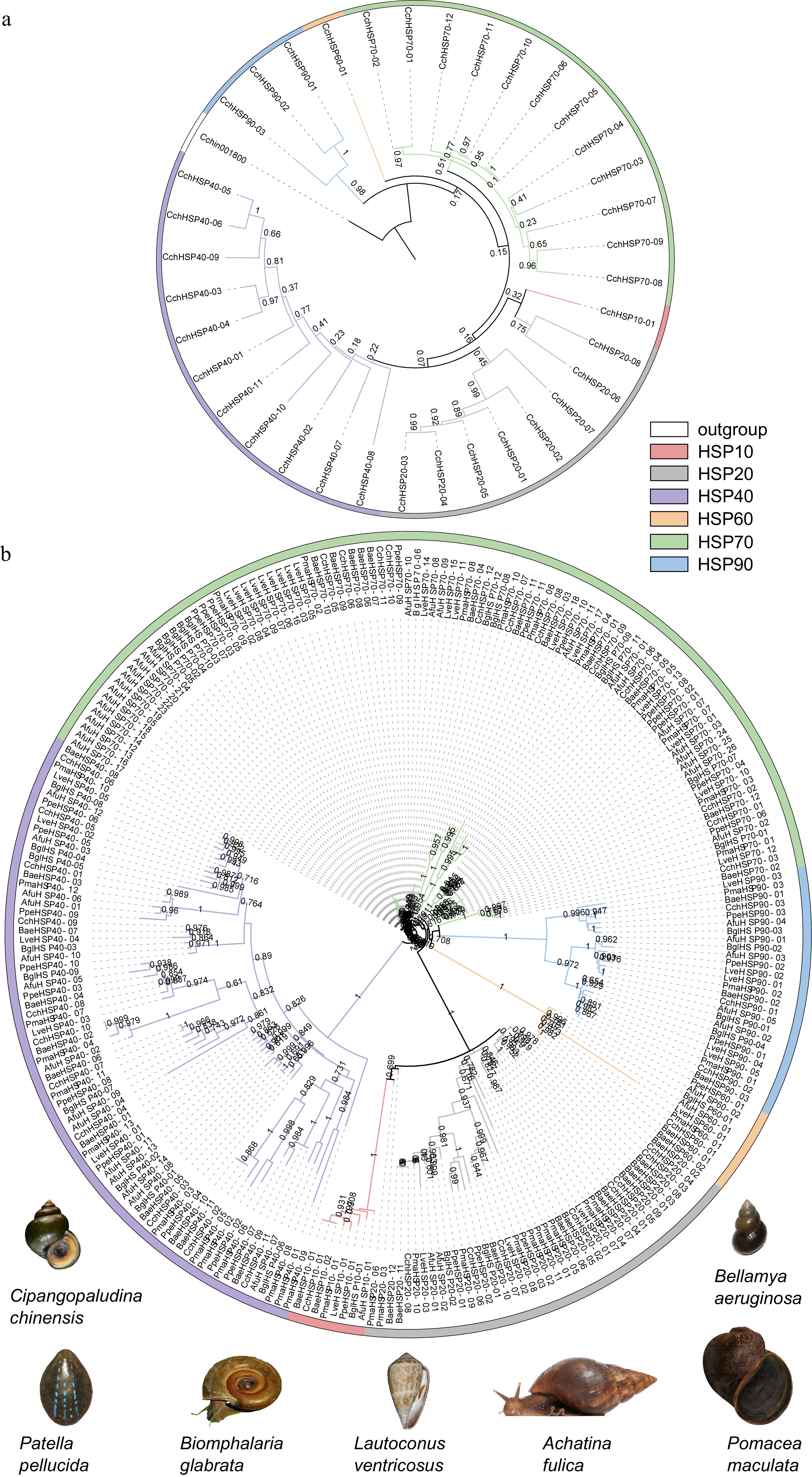
Figure 4.
(a) The phylogenetic tree of the CchHSPs via the maximum likelihood (ML) approach. (b) The phylogenetic analysis for the HSPs in C. chinensis, B. aeruginosa, P. maculate, B. glabrata, A. fulica, L. ventricosus, and P. pellucida.
