









-
MicroRNAs (miRNAs) are small, endogenous, non-coding RNA molecules, typically 20−22 nucleotides (nt) in length, that play critical regulatory roles in various biological processes, including development, abiotic stress responses, signal transduction, and pathogen resistance[1−5]. Extensive studies have demonstrated that miRNAs are ubiquitous in plants and exhibit high conservation across species. To date, a large number of miRNAs have been identified and cataloged in miRBase (version 22.0), highlighting their essential functions in numerous metabolic processes, particularly fruit development[6−9]. For instance, overexpression of miR156b has been linked to abnormal fruit morphology in tomato plants[10]. Additionally, miR156/157 and miR172 have been shown to regulate tomato ripening by targeting key regulators such as CNR and AP2a. In strawberries, miR159a targets FaGAMYB, which is involved in the transition from receptacle development to ripening[11]. Similarly, miR73 modulates ABI5, a regulator of strawberry ripening through the ABA signaling pathway[12].
MiRNAs have been identified by predictive bioinformatics analyses and Sanger sequencing[13,14]. Recently, rapid developments in high-throughput sequencing have facilitated the application of transcriptome profiling in identifications in various fruit-producing species such as apple[15], grape[16,17], peach[18,19], pomegranate[20], date palm[21], sweet orange[22], and blue berry[23]. Similarly, degradome sequencing has revealed miRNA-mRNA target pairs which are prevalent during fruit development[15,22]. Moreover, the combination analyses of miRNAome and degradome have verified various roles of miRNAs during fruit development.
Peach (Prunus persica L.) is a highly valued deciduous fruit crop, prized for its delicious and nutritious fruit, and is widely cultivated in temperate regions worldwide. As a member of the Rosaceae family, which comprises over 3,000 species across approximately 110 genera[24]. Given their small genome size of approximately 300 Mb and relatively short reproductive time, peaches have been considered a plant genome model for Rosaceae[25,26]. The draft genome of peach (peach v2.0, derived from the 'Lovell' haploid) was sequenced by the International Peach Genome Initiative and is publicly accessible in the Genome Database for Rosaceae (
www.rosaceae.org/peach/genome ). Peach fruit development follows a typical double-sigmoid growth pattern[27], characterized by three distinct stages: an initial phase of exponential fruit size increase (FWI), a central lag phase with reduced growth rates (FWII), and a final phase of rapid regrowth (FWIII). Throughout these stages, the external appearance and internal physiological processes of the fruit undergo significant changes, with variations in fruit size, color, sugar content, organic acid levels, and hormone dynamics reflecting the expression profiles of specific genes and their regulators. Although miRNAs have been identified in various peach organs, including leaves, winter buds, roots, stems, flowers, and fruits[18,28−30], comprehensive analyses of miRNAs and their target genes across the three key stages of peach fruit development remain limited. Previous studies relied on a single mixed library to identify peach fruit miRNAs and their targets[18], underscoring the need for more detailed investigations[18].In contrast to previous approaches, we utilized three sRNA libraries and their corresponding degradome libraries, each representing a distinct developmental stage, to identify and characterize miRNAs and their targets at each phase of peach fruit development. Furthermore, we conducted a comprehensive analysis of differentially expressed miRNAs and their targets across these stages. Our findings provide valuable insights into the regulatory roles of miRNAs in controlling peach fruit development, shedding light on the molecular mechanisms underlying this process.
-
After flowering, fruit samples were collected from 10-year-old 'Baifeng' peach trees cultivated under standard field conditions in Suzhou, China. Sampling was performed at 10, 20, 30, 40, 50, 63, 75, 87, and 95 days post-anthesis (DPA). Nine 'Baifeng' trees were evenly divided into three groups, with each group serving as a technical replicate containing three biological replicates. On each sampling date, single-bearing shoots were selected from each tree. The longitudinal and transverse diameters of the fruits were measured using vernier calipers, after which the pulp was separated, immediately snap-frozen in liquid nitrogen, and stored at −80 °C until further use.
Construction and sequencing of sRNA and degradome libraries
-
After identifying three key sampling points on the double-sigmoid growth curve of 'Baifeng' peaches, we selected DPA20, DPA50, and DPA75 as representative stages of FWI, FWII, and FWIII, respectively, for sRNA and degradome sequencing. Total RNA was extracted from fruit pulp samples at DPA20, DPA50, and DPA75 using TRIzol reagent (Invitrogen, CA, USA). RNA quality was assessed using an Agilent Bioanalyzer, and sample concentrations were determined with a Nanodrop Spectrophotometer. Subsequently, three sRNA and degradome libraries were constructed and subjected to single-end sequencing (50 bp) on an Illumina HiSeq 2500 platform (Illumina Inc., San Diego, CA, USA) at LC-BIO (Hangzhou, China). The raw sequencing data for small RNA and degradome libraries have been deposited in the NCBI Short Read Archive under the following accession numbers: DPA20-miRNAs (SRX4011348), DPA50-miRNAs (SRX4011349), DPA75-miRNAs (SRX4011350), DPA20-degradome (SRX4011351), DPA50-degradome (SRX4011352), and DPA75-degradome (SRX4011353).
Identification of conserved and novel miRNAs
-
Clean data were generated using the methodology detailed by Liu et al.[9]. sRNA within the size range of 18–30 nucleotides (nt) were extracted from the clean reads and subsequently aligned to the peach reference genome (
www.rosaceae.org/organism/Prunus/persica ) using Bowtie[31]. To exclude non-miRNA sequences, the Rfam (www.sanger.ac.uk/Software/Rfam ) and GenBank (www.ncbi.nlm.nih.gov/GenBank ) databases were used to filter out coding RNAs, rRNAs, tRNAs, small nuclear RNAs (snRNAs), and small nucleolar RNAs (snoRNAs). To identify conserved miRNAs, the small RNA reads were further mapped to the annotated miRNAs in miRBase 22. After removing the reads classified as conserved miRNAs, miREvo[32], and mirdeep2[33] with default parameters were integrated to predict novel miRNAs in the peach genome.Differential expression analysis of miRNAs
-
Differentially expressed miRNAs were identified by calculating the transcripts per million (TPM) values for each miRNA as follows: (miRNA counts / total mapped reads) × 1,000,000. Differential expression analyses of DPA50 vs DPA20 and DPA75 vs DPA50 were then performed using the DEGseq (2010) R package. P-values were adjusted as described previously using q-value < 0.01 and |log2(foldchange)| > 1 as the thresholds for significant differential expression[34].
Analysis of degradome sequencing data
-
Raw sequencing reads were processed using the Illumina software to remove adaptors and low-quality reads. The extracted sequencing reads were subsequently analyzed using the Cleave Land pipeline to identify potentially miRNA-mediated cleavage targets[35]. Degradome reads were aligned to the mRNA sequences obtained from the Genome Database for Rosaceae (GDR) (
www.rosaceae.org/species/prunus_persica/genome_v1.0 ), with only perfectly matching alignment(s) for degradation analysis. All reads were reverse-complemented and mapped to the miRNAs identified in this study, allowing a maximum of four mismatches. Alignments were retained and scored if the degradome sequence position corresponded to the 10th or 11th nucleotide of the miRNA. Targets were selected and categorized into classes 0, 1, 2, 3, or 4 as previously described[22]. In addition, to analyse miRNA targets and RNA degradation patterns, t-plots were built according to the distributions of signatures (and abundances) of transcripts. All identified targets were subjected to BlastX analysis to assess sequence similarities, followed by gene ontology (GO) analysis (www.geneontology.org ) to elucidate miRNA regulatory networks. Network visualization and analysis were performed using the Cytoscape platform[36].Validation of miRNA and target gene expression using quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR)
-
To validate the sequencing results, the expression levels of five miRNAs and nine target genes were confirmed using qRT-PCR. cDNA synthesis was performed following the method described by Zhang et al.[19]. The precise sequence of ppe-miRNA was used as the forward primer and the miR-RACE 3'-primer was used as the reverse primer (Supplementary Table S1)[37]. Gene-specific primers overlapping with predicted complementary sites were designed for target gene amplification (Supplementary Table S1). qRT-PCR was conducted on a Rotor-Gene 3000 instrument (Corbett Robotics, Australia) using Rotor-Gene software version 6.1[38] and SYBR Green reaction mix (Toyobo, Osaka, Japan). U6 and RPII were used as internal controls for miRNA and mRNA normalization, respectively[39]. Each sample was analyzed with three biological replicates and three technical replicates to ensure reproducibility and reliability.
RNA ligase-mediated 5' RACE
-
RLM-5'RACE was performed using the SMARTer™ RACE cDNA Amplification Kit (Clontech Laboratories Inc., PaloAlto, CA, USA) following the manufacturer's instructions. Briefly, the RNA adapter was ligated to the purified RNAs as described by Zhang et al.[19]. The ligation products were then reverse transcribed, followed by PCR amplification using universal primers and gene-specific primers (Supplementary Table S1). The RACE products were gel-purified, cloned, and sequenced for further analysis.
-
Growing peaches display typical double-sigmoid growth patterns, but the key transition points of peach vary across peach varieties. Therefore, to elucidate the regulatory mechanisms underlying peach development, it is essential to conduct analyses at multiple time points. To address this, we collected fruit samples from 10-year-old 'Baifeng' peach trees at various stages post-flowering (Fig. 1), and measured the longitudinal and transverse diameters of the fruits.

Figure 1.
Fruit profiles at defined peach developmental stages (scale bar = 1 cm).
Our observations confirmed the double-sigmoid growth pattern in 'Baifeng' peach fruits (Supplementary Fig. S1). From 10 to 40 d post-anthesis (DPA10 to DPA40), the fruits underwent the first exponential growth phase (FWI). This was followed by a slow growth phase (FWII) from DPA40 to DPA63, characterized by hardening of the fruit core. Subsequently, from DPA63 to DPA87, the fruits entered a second exponential growth phase (FWIII). Based on these findings, we selected DPA20, DPA50, and DPA75 as representative time points for FWI, FWII, and FWIII, respectively.
Global analysis of sequences from small RNA and degradome libraries
-
To identify miRNAs involved in the development of peach fruit, three sRNA libraries representing DPA20, DPA50, and DPA75 were constructed and sequenced. In total of 12,017,340, 12,258,319, and 10,356,759 raw reads were generated from the DPA20, DPA50, and DPA75 libraries, respectively (Supplementary Table S2). After filtering out low-quality reads, adapters, junk reads, and redundant repeats, 11,429,372, 11,828,096, and 9,995,951 clean reads were obtained. Small RNAs ranging from 18 to 30 nucleotides (nt) accounted for 85.05% (DPA20), 89.79% (DPA50), and 93.64% (DPA75) of the clean reads (Supplementary Table S2) and were retained for further analysis. The length distribution of sRNAs was analyzed across the three libraries (Fig. 2). sRNAs with 21 and 24 nt were the most abundant, collectively representing an average of 75.5% of all sRNAs. However, the length distribution varied slightly among libraries: 21-nt sRNAs constituted 29.4% of the reads in the DPA75 library, while 24-nt sRNAs were predominant in the DPA20 and DPA50 libraries, accounting for 18.3% and 22.8% of the reads, respectively.