A phylogenetic tree was also constructed utilizing identified HSP protein sequences from C. chinensis, B. aeruginosa, P. maculata, B. glabrata, A. fulica, L. ventricosus, and P. pellucida (Fig. 4b). This tree is categorized into four major branches, with genes from each subfamily clustering within their respective branches. Furthermore, gene duplication events were observed within the HSP70 subfamily (specifically between CchHSP70-05 and CchHSP70-06 as well as between CchHSP70-10 and CchHSP70-11). The clustering of HSP subfamily genes across species underscores the high reliability of this phylogenetic analysis.
Protein-protein interaction network of CchHSPs
-
Using the STRING, an interaction network analysis of C. chinensis proteins was conducted with P. canaliculata as the reference species to identify potential protein interactions (Supplementary Tables S5−S6). The analysis revealed that 26 CchHSPs were involved in protein interactions (Supplementary Fig. S3). Among these, CchHSP70-9, CchHSP70-12, and CchHSP90-2 had the most interactions, making them key nodes. These three nodes all interacted with NEDD8, A0A2T7PA24, and A0A2T7NZM9 proteins. There was a strong interaction between CchHSP40 genes and CchHSP70 genes. Additionally, the key node CchHSP90-02 had a strong interaction with the Elongin-C and NEDD8 proteins.
According to STRING function annotations, the NEDD8 protein is primarily involved in ubiquitin binding, ubiquitin-like protein ligases, biological processes of cellular macromolecules, various protein metabolism pathways, and the regulation of metabolic pathways. The A0A2T7PA24 protein can bind to HSP90, FK506, and unfolded proteins, participate in the synthesis, degradation, and metabolism of prostaglandins, and regulate the activity of prostaglandin-E synthase. The A0A2T7NZM9 protein can respond to various stimuli such as cytokines and chemicals, like A0A2T7PA24, and has the function of binding to HSPs. It participates in chaperone-mediated protein folding and response to heat and regulates the activity of prostaglandin-E synthase.
Expression profiles of the HSP gene family in different tissues
-
The expression levels of CchHSPs in different tissues were assessed using RNA-seq data and the results are illustrated in Fig. 5. CchHSP10-01, CchHSP20-02, CchHSP20-04, CchHSP40-03, CchHSP70-12, and CchHSP90-02 exhibited high expression (TPM > 90) in all tissues. The TPM values were further analyzed by one-way ANOVA using R software and Tamhane's T2 method was selected for post hoc comparison. The results showed that CchHSP10-01 expression had the highest expression in the kidney, which was significantly higher than that in the lip and gill (p < 0.05). CchHSP20-07 expression was significantly lower in hepatopancreas, ovary, endometrium, kidney, gill, and mantle than in testis (p < 0.05). In contrast, the expression of CchHSP40-08 in testis was the lowest and significantly lower than in foot muscle, hepatopancreas, and ovary (p < 0.05). CchHSP90-01, CchHSP90-02, and CchHSP90-03 were all highest in testis and all were significantly higher than foot muscle, hepatopancreas, and endometrium (p < 0.05).
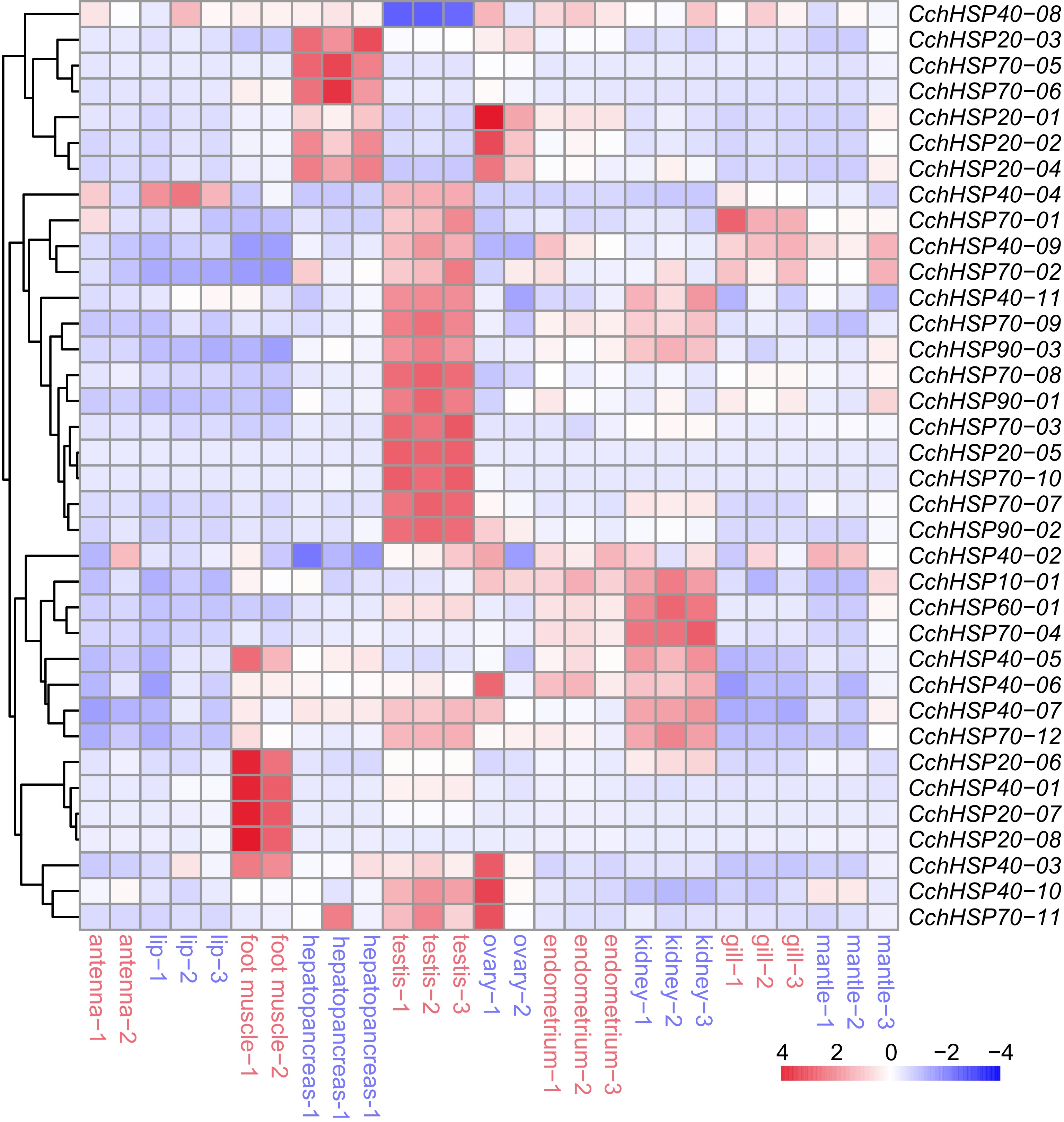
Figure 5.
The expression levels of CchHSPs in antennae, lip, foot muscle, testis, ovary, hepatopancreas, kidney, gill, endometrium, and mantle.
Effects of heat and cold stress on the expression levels of CchHSPs
-
C. chinensis has a strong temperature adaptability in natural conditions and can survive in both northern and southern regions of China. In this study, we used 0 and 30 °C as the survival limit temperatures for the snails under stress conditions. In this study, we investigated the expression of seven CchHSPs, namely CchHSP10-01, CchHSP20-02, CchHSP40-03, CchHSP60-01, CchHSP70-09, CchHSP70-12, and CchHSP90-02 in the kidney, gill, and foot muscle of C. chinensis using qPCR (Supplementary Tables S7−S9). Our results demonstrated that under high-temperature stress at 30 °C, the expression levels of these genes were significantly elevated in both the kidneys and foot muscle of C. chinensis. Additionally, while most genes in the gills exhibited a trend towards increased expression under similar conditions, exceptions were noted for CchHSP20-02, CchHSP40-03, and CchHSP70-12.
Under low-temperature stress at 0 °C, the expression of CchHSP70-09 and CchHSP90-02 are elevated in the foot muscle. Notably, the expression of CchHSP10-01, CchHSP40-03, and CchHSP90-02 is significantly upregulated in the gills. Furthermore, an increase in the expression of both CchHSP40-03 and CchHSP90-02 is observed in the kidneys. The correlation coefficient for the expression levels of these CchHSPs across different tissues compared to the control group indicates that their expression patterns are tissue-specific and markedly influenced by temperature stress (Fig. 6).