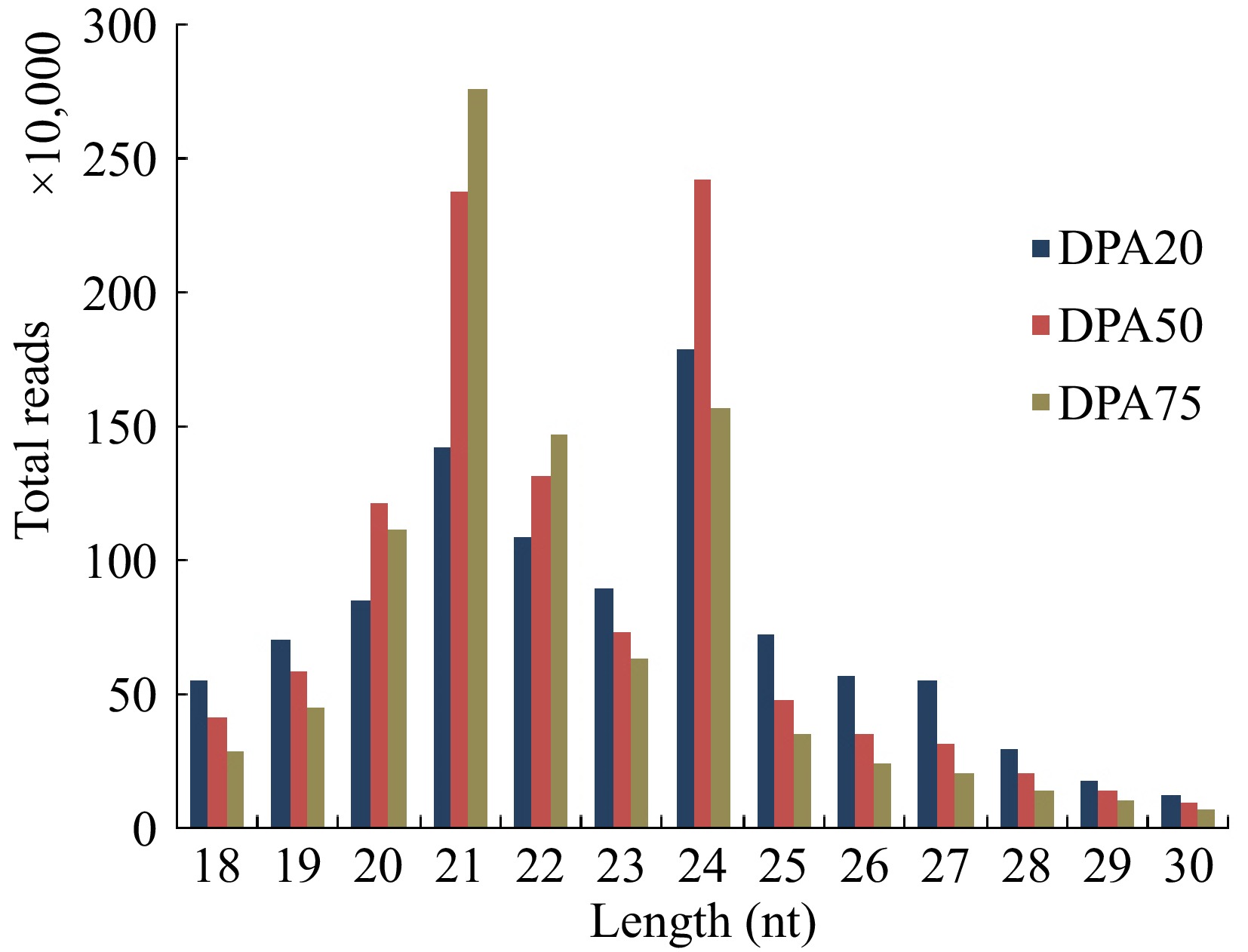
Figure 2.
Length distributions of sRNAs of peaches in the libraries at 20 d post anthesis (DPA20), DPA50, and DPA75.
To identify miRNA targets during peach fruit development, transcriptome-wide degradome sequencing was performed to detect miRNA-mediated cleavage sites, which are characterized by endonucleolytic cleavage driven by extensive, often perfect, mRNA complementarity[40]. These analyses identified 26,557,773, 24,876,000, and 31,586,572 raw reads from the DPA20, DPA50, and DPA75 libraries, respectively (Supplementary Table S2). Approximately 7.27, 6.88, and 7.69 million unique sequences were successfully mapped to the peach genome sequences, accounting for 99.29% of all unique reads (Supplementary Table S2). Among these, over 60% of the reads were mapped to peach cDNA sequences, and all mapped sequences were subsequently used to identify miRNA targets.
Identification and characterization of known and novel miRNAs in peach fruit
-
To identify fruit-specific miRNAs, mRNA sequences from DPA20, DPA50, and DPA75 fruits were aligned with known miRNAs from the publicly available database miRbase (version 22). Following a series of screening, 124 known miRNAs belonging to 75 families and 86 novel miRNAs were identified in the three peach fruit samples (Table 1; Supplementary Table S3). We further analyzed the distribution of these miRNAs among the three stages. Among the known miRNAs, 85 (68.5%) were detected in all stages, while seven miRNAs (ppe-miR5225-3p, ppe-miR5225-5p, ppe-miR6267a, ppe-miR6280, ppe-miR6282, ppe-miR6283, and ppe-miR8127-5p) were exclusively detected in DPA20 fruits, four (ppe-miR169f, ppe-miR6266a, ppe-miR6285, and ppe-miR8130-5p) were specific to DPA50 and 13 (ppe-miR156f, ppe-miR2111a, ppe-miR319b, ppe-miR6260, ppe-miR6261, ppe-miR6263, ppe-miR6271, ppe-miR6287, ppe-miR6288b-3p, ppe-miR6290, ppe-miR6292, ppe-miR6297b, and ppe-miR8128-5p) were unique to DPA75 (Fig. 3 & Table 1).
Table 1. Characteristics of know miRNAs identified from DPA20, DAP50, and DPA75 libraries in peach fruit.
miRNA ID miRNA sequences (5'-3') LT1 TPM2 DPA20 DPA50 DPA75 ppe-miR1511-5p CGUGGUAUCAGAGUCAUGUUA 21 154,692.1383 14,578.83618 17,587.11608 ppe-miR1511-3p ACCUGGCUCUGAUACCAUAAC 21 21.90796464 20.71184303 44.79237715 ppe-miR156a UGACAGAAGAAAGAGAGCAC 20 0 4.602631785 26.46822286 ppe-miR156c UGACAGAAGAGAGUGAGCAC 20 0 16.10921125 22.39618858 ppe-miR156f UGACAGAAGAUAGAGAGCAC 20 0 0 10.18008572 ppe-miR159a UUUGGAUUGAAGGGAGCUCUA 21 217,885.6623 131,131.2809 98,590.05813 ppe-miR159b CUUGGAUUGAAGGGAGCUCCA 21 381,669.606 15,059.8112 12,138.73421 ppe-miR160a UGCCUGGCUCCCUGUAUGCCA 21 1,084.44425 333.6908044 177.1334915 ppe-miR162 UCGAUAAACCUCUGCAUCCAG 21 9,792.860194 7,748.53061 4,200.303367 ppe-miR164a UGGAGAAGCAGGGCACGUGCA 21 613.4230099 2,285.206681 804.2267716 ppe-miR164d UGGAGAAGCAGGGCACAUGCU 21 10.95398232 2.301315892 0 ppe-miR166a UCGGACCAGGCUUCAUUCCCC 21 18,358.87437 7,847.487193 3,273.915566 ppe-miR167a UGAAGCUGCCAGCAUGAUCUA 21 1,424.017702 352.1013315 677.9937087 ppe-miR167c UGAAGCUGCCAGCAUGAUCUGA 22 3,768.169918 844.5829325 659.6695544 ppe-miR167d UGAAGCUGCCAGCAUGAUCUUA 22 1,577.373454 342.896068 494.7521658 ppe-miR168 UCGCUUGGUGCAGGUCGGGAA 21 492.9292044 92.0526357 89.5847543 ppe-miR169f UAGCCAAGGAUGACUUGCCUGC 22 0 2.301315892 0 ppe-miR171b UUGAGCCGCGCCAAUAUCACU 21 120.4938055 23.01315892 30.54025715 ppe-miR171d-5p UGUGAUAUUGGUUCGGUUCAUA 22 10.95398232 11.50657946 22.39618858 ppe-miR171d-3p CGAGCCGAAUCAAUAUCACUC 21 10.95398232 4.602631785 16.28813715 ppe-miR171e UUAUUGAACCGGACCAAUAUC 21 10.95398232 2.301315892 0 ppe-miR171f UGAUUGAGCCGUGCCAAUAUC 21 295.7575226 133.4763218 67.18856573 ppe-miR171h UUGAGCCGCGUCAAUAUCUCC 21 635.3309746 211.7210621 24.43220572 ppe-miR172a-3p AGAAUCUUGAUGAUGCUGCAU 21 120.4938055 20.71184303 12.21610286 ppe-miR172c GGAAUCUUGAUGAUGCUGCAU 21 32.86194696 4.602631785 16.28813715 ppe-miR172d GGAAUCUUGAUGAUGCUGCAG 21 65.72389392 2.301315892 4.072034287 ppe-miR2111a UAAUCUGCAUCCUGAGGUUUA 21 0 0 2.036017143 ppe-miR319a UUGGACUGAAGGGAGCUCCC 20 164.3097348 29.9171066 171.02544 ppe-miR319b UAGCUGCCGAGUCAUUCAUCCA 22 0 0 2.036017143 ppe-miR319c UUUGGACUGAAGGGAGCUCC 20 54.7699116 29.9171066 24.43220572 ppe-miR319d CUUGGACUGAAGGGAGCUCCC 21 251.9415934 20.71184303 16.28813715 ppe-miR319e CUUGGACUGAAGGGAGCUCCU 21 328.6194696 36.82105428 81.44068573 ppe-miR319f UUUGGACUGAAGGGAGCUCUC 21 109.5398232 126.5723741 77.36865144 ppe-miR3627-5p UCGCAGGAGAGAUGGCACUGUC 22 43.81592928 6.903947677 2.036017143 ppe-miR390 AAGCUCAGGAGGGAUAGCGCC 21 153.3557525 2.301315892 0 ppe-miR393a CAUCCAAAGGGAUCGCAUUGA 21 262.8955757 87.45000391 136.4131486 ppe-miR393b UCCAAAGGGAUCGCAUUGAUC 21 306.711505 128.87369 215.8178172 ppe-miR394a UUGGCAUUCUGUCCACCUCC 20 3,308.102661 3,516.410684 3,249.483361 ppe-miR394b UUGGCAUUCUGUCCACCUCC 20 54.7699116 39.12237017 44.79237715 ppe-miR395a-3p CUGAAGUGUUUGGGGGGACCC 21 898.2265503 2,908.863288 1,667.49804 ppe-miR395c CUGAAGUGUUUGGGGGAACUC 21 65.72389392 174.9000078 215.8178172 ppe-miR396a UUCCACAGCUUUCUUGAACGU 21 6,364.263728 59,399.2645 71,602.65089 ppe-miR396b UUCCACAGCUUUCUUGAACUU 21 61,791.41427 484,650.223 540,299.9053 ppe-miR396c UUCCACAGCUUUCUUGAACUG 21 12,421.81595 118,018.3829 109,040.9341 ppe-miR397 UCAUUGAGUGCAGCGUUGAUG 21 43.81592928 85.14868802 46.8283943 ppe-miR398a-5p GGAGCGACCUGGGAUCACAUG 21 6,484.757534 11,021.00181 2,640.714235 ppe-miR398a-3p