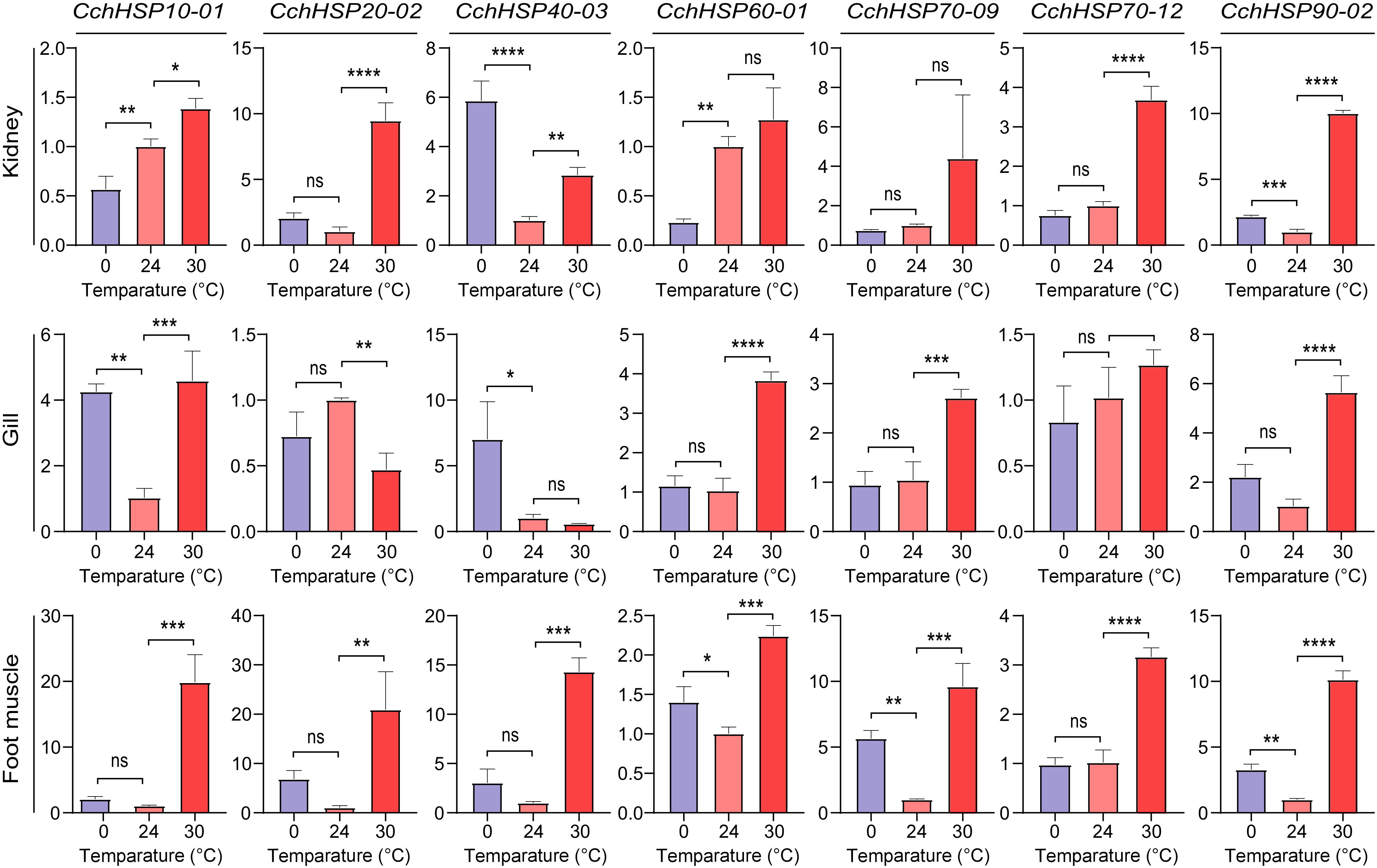
Figure 6.
Relative expression levels of CchHSPs in kidney, gill, and foot muscles under temperature stress. The horizontal axis is the temperature of temperature stress and the vertical axis is the relative expression level of genes.