UGUGUUCUCAGGUCGCCCCUG 21 109.5398232 71.34079267 59.04449715 ppe-miR398b CGUGUUCUCAGGUCGCCCCUG 21 1,654.05133 35,502.40027 37,275.40186 ppe-miR398c UGUGUUCUCAGGUCACCCCUU 21 32.86194696 55.23158142 22.39618858 ppe-miR399a CGCCAAAGGAGAGUUGCCCUU 21 0 34.51973839 114.01696 ppe-miR399b UCUGCCAAAGGAGAAUUGCCC 21 32.86194696 27.61579071 73.29661716 ppe-miR399c UGCCAAAGAAGAGUUGCCCUA 21 0 11.50657946 8.144068573 ppe-miR403 UUAGAUUCACGCACAAACUCG 21 230.0336287 191.0092191 425.5275829 ppe-miR477-5p ACUCUCCCUCAAAGGCUUCUAG 22 0 6.903947677 8.144068573 ppe-miR477-3p CGAAGCCUUUGGGGAGAGUAA 21 87.63185856 561.5210778 486.6080972 ppe-miR477a-3p GUUGGGGGCUCUUUUGGGACG 21 65.72389392 987.2645179 787.9386344 ppe-miR477a-5p UCCCUCAAGGGCUCCCAAUAUU 22 0 64.43684499 103.8368743 ppe-miR482a-3p UUUCCGAAACCUCCCAUUCCAA 22 3,833.893812 697.2987154 238.2140058 ppe-miR482a-5p GGGUGAGAGGUUGCCGGAAAGA 22 449.1132751 98.95658337 42.75636001 ppe-miR482b-5p GGAAUGGGAGGAUUGGGAAAA 21 12,750.43542 3,960.564651 2,972.585029 ppe-miR482b-3p CUUCCCAAACCUCCCAUUCCUA 22 32.86194696 4.602631785 10.18008572 ppe-miR482c-5p GGAAUGGGCUGUUUGGGAUG 20 755.8247801 409.6342289 191.3856115 ppe-miR482c-3p UUCCCAAGCCCGCCCAUUCCAA 22 3,428.596466 554.6171301 555.8326801 ppe-miR482e UUGCCUAUUCCUCCCAUGCCAA 22 942.0424795 849.1855643 572.1208173 ppe-miR482f UCUUUCCUACUCCACCCAUUCC 22 8,839.863732 10,813.88338 16,467.30665 ppe-miR5225-3p UCAUCUCUCCUCGACUGAA 19 21.90796464 0 0 ppe-miR5225-5p UCUGUCGUAGGAGAGAUGGCGC 22 32.86194696 0 0 ppe-miR530 UCUGCAUUUGCACCUGCACCU 21 87.63185856 23.01315892 30.54025715 ppe-miR535a UGACAACGAGAGAGAGCACGC 21 76.67787624 62.1355291 12.21610286 ppe-miR535b UGACGACGAGAGAGAGCACGC 21 197.1716818 186.4065873 77.36865144 ppe-miR6257 UCUUAACUGUUGGAUUAGGCU 21 10.95398232 11.50657946 2.036017143 ppe-miR6258 UUCCAGCUGUAAAGAUCAAGA 21 10.95398232 6.903947677 12.21610286 ppe-miR6260 UGGAGUGAGAGAAUGGGAGGU 21 0 0 6.10805143 ppe-miR6261 AAGUGAUUAUAUGGAGAAGCAC 22 0 0 2.036017143 ppe-miR6263 AAGUGGACAAAAGGGGAGUGG 21 0 0 2.036017143 ppe-miR6264 AUGCCUAUGGACACGUGUCAA 21 10.95398232 4.602631785 6.10805143 ppe-miR6265 UUGAACUUUGACCCGAUUCGCAU 23 21.90796464 4.602631785 2.036017143 ppe-miR6266a UAAAUGCAGGGGCAAAAUGAU 21 0 2.301315892 0 ppe-miR6267a UAGAGAGGUGGUACAAUUGUG 21 10.95398232 0 0 ppe-miR6267c-5p AUUGCUGAUCACCUCUCUAAU 21 21.90796464 59.8342132 185.27756 ppe-miR6267c-3p UAGAGAGAUGGUCAGCAAUGU 21 21.90796464 27.61579071 105.8728914 ppe-miR6270 UUCUGGUAUUGGAAUUUCAUU 21 142.4017702 18.41052714 40.72034287 ppe-miR6271 UCAAGAUUGAGAGAUAUAAUG 21 0 0 8.144068573 ppe-miR6274b-5p AUUUCGACUAAUAACACAAUG 21 0 2.301315892 2.036017143 ppe-miR6274b-3p UUGUGUUAUUGGCCGAAAAUAG 22 208.1256641 18.41052714 46.8283943 ppe-miR6277 UGUGUGUGGAAAGAGCGAGAC 21 21.90796464 9.20526357 12.21610286 ppe-miR6280 UUGGCAGUAAGAUUUUUGGUG 21 10.95398232 0 0 ppe-miR6281 GUUAGAGAUAGAGAGAGUGAG 21 76.67787624 52.93026553 69.22458287 ppe-miR6282 GUUGAUCGAUGUGGGAUGUUACA 23 10.95398232 0 0 ppe-miR6283 CAAAAGGGGAGUGGGAAAAUC 21 21.90796464 0 0 ppe-miR6284 UUUGGACCAUGGAUGAAGAUU 21 98.58584088 29.9171066 20.36017143 ppe-miR6285 UAGUGAAGUUUGAAUUAGGGCU 22 0 2.301315892 0 ppe-miR6287 CAAGAAGUGGAAGUUUUGGGC 21 0 0 2.036017143 ppe-miR6288a GAAAAUGACAAGUGGCUAGUU 21 0 2.301315892 2.036017143 ppe-miR6288b-3p UCAAUUAGAAAAUGAUAAGUG 21 0 0 4.072034287 ppe-miR6290 UGAAUGAGUUCAGAGAUCGUGUA 23 0 0 2.036017143 ppe-miR6292 UAUCUUUUAAUCGUUAGAUCA 21 0 0 2.036017143 ppe-miR6293 UAAGAGGCUGAUGACUAAAAC 21 65.72389392 46.02631785 61.0805143 ppe-miR6294 UGGUGUAGGCUAAUCACAAUC 21 32.86194696 20.71184303 42.75636001 ppe-miR6295 GAGGACAGAAGAUGAUUCAGC 21 32.86194696 13.80789535 20.36017143 ppe-miR6297a AAUAAUUUUUCGUCGCGCAAAAU 23 32.86194696 20.71184303 8.144068573 ppe-miR6297b GAUGUAUUGUCGUCGCGCAAAGU 23 0 0 2.036017143 ppe-miR7122a-5p UUAUACAAUGAAAUCACGGCCG 22 251.9415934 69.03947677 83.47670287 ppe-miR7122a-3p GCCGUGUUUCUUUGUAUAAAG 21 1,237.800002 1,456.73296 2,158.178172 ppe-miR7122b-3p CCGUGUUUCCUUGUAUAAAG 20 0 2.301315892 8.144068573 ppe-miR7122b-5p UUAUACAAUGAAAUCACGGUCG 22 32.86194696 46.02631785 140.4851829 ppe-miR7125-3p CGAACUUAUUGCAACUAGCUU 21 0 27.61579071 8.144068573 ppe-miR8122-5p UUCCACAGAUCUUUCCUCAUU 21 5,466.037178 1,233.505318 1,284.726817 ppe-miR8122-3p UGAAGGAAGAUUUGUGGAAAG 21 887.2725679 667.3816088 637.2733658 ppe-miR8124-3p UGGCACCAAUGAUACCAAGUUU 22 54.7699116 11.50657946 4.072034287 ppe-miR8124-5p ACUUGGUAUCUUGGUGCCGGU 21 0 2.301315892 2.036017143 ppe-miR8125 CAGGAAAGAAUGUGAUGAGUA 21 65.72389392 27.61579071 46.8283943 ppe-miR8126-5p UCUGAGUCAGAUUACUGAAUA 21 10.95398232 4.602631785 8.144068573 ppe-miR8126-3p UUCAGUAUUUUGACUCAGAA 20 21.90796464 23.01315892 42.75636001 ppe-miR8127-3p UUCAAAGGGUACAUCCACAGU 21 10.95398232 0 0 ppe-miR8128-5p AUUAGACCUCUCCCGACGAAA 21 21.90796464 43.72500196 63.11653144 ppe-miR8128-3p UCGUGGGGAGAGAUCUAAUCG 21 0 0 4.072034287 ppe-miR8130-5p GGGUUCCUUGUUGGAAGGACU 21 0 2.301315892 0 ppe-miR8131-5p AUUUCAGCUAAGUUGAGUUGU 21 10.95398232 0 2.036017143 ppe-miR8132 UCCAACGAUGGGUGACCACAA 21 10.95398232 2.301315892 10.18008572 ppe-miR8133-5p UCCUGUGCGAACGUCCAGAAG 21 87.63185856 46.02631785 28.50424001 ppe-miR8133-3p UAACUUCCGAACGUCCGCAUA 21 32.86194696 16.10921125 12.21610286 ppe-miR827 UUAGAUGACCAUCAACAAACA 21 1,084.44425 409.6342289 309.4746058 ppe-miR858 CUCGUUGUCUGUUCGACCUUG 21 10.95398232 4.602631785 2.036017143 1 Length of miRNA; 2 the expression of transcripts per million. 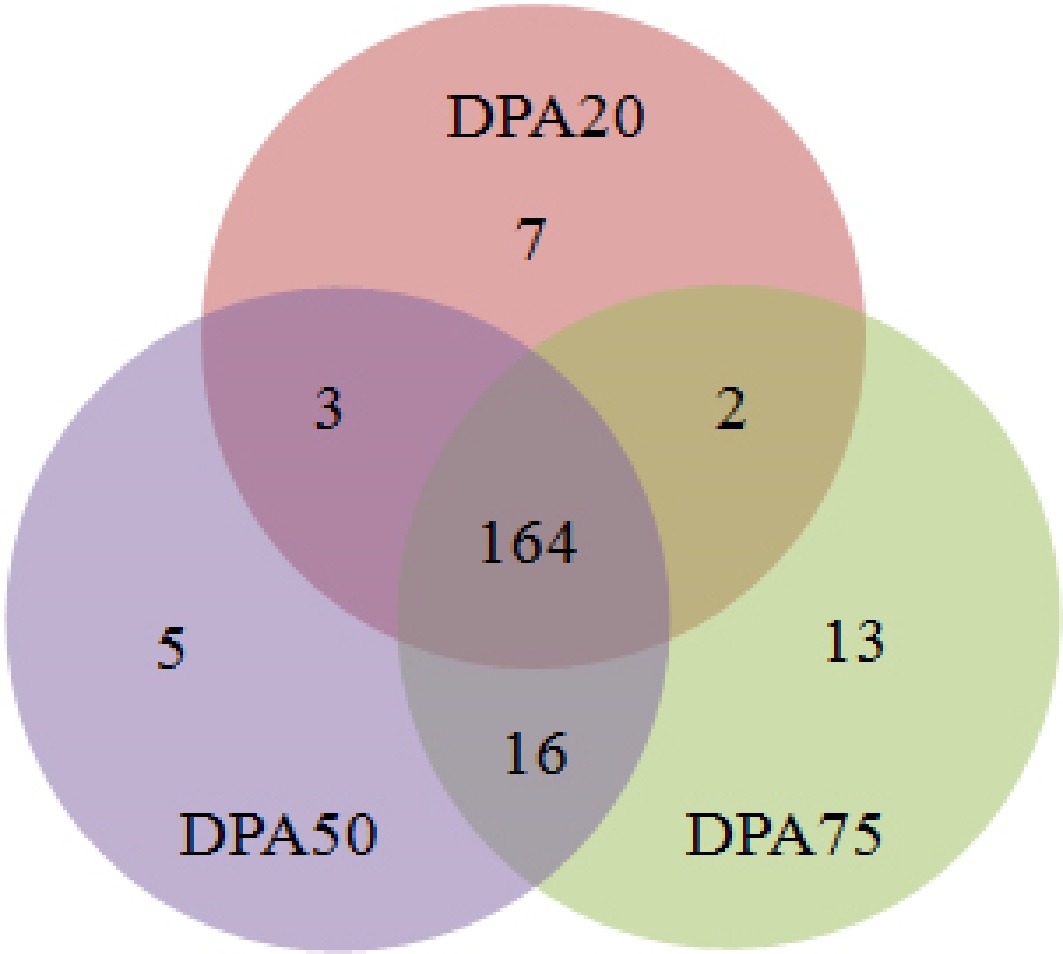
Figure 3.
Venn diagram of peach miRNAs among these libraries at DPA20, DPA50, and DPA75.
These stage-specific miRNAs are likely to play regulatory roles during peach fruit development. Notably, expression levels of some members within known miRNA families varied significantly across growth stages. For example, eight members of the ppe-miR482 family were detected, while only one miRNA was identified for each of the other 41 families (Table 1).
Among 86 novel miRNA sequences, 79 were detected in all three development stages, whereas one (novel_193) was specific to DPA50 fruit, and five (novel_100, novel_101, novel_197, novel_59, and novel_99) were detected in DPA50 and DPA75 fruits but not in DPA20. The characteristics of all 86 novel miRNAs are summarized in Supplementary Table S3. The lengths of these novel miRNAs varied from 20 to 25 nt, with the majority (64.0%) being 24 nt in length (64.0%). The precursor lengths varied from 52 to 293 nt, with an average minimum free energy (MFE) of −71.70 kCal/mol. Nucleotide bias analysis revealed that novel miRNAs with a uridine (U) residue at their 5' end were the most frequent (54.17%) among 20 to 25-nt miRNAs (Supplementary Fig. S2)[41].
Spatiotemporal expression patterns of miRNAs during peach development
-
The abundance of miRNA reads in a library reflects the relative expression levels of miRNAs. In this study, the transcripts per million (TPM) values of known and novel miRNAs ranged from 0 to 381,669.61 (ppe-miR159b) in DPA20, 0 to 484,650.22 (ppe-miR396b) in DPA50, and 0 to 540,299.90 (ppe-miR396b) in DPA75. The expression patterns of miRNAs varied widely, with TPM values spanning several orders of magnitude (Table 1; Supplementary Table S3). Among the novel miRNAs, novel_128 and novel_132 exhibited significantly higher expression levels compared to others, suggesting their potential importance in peach fruit development. Specifically, novel_128 showed the highest read counts in DPA75, while novel_132 was most abundant in DPA20. These findings indicate that novel_128 may play a critical role during the FWIII stage, whereas novel_132 is likely more important during the FWI stage. Among the known miRNAs, ppe-miR159b had the highest number of sequenced reads in DPA20, while ppe-miR396b dominated in DPA50 and DPA75, suggesting stage-specific regulatory roles for these miRNAs in peach development.
To further elucidate the roles of miRNAs during fruit development, we analyzed their abundance patterns across the three stages. As shown in Supplementary Fig. S3, 210 miRNAs were categorized into four groups based on their accumulation trends. Some miRNAs, such as ppe-miR156a, ppe-miR156c, ppe-miR399a, ppe-miR395c, and ppe-miR398b, exhibited sustained increases across all three stages. In contrast, others, including ppe-miR160a, ppe-miR164d, ppe-miR166a, and ppe-miR171f, showed sustained decreases (Supplementary Fig. S3). Notably, different members of the same miRNA family displayed opposing expression trends. For example, ppe-miR164a expression increased by 3.7-fold at DPA50, while ppe-miR164d decreased by 4.7-fold compared to DPA20 levels. These observations highlight the functional diversity of miRNAs, even within the same family, during fruit development.
To investigate the roles of miRNAs in peach fruit development, we performed differential expression analyses between adjacent stages (DPA50 vs DPA20 and DPA75 vs DPA50). MiRNAs were considered differentially expressed if |log2(fold change)| w >1 and the associated q-value was < 0.01. In the DPA50 vs DPA20 comparison, 97 miRNAs were differentially expressed, with 39 up-regulated and 58 down-regulated (Supplementary Table S3). Similarly, in the DPA75 vs DPA50 comparison, 79 miRNAs showed differential expression, with 43 up-regulated and 36 down-regulated (Supplementary Table S4). These results suggest that up-regulated miRNAs may positively correlate with fruit development, while down-regulated miRNAs may have negative regulatory roles.
Identification of miRNA target genes using degradome sequencing
-
MiRNAs can regulate multiple target genes, and conversely, a single gene can be regulated by multiple miRNAs[42]. Therefore, identifying miRNA targets is crucial for understanding their functional roles. In this study, we performed genome-wide degradome sequencing to identify miRNA-guided RNA cleavage targets. A total of 216 sliced target genes were identified for 74 known and eight novel miRNAs across the three libraries using the Cleave Land pipeline (Supplementary Tables S5). Specifically, 120, 139, and 157 target mRNAs were cleaved in the DPA20, DPA50, and DPA75 libraries, respectively. Based on the heights of the degradome peaks at each occupied transcript position, these cleaved targets were classified into the categories 0, 1, 2, 3, and 4 (Supplementary Table S5)[43]. Analysis of the cleavage products revealed that target genes regulated by single miRNAs exhibited varying expression levels across the three developmental stages, as illustrated in Fig. 4. Additionally, the cleavage frequencies of target genes varied significantly during different stages of peach development.
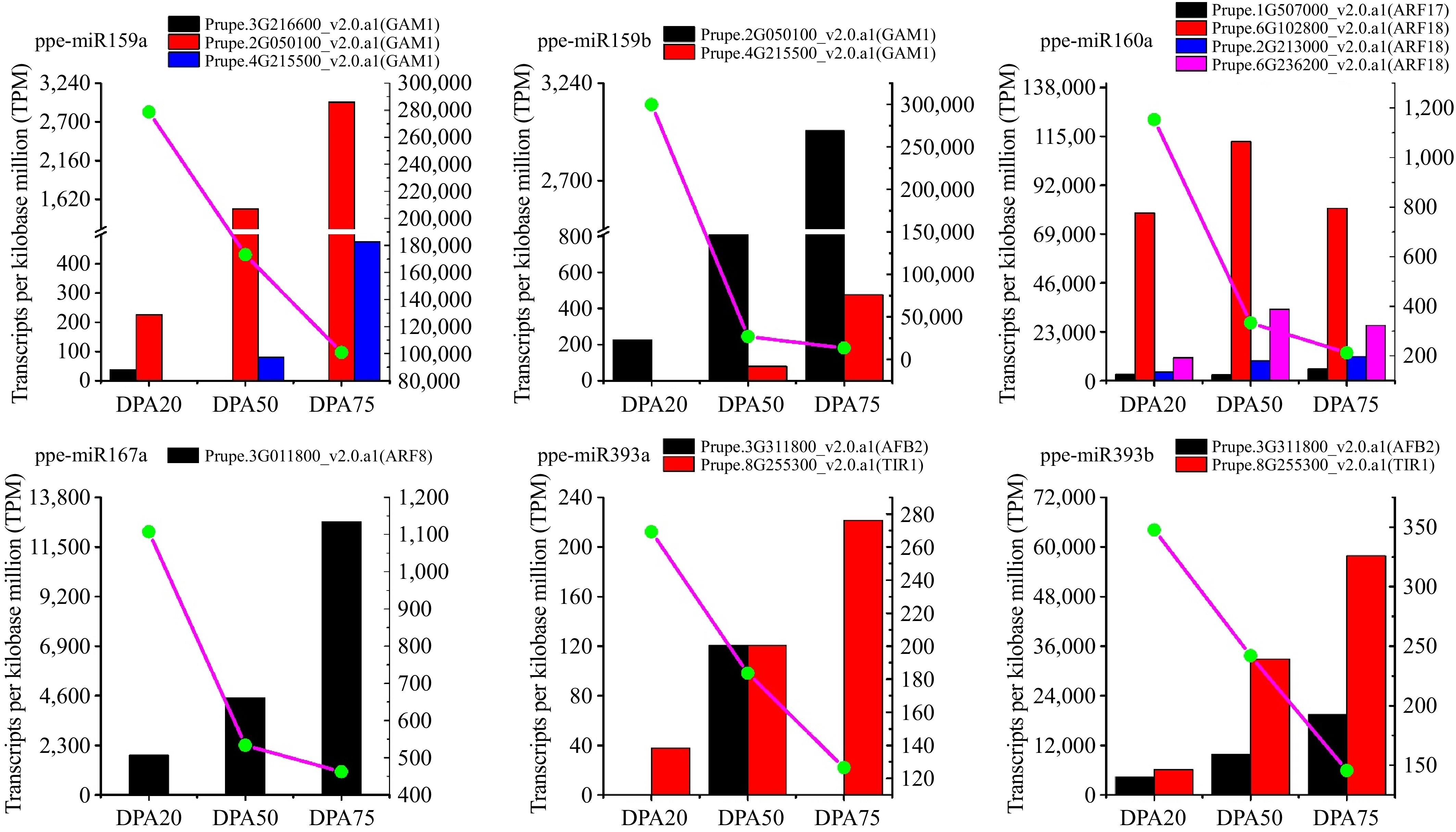
Figure 4.