-
In this study, we utilized Hidden Markov Model (HMM) and homologous blast methods to identify the HSP gene family in C. chinensis. According to the physicochemical properties, the pI of CchHSPs ranges from 4.91 to 10.04, with a high proportion (77.78%) being acidic proteins which resembled the findings in P. canaliculata[13], suggesting that CchHSPs primarily function under acidic conditions. Almost all CchHSPs are hydrophilic, and hydrophilic interactions play an important role in protein-protein association, assembly, the process of protein folding, and molecular recognition[40,41]. Subcellular localization analysis shows that most CchHSPs are located in the cytoplasm, which is crowded with macromolecules. These proteins primarily perform functions such as protein folding and cell signaling in this cellular compartment[42]. In addition, four sets of tandem duplicated genes were identified in this study on chromosomes 1, 2, 4, and 7. Tandem duplication is crucial for the expansion and evolution of gene families in organisms, as well as the generation of new functions[43,44].
Gene structure analysis reveals that each family contains specific motifs and different gene domains, reflecting the functional differences among the subfamilies. There are significant variations in the size, number, and positions of introns and exons among CchHSPs. Notably, some genes lack introns, which resembled the situation in P. canaliculate[13]. Some sister branches shared motifs, indicating that genes in the same subfamily have similar gene structures and functions. Significant motif differences among different subfamilies indicate potential functional differences among subfamilies. Most members within the same clade contain identical domains, suggesting they may perform similar functions. However, different domains can exist within the same family. For example, the domains of CchHSP90s vary, likely due to the development of new functions in family members during evolution.
The secondary structure of CchHSPs mainly comprises alpha helices, extended strands, and random coils. HSPs regulate the folding, localization, accumulation, and degradation of protein molecules[11]. A previous study reported that the alpha-helix structure of HSPs aids in faster protein folding[45], while the extended strand structure forms intermolecular hydrogen bonds which are essential for chaperone function and establishes hydrophobic contacts with small HSP proteins, maintaining protein oligomerization and stability, thus fulfilling the protein chaperone function of HSPs[7,46,47]. CchHSP60s, CchHSP70s, and CchHSP90s have a high proportion of alpha helices, which may facilitate rapid protein folding. CchHSP40s can collaborate with CchHSP70s to perform chaperone functions. This collaboration is facilitated by the J domain of the HSP40 subfamily protein, which is composed of alpha-helices that interact with HSP70 proteins to form a binary complex[48]. According to the prediction of three-dimensional structure, it is speculated that genes with similar structures in sister branches of the same subfamily may have similar functions.
The phylogenetic tree of CchHSPs within the species in this study is divided into three branches: one branch consists of members of the CchHSP40 subfamily; one branch consists of members of the CchHSP70 subfamily; and one branch consists of members of other subfamilies. In the interspecific phylogenetic tree, members of various families from seven species cross-clustered together, suggesting that HSP gene subfamilies were formed before the divergence of these species.
Most CchHSPs exhibit tissue-specific expression. Previous studies have shown that HSP10 is involved in the regulation of apoptosis in ovarian granulosa cells[49], and CchHSP10 may be involved in the regulation of follicular and oocyte development and maturation in C. chinensis. CchHSP20-06, CchHSP20-07, and CchHSP20-08 are highly expressed in the foot muscles. HSP20, a chaperone protein abundantly expressed in smooth muscle, is a crucial regulator of muscle contraction, cell migration, and cell survival[50]. This high expression in the foot muscles indicates that CchHSP20s may have a potential regulatory role in smooth muscle contraction in C. chinensis. Additionally, some CchHSP20s are highly expressed in the hepatopancreas. In Procambarus clarkii, immune elicitors significantly enhance HSP20 expression in the hepatopancreas[51], suggesting that CchHSP20s may participate in immune activities in this organ. CchHSP60s are predominantly expressed in the kidney, indicating a potential role in the complex communication network between the immune system and the body[52]. HSP60 acts as a ligand that activates B cells and regulates effector T cell activity through innate TLR2 signaling[53]. The HSP60 epitope is also recognized by T cells in both health and autoimmune diseases[54]. HSP70 and HSP90 are primarily involved in the metabolism of non-reproductive organ epithelium, spermatogenic cells, and testicular mesenchymal cells[55]. Some CchHSP70s and CchHSP90s are highly expressed in the testis, implying that these family members may play critical regulatory roles in testis development and spermatogenesis in C. chinensis. A previous study has shown that HSP90 is intensely expressed in prespermatogonia and spermatogonia in rabbit testes[56].