Abundance of miRNA and its target genes at different peach developmental stages. Normalized sequenced read abundance, height = transcripts per kilobase million reads (TPM) on the y-axis. The histogram represents the abundance of miRNA target genes and the dot linked by a line represent the abundance of miRNA.
Among all target mRNAs which are cleaved by miRNAs, three MYB domain protein 30 were found to be the targets of ppe-miR159a, but not all the corresponding cleavage products were found at the three developmental stages (Supplementary Table S5). Notably, the cleavage products of Prupe.2G050100_v2.0. a1(GAM1) are present at all stages in greater abundance than Prupe.3G216600_v2.0.a1(GAM1) and Prupe.4G215500_v2.0.a1(GAM1) (Fig. 4; Supplementary Table S5). These observations differ from the findings of previous reports which failed to detect the cleavage products of the miR159a[44], likely reflecting different species and developmental stages. The miR160 family is known to target transcription factor genes of the ARF family, which are involved in the auxin signaling pathway during plant development[45]. Four ARF genes with cleavage products were detected across all three developmental stages, including Prupe.1G507000_v2.0.a1, which has been previously validated[44]. For ppe-miR167a, a different ARF family member (Prupe.3G011800_v2.0. a1) was identified as a target (Supplementary Table S5; Fig. 5), suggesting that ppe-miR167a also involved in auxin signaling during peach development.
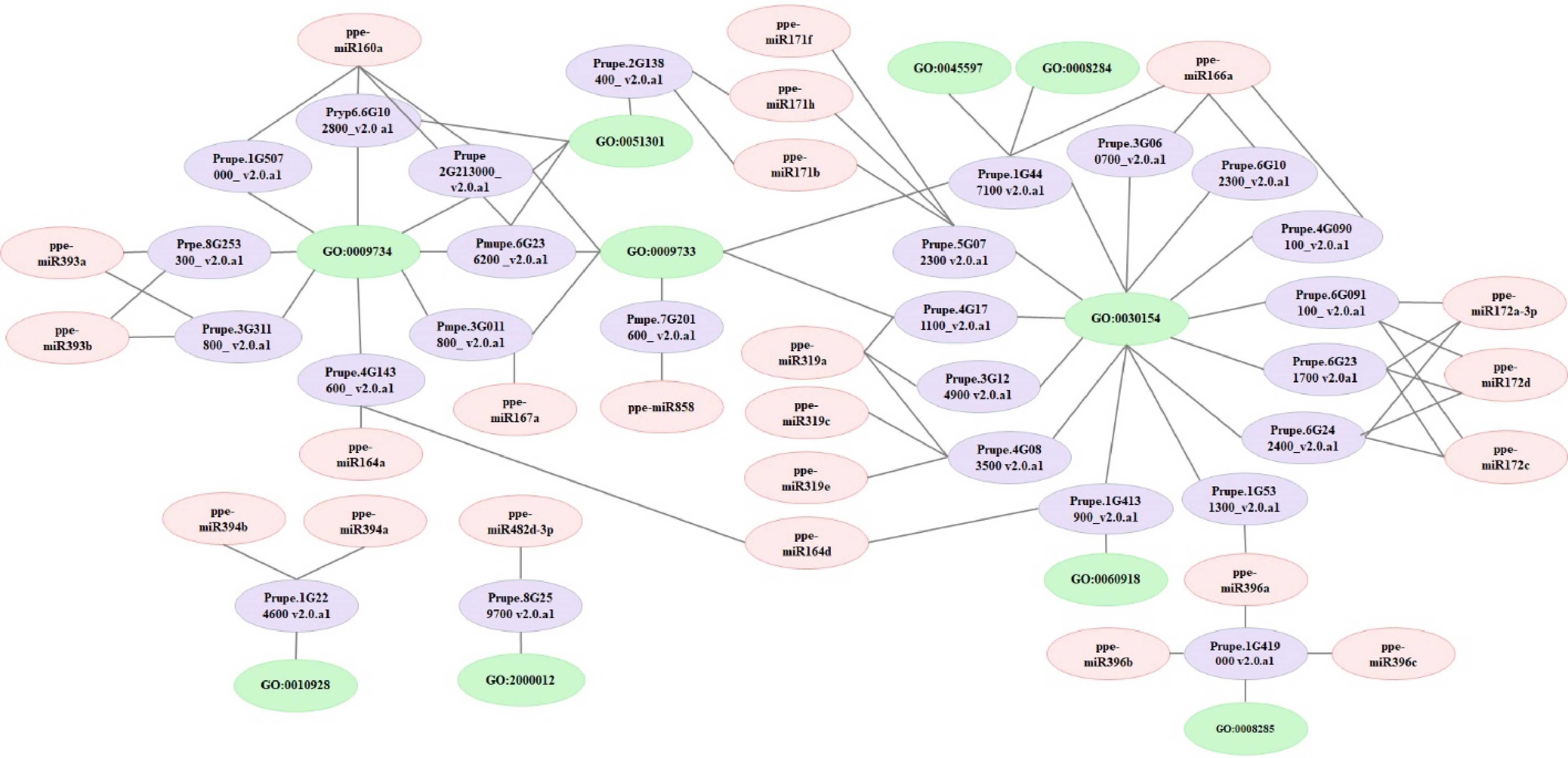
Figure 5.
Network analysis of miRNAs, targets, and GO terms involved in auxin signalling and fruit enlargement. Network analyses were performed using the Cytoscape network platform.
In tomatoes, TIR1 and two TIR1 homologs have been identified as targets of miR393 family[41]. Here we found that TIR1 (Prupe.8G253300_v2.0.a1) and AFB2 (Prupe.3G311800_v2.0.a1) are targeted by ppe-miR393a and ppe-miR393b. However, the cleavage products of ppe-miR393b target genes were significantly more abundant, while no cleavage products of AFB2 (Prupe.3G311800_v2.0.a1) were detected for ppe-miR393a. These results suggest that ppe-miR393b plays a critical role in peach development, particularly during the FWI stage, with peak degradation occurring around 20 DPA, consistent with findings in tomatoes[42]. Additionally, two MYB domain protein genes were identified as targets of ppe-miR159b, similar to ppe-miR159a. Both miRNAs were found to cleave Prupe.2G050100_v2.0.a1 (GAM1) and Prupe.4G215500_v2.0.a1 (GAM1), confirming that a single miRNA can regulate multiple genes and that multiple miRNAs can collectively regulate a single gene[42].
GO pathway analysis and identification of fruit development-specific miRNA-mediated regulatory networks
-
We identified 216 target genes through degradome sequencing and performed Gene Ontology (GO) pathway analyses to elucidate their functional roles. Differentially expressed target genes in the DPA50 vs DPA20 comparison were classified into 382 GO terms, with 25 significantly enriched terms (p-value ≤ 0.05), including 13 biological processes, eight molecular functions, and four cellular components (Supplementary Table S6). Key enriched biological processes included cell differentiation (GO:0030154), auxin-activated signaling (GO:0009734), and leaf development (GO:0048366). In the cellular component category, the nucleus (GO:0005634), microtubule (GO:0005874), and endosome (GO:0005768) were predominant, while ADP binding (GO:0043531), ATP binding (GO:0005524), and metal ion binding (GO:0046872) were highly enriched in the molecular function category (Supplementary Table S6). The top 20 enriched GO terms for the DPA50 vs DPA20 comparison are illustrated in Supplementary Fig. S4a. Similarly, in the DPA75 vs DPA50 comparison, differentially expressed target genes were classified into 391 GO terms, with 27 significantly enriched terms (p-value ≤ 0.05), including 15 biological processes, nine molecular functions, and three cellular components (Supplementary Table S6). Key biological processes included transcription (GO:0006351), cell differentiation (GO:0030154), and auxin-activated signaling (GO:0009734). The cellular component category was dominated by the nucleus (GO:0005634), cytosol (GO:0005829), and extracellular region (GO:0005576), while ADP binding (GO:0043531), metal ion binding (GO:0046872), and DNA binding (GO:0003677) were highly enriched in the molecular function category (Supplementary Table S6). The top 20 enriched GO terms for the DPA75 vs DPA50 comparison are shown in Supplementary Fig. S4b. These results suggest that the targets of some miRNAs that were identified by degradome sequencing are involved in many biological processes during peach development.
Our GO analyses revealed that the most enriched target mRNAs are associated with cell differentiation (GO:0030154) and auxin-activated signaling (GO:0009734), both of which are critical for peach fruit development. Congruently, fruit development is accompanied by constant division and differentiation of cells, and a previous report showed concordant effects of auxin on fruit enlargement[46]. To investigate the relationships among cell differentiation, auxin-activated signaling, ppe-miRNAs, and their targets, we conducted network analyses using the Cytoscape platform. This analysis included 23 miRNAs and 26 target genes known to be involved in cell differentiation, cell division, auxin transport, and auxin signaling (Table 2 & Fig. 5). The network revealed that a single pathway can be regulated by multiple miRNAs, and a single miRNA can influence multiple pathways, such as the auxin-activated signaling pathway. Specifically, we identified six ppe-miRNAs (ppe-miR160a, ppe-miR164a, ppe-miR164d, ppe-miR167a, ppe-miR393a, and ppe-miR393b) targeting 11 mRNAs, including auxin response factors, auxin receptor mRNAs, and NAC domain transcriptional regulators. For example, ppe-miR164d targets Prupe.4G143600_v2.0.a1 and Prupe.1G413900_v2.0.a1, both of which are involved in auxin signaling and cell differentiation. Similarly, ppe-miR160a targets Prupe.2G213000_v2.0.a1, Prupe.6G102800_v2.0.a1, and Prupe.6G236200_v2.0.a1, which are associated with auxin-activated signaling and cell division (Table 2 & Fig. 5). These findings demonstrate that a complex miRNA-mediated regulatory network governs multiple biological processes during peach development.
Table 2. The miRNA, target genes, and GO terms information used for network analysis.