CchHSP70-9, CchHSP70-12, and CchHSP90-2 are key nodes in the PPI network, and they all interact with NEDD8, A0A2T7PA24, and A0A2T7NZM9 proteins. Previous studies have revealed that NEDD8 is a ubiquitin-like protein essential for protein degradation and regulation within cells[57]. It is speculated that these interactions may regulate cell proliferation and apoptosis through the NEDD8 gene. The Elongin-C, primarily involved in the metabolic processes of various substances, can act as the ubiquitin ligase E3 necessary for ubiquitin-mediated protein degradation[58,59]. It shows strong interactions with CchHSP70-9, CchHSP70-12, CchHSP90-2, and NEDD8, suggesting a joint involvement in the ubiquitin-mediated protein degradation process.
The findings also demonstrated that there is also a strong interaction between A0A2T7PA24, A0A2T7NZM9, and the three key nodes. Both A0A2T7PA24 and A0A2T7NZM9 can bind to FK506 and are associated with prostaglandin-E synthase activity. Studies indicate that FK506 is involved in regulating the post-traumatic inflammatory process and has a neuroprotective effect on various cell populations in the central nervous system[60]. A0A2T7NZM9 contains the STI1 domain[61,62], which is present in many co-chaperone proteins and referred to as the heat shock chaperone protein binding domain. A0A2T7NZM9 received stimulation from cytokines and chemicals, so it is speculated that it is a stress-induced gene. It is hypothesized that A0A2T7NZM9 receives stimulation signals, activates the synthesis of HSP, and assists in the correct folding, assembly, and refolding of proteins along with A0A2T7PA24 and HSPs, which further binds to FK506 to reduce nerve damage and cell damage[60].
Numerous studies have demonstrated that HSPs play a crucial role in response to both high- and low-temperature stress. For instance, in Nacella concinna, the expression levels of HSP70 were elevated under heat stress and significantly reduced under cold stress[63]. Similarly, in this study, HSP70 showed increased expression under heat stress, with upregulation observed in the foot under cold stress, while other tissues exhibited minimal changes. In Laternula elliptica, the expression level of HSP70 in gills significantly increases under heat stress[64]. For Pinctada martensii, the expression level of HSP40 in the gills initially rise significantly under heat stress and then returns to baseline levels after 48 h[65]. In this study, CchHSP40-03 was significantly upregulated in the kidney and foot muscle after three days of heat stress, while the expression level in gills remained similar to the normal level, likely returning to baseline after the stress period which is similar to what happened in P. martensii. The expression of CchHSP90-02 was up-regulated under cold stress and heat stress, and a similar situation also appeared in the muscle and skin of Andrias davidianus, indicating that HSP90 plays an important role in coping with temperature stress[66]. The expression levels of HSP70 and HSP90 in land snail species also increased continuously during the first 15−20 d after exposure to 34 °C[67]. It can be seen that the HSP gene family plays an important role in the adaptability to temperature. The expression of CchHSPs is more sensitive to heat stress than cold stress, particularly in the foot muscle. Changes in the expression levels of CchHSP40-3 in kidney and gill under cold stress were significant and even greater than the effects produced by heat stress. HSP40 may play a key role in the adaptation of C. chinensis to cold environments.
-
In this study, we identified the HSP gene family associated with stress responses in C. chinensis at the whole-genome level. A total of 36 genes were identified, encompassing the HSP10, HSP20, HSP40, HSP60, HSP70, and HSP90 subfamilies distributed across nine chromosomes, and some subfamilies exhibited gene duplication events.
Structural analysis indicated that each subfamily possesses specific sequence patterns and distinct structural domains, reflecting functional differences among them. Significant variations were observed in terms of size, number, and location of introns and exons within CchHSPs. The secondary structure of CchHSPs predominantly consisted of alpha helices, extended chains, and random coils.
Protein interaction analysis demonstrated that genes such as CchHSP90-02, CchHSP70-12, and CchHSP70-09 occupied central positions within interaction networks, suggesting their core roles in cellular processes. It is worth noting that the expression levels of HSP genes in C. chinensis are significantly different in different tissues, indicating that these genes have different functional roles. The qPCR experiments assessing the expression profiles of the HSP gene family under cold and heat stress confirmed that CchHPS70-09, CchHPS70-12, and CchHPS90-02 were highly expressed during these stresses, underscoring their important roles in stress response mechanisms.
This study provides a foundational data set and theoretical support for further investigations into the regulation of HSPs in relation to environmental adaptability, particularly temperature. Through these studies, we aim to offer more comprehensive theoretical insights into the molecular adaptation mechanisms and environmental resilience of C. chinensis.
This research was supported by the National Natural Science Foundation of China (32070526 and 32270487), the Key Research and Development Programme of Jiangsu Province (BE2020673), and the Major Project of Jiangsu Higher Education Institutions for Basic Science (Natural Science) Research (24KJA240003).
-
The authors confirm contribution to the paper as follows: study conception and design: Wu X, Wang G, Chen H; data collection: Sun Q, Wei Z, Xuan F, Zhao S; analysis and interpretation of results: Yang W, Chen S, Jin W, Ji Q; draft manuscript preparation: Wu X, Wang G, Yang W, Sun Y; project organization and supervision: Tang B, Zhang D. All authors reviewed the results and approved the final version of the manuscript.
-
The transcriptome datasets generated and analyzed during the current study are available in the NCBI database: PRJNA1134349. The genomes of B. aeruginosa have been deposited in the Genome Warehouse at the NGDC, BIG, CAS / CNCB (BioProject: PRJCA032440).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Xiaoxiao Wu, Wenqi Yang
- Supplementary Table S1 Primers used for qPCR assays.
- Supplementary Table S2 Sequence list of the HSP genes identified from the genomes of B. aeruginosa, B. glabrata, C. chinensis, P. maculata, L. ventricosus, P. pellucida, and A. fulica.
- upplementary Tables S3 HSP genes identified in the genomes of four species.
- Supplementary Table S4 Location of CchHSPs in four clusters of tandem duplications.
- Supplementary Table S5 String interactions of CchHSPs with P. canaliculata as the model.
- Supplementary Table S6 String interactions annotations of CchHSPs with P. canaliculata as the model.
- Supplementary Table S7 The relative gene expression of CchHSPs under different temperature in kidney. Each value represents as Mean ± SE, n = 3 per group.
- Supplementary Table S8 The relative gene expression of CchHSPs under different temperature in gill. Each value represents as Mean ± SE, n = 3 per group.
- Supplementary Table S9 The relative gene expression of CchHSPs under different temperature in foot muscle. Each value represents as Mean ± SE, n = 3 per group.
- Supplementary Fig. S1 Sequence logos for the conserved motifs of CchHSPs.
- Supplementary Fig. S2 The three-dimensional structure prediction of CchHSPs.
- Supplementary Fig. S3 Protein-protein interactions of CchHSPs.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wu X, Yang W, Chen R, Chen S, Sun Q, et al. 2025. Comprehensive genomic profiling and expression dynamics of the HSP gene family in Cipangopaludina chinensis under temperature stress. Genomics Communications 2: e009 doi: 10.48130/gcomm-0025-0009 

Comprehensive genomic profiling and expression dynamics of the HSP gene family in Cipangopaludina chinensis under temperature stress
- Received: 23 December 2024
- Revised: 03 April 2025
- Accepted: 11 April 2025
- Published online: 09 May 2025
Abstract: Heat shock proteins (HSPs) play a crucial role in the survival of organisms by facilitating their response to various stress conditions. Cipangopaludina chinensis is an important aquaculture species in China that is highly resistant to temperature changes. To better understand the adaptation mechanisms of C. chinensis to temperature fluctuations, we identified and analyzed the HSPs in C. chinensis (CchHSPs). The 36 identified CchHSPs are distributed on nine chromosomes, and the proteins they encode predominantly exhibit alpha-helix structures. In phylogenetic analyses, each HSP subfamily formed a monophyletic clade. Different CchHSPs showed specific high expression in different tissues. In addition, CchHPS70-09, CchHPS70-12, and CchHPS90-02 were identified as hub genes in the protein-protein interaction network, and the significant elevation of the expression of these genes in foot muscle under heat stress further validated their importance for the adaptation of C. chinensis to environmental stress. This study advances our understanding of the mechanisms underlying the adaptability and stress resistance conferred by the HSP gene family in C. chinensis. These insights will facilitate future efforts aimed at designing molecular markers and conducting molecular breeding programs for this economically valuable species.
-
Key words:
- Heat shock protein /
- Freshwater snail /
- Stress resistance /
- Gastropod /
- Gene family