GO_Id GO_term miRNA DPA20_TPM DPA50_TPM DPA75_TPM Transcript_ID Symbol Annoation DPA20_TPM DPA50_TPM DPA75_TPM GO:0009734 Auxin-activated signaling pathway ppe-miR160a 1,084.44425 333.6908044 177.1334915 Prupe.1G507000_v2.0.a1 ARF17 Auxin response factor 3 2,936.993249 2,693.359061 5,571.987995 Prupe.2G213000_v2.0.a1 ARF18 Auxin response factor 3 4,104.259796 9,286.058852 11,365.58915 Prupe.6G102800_v2.0.a1 ARF18 Hypothetical protein 78,884.62636 112,477.8903 81,142.07518 Prupe.6G236200_v2.0.a1 ARF18 Auxin response factor 3 10,806.629 33,566.48979 26,055.37568 ppe-miR164a 613.4230099 2,285.206681 804.2267716 Prupe.4G143600_v2.0.a1 NAC021 NAC domain-containing protein 89-like 489.4988748 8,562.46985 5,160.4207 ppe-miR164d 10.95398232 2.301315892 0 Prupe.4G143600_v2.0.a1 NAC021 NAC domain-containing protein 89-like 489.4988748 8,562.46985 5,160.4207 ppe-miR167a 1,424.017702 352.1013315 677.9937087 Prupe.3G011800_v2.0.a1 ARF8 Auxin response factor 19-like 1,845.03422 4,502.331565 12,663.60908 ppe-miR393a 262.8955757 87.45000391 136.4131486 Prupe.3G311800_v2.0.a1 AFB2 Protein TRANSPORT INHIBITOR RESPONSE 1-like 0 120.5981669 0 Prupe.8G253300_v2.0.a1 TIR1 Hypothetical protein 37.6537596 120.5981669 221.6131589 ppe-miR393b 306.711505 128.87369 215.8178172 Prupe.3G311800_v2.0.a1 AFB2 Protein TRANSPORT INHIBITOR RESPONSE 1-like 4,367.836113 9,848.850297 19,406.98092 Prupe.8G253300_v2.0.a1 TIR1 Hypothetical protein 6,137.562814 32,923.29957 57,841.03448 GO:0010928 regulation of auxin mediated signaling pathway ppe-miR394b 54.7699116 39.12237017 44.79237715 Prupe.1G224600_v2.0.a1 FBX6 − 978.9977495 4,140.537064 7,408.211312 ppe-miR394a 3,308.102661 3,516.410684 3,249.483361 Prupe.1G224600_v2.0.a1 FBX6 − 978.9977495 4,140.537064 7,408.211312 GO:0009733 response to auxin ppe-miR167a 1424.017702 352.1013315 677.9937087 Prupe.3G011800_v2.0.a1 ARF8 Auxin response factor 19-like 1,845.03422 4,502.331565 12,663.60908 ppe-miR160a 1,084.44425 333.6908044 177.1334915 Prupe.2G213000_v2.0.a1 ARF18 Auxin response factor 3 4,104.259796 9,286.058852 11,365.58915 Prupe.6G236200_v2.0.a1 ARF18 Auxin response factor 3 10,806.629 33,566.48979 26,055.37568 ppe-miR858 10.95398232 4.602631785 2.036017143 Prupe.7G201600_v2.0.a1 C1 Transcription factor MYB75-like 0 40.19938897 886.4526356 ppe-miR166a 18,358.87437 7,847.487193 3,273.915566 Prupe.1G447100_v2.0.a1 ATHB-8 − 414.1913556 2,170.767004 2,421.915237 ppe-miR319a 164.3097348 29.9171066 171.02544 Prupe.4G171100_v2.0.a1 PCF5 − 112.9612788 0 94.9770681 GO:0060918 Auxin transport ppe-miR164d 10.95398232 2.301315892 0 Prupe.1G413900_v2.0.a1 SF21 − 37.6537596 0 0 GO:2000012 Regulation of auxin polar transport ppe-miR482d-3p 0 0 0 Prupe.8G259700_v2.0.a1 OsI_027940 Uncharacterized protein OsI_027940-like 37.6537596 120.5981669 31.6590227 GO:0030154 Cell differentiation ppe-miR319c 54.7699116 29.9171066 24.43220572 Prupe.4G083500_v2.0.a1 TCP2 − 0 0 63.3180454 ppe-miR319e 328.6194696 36.82105428 81.44068573 Prupe.4G083500_v2.0.a1 TCP2 − 0 0 63.3180454 ppe-miR164d 10.95398232 2.301315892 0 Prupe.1G413900_v2.0.a1 SF21 − 37.6537596 0 0 ppe-miR166a 18,358.87437 7,847.487193 3,273.915566 Prupe.1G447100_v2.0.a1 ATHB-8 − 414.1913556 2,170.767004 2,421.915237 Prupe.3G060700_v2.0.a1 ATHB-15 − 414.1913556 2,170.767004 2,421.915237 Prupe.4G090100_v2.0.a1 ATHB-14 − 941.3439899 924.5859463 569.8624086 Prupe.6G102300_v2.0.a1 REV − 715.4214324 1,165.78228 1,677.928203 ppe-miR171b 120.4938055 23.01315892 30.54025715 Prupe.5G072300_v2.0.a1 SCL6 Hypothetical protein 715.4214324 442.1932787 348.2492497 ppe-miR171f 295.7575226 133.4763218 67.18856573 Prupe.5G072300_v2.0.a1 SCL6 Hypothetical protein 0 241.1963338 1,171.38384 ppe-miR171h 635.3309746 211.7210621 24.43220572 Prupe.5G072300_v2.0.a1 SCL6 Hypothetical protein 715.4214324 442.1932787 348.2492497 ppe-miR172a-3p 120.4938055 20.71184303 12.21610286 Prupe.6G091100_v2.0.a1 RAP2-7 − 37.6537596 200.9969448 284.9312043 Prupe.6G231700_v2.0.a1 AP2 Uncharacterized LOC106378603 376.537596 1,045.184113 3,609.128588 Prupe.6G242400_v2.0.a1 AP2 − 0 0 94.9770681 ppe-miR172c 32.86194696 4.602631785 16.28813715 Prupe.6G091100_v2.0.a1 RAP2-7 − 37.6537596 200.9969448 284.9312043 Prupe.6G231700_v2.0.a1 AP2 Uncharacterized LOC106378603 376.537596 1,045.184113 3,609.128588 Prupe.6G242400_v2.0.a1 AP2 − 0 0 94.9770681 ppe-miR172d 65.72389392 2.301315892 4.072034287 Prupe.6G091100_v2.0.a1 RAP2-7 − 37.6537596 200.9969448 284.9312043 Prupe.6G231700_v2.0.a1 AP2 Uncharacterized LOC106378603 376.537596 1,045.184113 3,609.128588 Prupe.6G242400_v2.0.a1 AP2 − 0 0 94.9770681 ppe-miR319a 164.3097348 29.9171066 171.02544 Prupe.3G124900_v2.0.a1 TCP4 − 37.6537596 0 63.3180454 Prupe.4G083500_v2.0.a1 TCP2 − 75.3075192 80.39877794 664.8394767 Prupe.4G171100_v2.0.a1 PCF5 − 112.9612788 0 94.9770681 ppe-miR396a 6364.263728 59399.2645 71602.65089 Prupe.1G531300_v2.0.a1 AGL24 D-3-phosphoglycerate dehydrogenase 0 93.79857426 164.626918 GO:0045597 Positive regulation of cell differentiation ppe-miR166a 18,358.87437 7,847.487193 3,273.915566 Prupe.1G447100_v2.0.a1 ATHB-8 − 414.1913556 2,170.767004 2,421.915237 GO:0051301 Cell division ppe-miR160a 1,084.44425 333.6908044 177.1334915 Prupe.2G213000_v2.0.a1 ARF18 Auxin response factor 3 4,104.259796 9,286.058852 11,365.58915 Prupe.6G102800_v2.0.a1 ARF18 Hypothetical protein 78,884.62636 112,477.8903 81,142.07518 Prupe.6G236200_v2.0.a1 ARF18 Auxin response factor 3 10,806.629 33,566.48979 26,055.37568 ppe-miR171b 120.4938055 23.01315892 30.54025715 Prupe.2G138400_v2.0.a1 SCL6 DELLA protein GAI 1 37.6537596 0 0 ppe-miR171h 635.3309746 211.7210621 24.43220572 Prupe.2G138400_v2.0.a1 SCL6 DELLA protein GAI 1 37.6537596 0 0 GO:0008284 Positive regulation of cell proliferation ppe-miR166a 18,358.87437 7,847.487193 3,273.915566 Prupe.1G447100_v2.0.a1 ATHB-8 − 414.1913556 2,170.767004 2,421.915237 GO:0008285 Negative regulation of cell proliferation ppe-miR396a 6,364.263728 59,399.2645 71,602.65089 Prupe.1G419000_v2.0.a1 GRF6 − 1,242.574067 6,070.107734 4,558.899269 ppe-miR396b 61,791.41427 484,650.223 540,299.9053 Prupe.1G419000_v2.0.a1 GRF6 − 1,242.574067 6,070.107734 4,558.899269 ppe-miR396c 12,421.81595 118,018.3829 109,040.9341 Prupe.1G419000_v2.0.a1 GRF6 − 1,242.574067 6,070.107734 4,558.899269 Validation of identified miRNAs and their targets in peach
-
To validate the deep sequencing data, we employed the RLM-5'RACE approach to confirm four targets of ppe-miR160a and two targets of ppe-miR393a/b. Additionally, the expression levels of five miRNAs and their corresponding target genes were analyzed using qRT-PCR. For this validation, we selected five conserved miRNAs (ppe-miR159b, ppe-miR160a, ppe-miR167a, ppe-miR393a, and ppe-miR393b) and their target mRNAs, including auxin response factors, TIR proteins involved in auxin signaling, and MYB domain proteins. As shown in Fig. 6, five miRNAs showed differential expression patterns during fruit development. The expression levels of ppe-miR160a and ppe-miR159b gradually decreased throughout fruit development, while ppe-miR167a, ppe-miR393a, and ppe-miR393b showed an initial decline from DPA20 to DPA50, followed by an increase from DPA50 to DPA75 (Fig. 6). These expression patterns align with our high-throughput sequencing data (Table 1; Supplementary Table S3) and are consistent with previous studies highlighting the critical roles of ppe-miR160, ppe-miR167, and ppe-miR393 in auxin signaling[44].
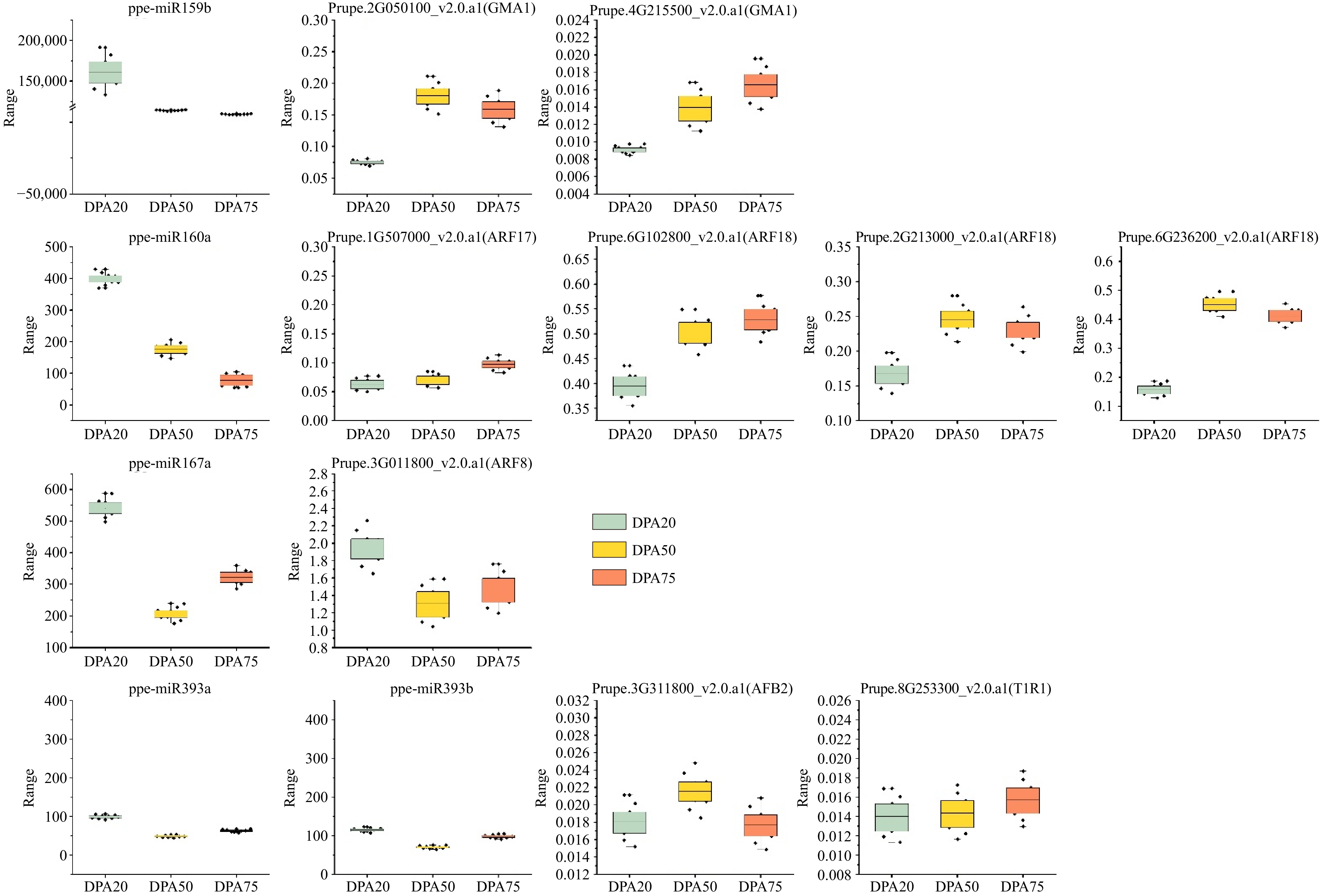
Figure 6.
Expression patterns of the four miRNAs identified and their target genes during fruit development and ripening; U6 and RPII were used as reference genes for miRNA and target genes, respectively.
To further confirm the regulatory roles of these miRNAs, we examined the expression patterns of nine target genes corresponding to the five miRNAs using qRT-PCR at the same developmental stages (Fig. 6). As expected, the expression patterns of the miRNAs and their target genes exhibited an inverse relationship. This finding was supported by degradome sequencing data, which revealed abundant cleavage products for these target genes (Supplementary Table S5). Additionally, RLM-5'RACE results confirmed the same cleavage sites for four targets of ppe-miR160a and two targets of ppe-miR393b (Supplementary Fig. S5, Supplementary Tables S7 & S8), further validating the reliability of the degradome sequencing data.
-
Numerous studies have highlighted the critical roles of miRNAs in fruit development[10,11,18,22,23]. Peach fruits are well-known to exhibit a double-sigmoid growth pattern, characterized by three distinct stages: FWI, FWII, and FWIII. While Zhu et al.[18] identified miRNAs in peach fruits using a mixed fruit sample, their analysis was limited to a single mixed library, leaving the dynamic regulation of miRNAs across sequential developmental stages largely unexplored. In this study, we conducted a comprehensive analysis of peach miRNAs and their targets across three key developmental stages, representing the double-sigmoid growth curve, through small RNA (sRNA) and degradome sequencing. Integrative analyses revealed that miRNA-mediated regulatory networks play a pivotal role in peach growth and development. However, the lack of biological replicates in our study presents a limitation in distinguishing true biological variations from technical noise. Future studies incorporating multiple replicates are essential to validate our findings and provide a more robust understanding of miRNA expression dynamics during peach fruit development.
MiRNAs during peach fruit development
-
Numerous fruit development-associated miRNAs have been identified in fruit-producing plants, including grape[16,17], apple[15], citrus[22], and peach[18]. In this study, 124 known miRNAs were detected across three small RNA libraries, with 85 miRNAs present in all three libraries. Previous studies have shown clear functional dose effects of miRNAs and suggested that highly and/or specifically expressed miRNAs are more likely to play versatile biological roles than with low abundance[47,48]. For example, in Hami melon, the miR159 and miR396 families exhibited high redundancy in their respective small RNA libraries[49]. Similarly, in our datasets, ppe-miR396b accounted for the highest proportion of normalized reads across all three libraries, representing approximately 50% of the known miRNA reads. In contrast, ppe-miR159a accounted for about 21% of conserved reads (Table 2), suggesting that both ppe-miR396b and ppe-miR159a play critical roles during peach development. Interestingly, in a mixed peach library from 'Troubadour', the ppe-miR156 family was the most abundantly expressed. However, in our study, ppe-miR156 was detected only in the DPA50 and DPA75 libraries, with no reads detected in the DPA20 library. This discrepancy suggests that single miRNAs may have distinct roles in different cultivars or developmental stages. Additionally, we identified 86 novel miRNAs, 79 of which were detected across all three libraries, while novel_193 was specific to the DPA50 library. The total number of novel miRNAs was lower than that of known miRNAs, and their absolute sequencing frequencies were considerably reduced, with most novel miRNAs exhibiting relatively low TPM values. This observation aligns with previous studies[50,51], which have shown that species-specific novel miRNAs often have low sequencing frequencies and exhibit more spatiotemporal expression patterns compared to conserved miRNAs.
Differences in miRNA expression profiles may reflect their regulatory roles in peach development. In this study, 97 and 79 miRNAs were differentially expressed in the DPA50 vs DPA20 and DPA75 vs DPA50 comparisons, respectively (Supplementary Table S4), indicating their involvement in cellular and developmental processes. Notably, some miRNAs were detected exclusively in one of the three developmental stages (Supplementary Table S4), suggesting that specific miRNAs may exhibit unique expression patterns during certain stages, enabling them to perform diverse regulatory functions in peach fruit development.
Target gene analysis of miRNAs in peach fruit
-
It has been proposed that plant miRNAs mediate gene expression mainly via miRNA-guided cleavage of target transcripts[51]. Therefore, identifying miRNA targets is a critical step in understanding the regulatory roles of miRNAs. In this study, degradome sequencing identified 216 cleaved targets for 74 known and eight novel miRNAs across the three developmental stages (Supplementary Table S5). Notably, the degradation levels of these targets varied across the three stages, suggesting that miRNAs may play distinct roles at different phases of peach fruit development. For example, the target genes of ppe-miR172a-3p, which encode RAP2-7 and AP2, showed an increasing abundance of degradome tags as fruit development progressed. This indicates that RAP2-7 and AP2 are actively targeted by ppe-miR172a-3p during fruit development and ripening. Many conserved miRNA targets, particularly transcription factors, regulate diverse physiological processes, and genetic programs[47−50]. In this study, the majority of targets for peach miRNAs were also conserved. For instance, the miR156 family is known to target SQUAMOSA promoter binding protein-like (SPL) transcription factors[42,44]. In the present experiments, six SPL transcription factors genes were identified as targets of ppe-miR156a/c/f. Similarly, the miR166 family has highly conserved targets encoding class III homeodomain leucine zipper (HD-ZIP III) transcription factors, which are involved in various developmental processes across plant species[52−54]. We predicted that four HD-ZIP III family members are the targets of ppe-miR166a and except ATHB-14, the cleavage products of REV, ATHB-8, and ATHB-15 are present in much greater quantities during FWII and FWIII than during FWI. This finding confirms that ppe-miR166a promotes late-stage development of fruits and may contribute to the fine-tuning of HD-ZIP III proteins. In addition to conserved targets, some miRNAs were found to have novel or additional targets in peach. For example, the miR396 family, known to target growth-regulating factor (GRF) genes, also targeted D-3-phosphoglycerate dehydrogenase, animal HSPA9 nucleotide-binding domain protein, and hypothetical proteins such as DAP and WEB1. Furthermore, 21 targets were detected for the eight novel miRNAs, many of which are involved in diverse biological processes. Specifically, 17 targets were identified for ppe-miR396a, including nine GRF genes, DNA topoisomerase 3-alpha, and other proteins. These findings suggest that miRNA-target interactions may exhibit species-specific characteristics during peach development.
Roles of miRNAs in peach fruit development
-
Auxin has long been considered a central regulator of ovary wall enlargement and pericarp development, and synthetic auxins have been shown to enhance fruit growth by stimulating fruit cell enlargement in peaches[54]. miRNAs have been shown to directly and indirectly regulate the auxin signaling pathway. For instance, the miR160 family targets auxin response factors (ARF10, ARF16, and ARF17) in multiple plant species[41,45,55], while the miR167 family regulates ARF6 and ARF8 to control reproductive processes[45,55]. Additionally, the miR393 family regulates the expression of the auxin receptor TIR1 and related auxin signaling F-box proteins AFB1, AFB2, and AFB3 in Arabidopsis and peaches[56]. In peaches, ppe-miR160a targets ARF17 to regulate peach development during the hard-core stage[44], and the miR164 family negatively regulates a subset of NAC transcription factor family members, which are auxin-responsive and the key regulators of plant growth and development[57]. Our sequencing data show that ppe-miR160a, ppe-miR167a, ppe-miR393a/b, and ppe-miR164a/d regulate auxin signaling by targeting ARFs, auxin receptor TIR1, auxin signaling F-box proteins (AFB2), and NAC transcription factors (Fig. 7; Supplementary Table S5). Specifically, ARF17 (Prupe.1G507000_v2.0.a1), ARF18 (Prupe.6G102800_v2.0.a1, Prupe. 2G213000_v2.0.a1, and Prupe.6G236200_v2.0.a1), and ARF8 (Prupe.3G011800_v2.0.a1) were identified as targets gene of ppe-miR160a and ppe-miR167a, with cleavage products varying across developmental stages. These indicates that the targets of single miRNAs play different roles in peach developmental stages. Both ppe-miR393a and ppe-miR393b target the TIR1/AFB2 Auxin Receptor Clade (Table 2), as observed in Arabidopsis[56], but ppe-miR393b exhibited significantly stronger degradation effects on TIR1/AFB2 than ppe-miR393a across all stages. Furthermore, NAC021, a target of ppe-miR164a, was implicated in auxin signaling, with its cleavage products more abundant during FWII and FWIII than FWI. These findings collectively demonstrate that miRNAs regulate peach fruit development by modulating auxin signaling.
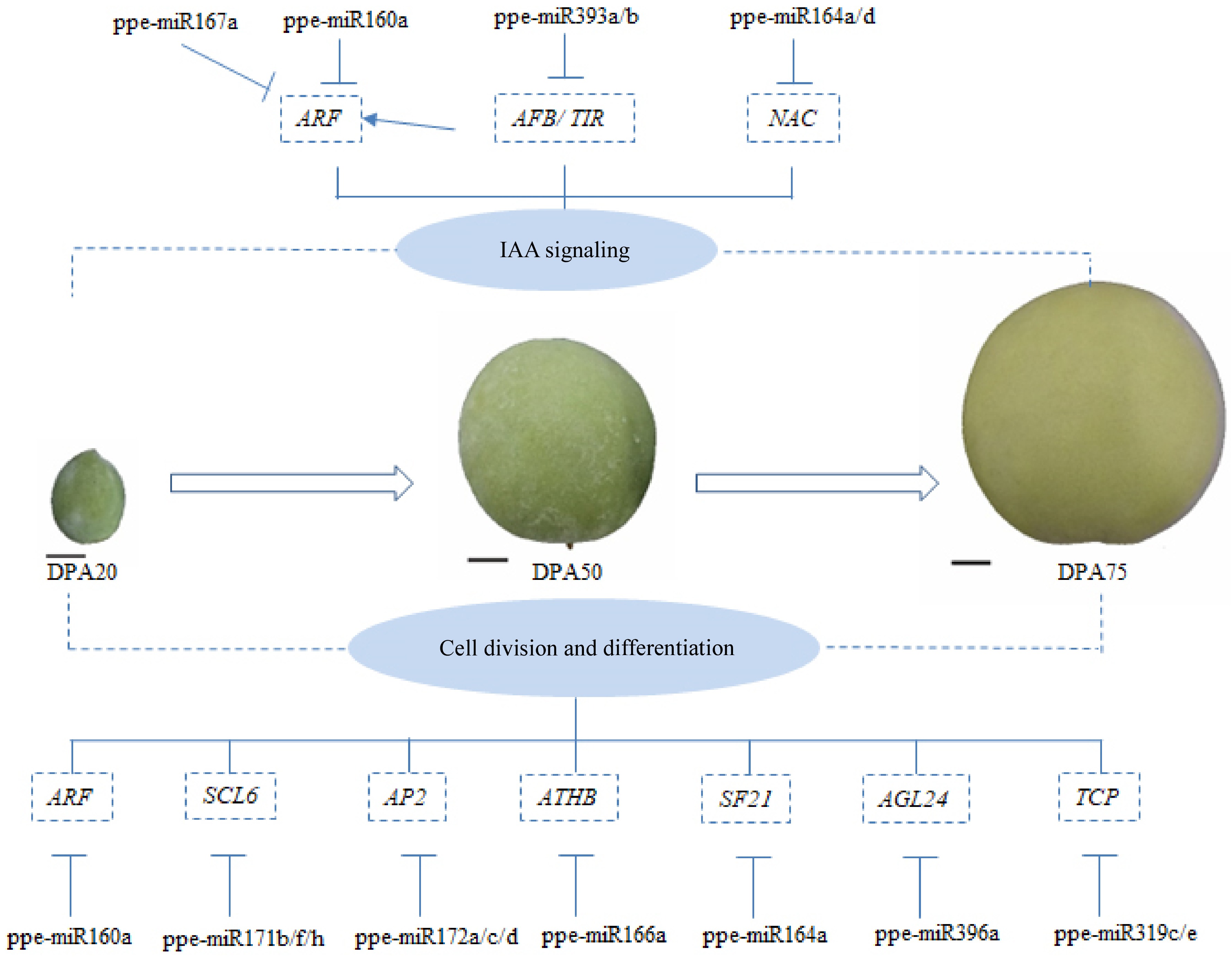
Figure 7.
A schematic model of the proposed roles of miRNAs in the early development of peaches; T bars and arrows refer to negative and positive effects on downstream effectors or biological process, respectively; scale bar = 1 cm
Fruit enlargement, a critical developmental process in peach and other fruiting plants, requires precise control of cell proliferation, expansion, and differentiation. To better understand the roles of miRNAs in peach enlargement, we analyzed miRNAs involved in cell proliferation and differentiation (Fig. 7; Table 2). For example, ppe-miR319c/e targets TCPs, which promote cell expansion and repress cell proliferation[58]. In our study, TCP2 cleavage products were more abundant during FWIII than FWI and FWII (Supplementary Table S5). Additionally, ppe-miR396a targets GRFs were detected, and these have been previously shown to regulate the growth of citrus fruits and leaves[59]. We also confirmed conserved interactions, such as ppe-miR172 a-3p/c/d targeting AP2 and ppe-miR166a targeting HD-ZIP III genes (ATHB8, ATHB14, ATHB15, and REV). In agreement, sl-miR172 targeting SlAP2a negatively regulated ethylene production during tomato ripening[60], and ppe-miR166a targeting the four HD-ZIP III subfamily genes ATHB8, ATHB14, ATHB15, and REV reportedly influences peach enlargement[44]. Notably, ppe-miR171b/f/h targeting SCL6 was implicated in cell division, a novel finding not previously reported. The MADS-box protein, encoded by AGL24 reportedly promotes flowering in Arabidopsis and was targeted by ppe-miR396a, which is also involved in cell differentiation (Fig. 7). Finally, ppe-miR160-ARF interactions, previously linked to fruit enlargement through auxin signaling, were also observed[44].
-
Using high-throughput RNA sequencing, we identified 124 conserved and 86 novel miRNAs in peach. Moreover, 216 sliced targets for 74 known and eight novel miRNAs were identified by degradome sequencing. Subsequent miRNA–gene–GO term network analyses highlighted a complex miRNA regulatory network involved in auxin signaling, which plays a central role in peach enlargement (Fig. 5). Collectively, these findings provide a foundation for future studies on miRNA-mediated mechanisms underlying peach development.
This work was supported by the Natural Science Foundation of Jiangsu (BK20201176 and BK20190542), Jiangsu University 'Qinglan Project' (2022). We thank Professor Chen Wang for her support and cooperation throughout this research.
-
The authors confirm contribution to the paper as follows: experiments conception and design: Zhang Y; experiments performing: Zhu X; data analysis: Wang C; manuscript writing: Zhang Y, Ge M. All authors reviewed the results and approved the final version of the manuscript.
-
This transcriptome project has been deposited in the NCBI SRA (Sequence Read Archive) database (www.ncbi.nlm.nih.gov/sra). The BioProject ID related to this paper is SRP143921 (www.ncbi.nlm.nih.gov/sra/?term=SRP143921).
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Statistical analysis of the sequencing reads from peach sRNA and degradome libraries; DPA, days post-anthesis.
- Supplementary Table S2 Characteristics of novel peach miRNAs from DPA20, DAP50 and DPA75 libraries.
- Supplementary Table S3 Differentially expressed peach miRNAs in DPA50 vs. DPA20 and DPA75 vs. DPA50 libraries; |log2(fold change)| > 1 and q-value < 0.01.
- Supplementary Table S4 Summary of the ppe-miRNAs targets identified by degradome sequencing at DPA 20, DPA50 and DPA75.
- Supplementary Table S5 Enrichment of differentially expressed target genes in DPA50 vs. DPA20 and DPA75 vs. DPA50 libraries; Threshold p-value ≤ 0.05.
- Supplementary Table S6 Details of the ppe-miRNAs targets identified by degradome sequencing at DPA 20 and DPA50.
- Supplementary Table S7 Details of the ppe-miRNAs targets identified by degradome sequencing at DPA 50 and DPA75.
- Supplementary Table S8 Primers used for qRT-PCR analyses and RLM-5'RACE.
- Supplementary Fig. S1 Changes in the longitudinal and transverse diameters of 'Baifeng' peach during different developmental stages.
- Supplementary Fig. S2 Proportion of the first nucleotide bias within novel miRNAs of the same length in the three libraries of peach sRNAs
- Supplementary Fig. S3 Expression analysis of the known and novel peach miRNAs identified by sequencing analyses; miRNAs were grouped into four classes (I–IV) based on their accumulation patterns. DPA20, DPA50 and DPA75 indicate 20, 50 and 75 days post anthesis, respectively.
- Supplementary Fig. S4 Significantly enriched GO terms for the target genes of peach miRNAs in DPA50 vs. DPA20 (a) and DPA75 vs. DPA50 (b) libraries; enriched factors were calculated according to the numbers of differentially expressed genes which were mapped to GO term divided by the numbers of all genes in that GO term; the top 20 enriched GO terms are listed.
- Supplementary Fig. S5 Target plot (t-plots) of peach miRNA targets and validation of target genes by RLM-5'RACE. The red line represents the sliced target transcripts. miRNA: mRNA alignments along with the miRNA-directed cleavage positions are shown below. Watson-Crick pairing (vertical dashes) and G:U wobble pairing (circles) are indicated and the numbers show the frequency of clones sequenced.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Zhu X, Wang C, Ge M. 2025. Characterisation of early fruit development-specific miRNAs and their targets in peach using small RNA and degradome sequencing. Fruit Research 5: e021 doi: 10.48130/frures-0025-0010 

Characterisation of early fruit development-specific miRNAs and their targets in peach using small RNA and degradome sequencing
- Received: 15 March 2023
- Revised: 28 February 2025
- Accepted: 05 March 2025
- Published online: 26 May 2025
Abstract: Peach (Prunus persica L.) is a key drupe fruit crop characterized by a double-sigmoid growth pattern during fruit development. MicroRNAs (miRNAs) play pivotal regulatory roles in this process; however, their functions in early peach fruit development remain poorly understood. To address this, we conducted high-throughput small RNA (sRNA) sequencing at three critical developmental stages: 20, 50, and 75 d post-anthesis (DPA20, DPA50, DPA75), representing exponential growth, lag phase, and rapid regrowth, respectively. We identified 124 known miRNAs from 70 families and 86 novel miRNAs, with 97 and 79 miRNAs differentially expressed between DPA50 vs DPA20 and DPA75 vs DPA50, respectively. Degradome sequencing revealed 216 target genes for 74 known and eight novel miRNAs, with targets of ppe-miR160a and ppe-miR393b validated using RNA ligase-mediated rapid amplification of 5' cDNA ends (RLM-5'RACE). Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) confirmed the expression patterns of five miRNAs and their target genes, consistent with sequencing data. Network analysis revealed the involvement of the auxin signaling pathway and a complex miRNA regulatory network in fruit enlargement. Key miRNA-target interactions, such as ppe-miR160a-ARFs, ppe-miR172a/c/d-AP2s, ppe-miR166a -ATHBs, and ppe-miR319c/e-TCPs were implicated in cell proliferation and differentiation, with novel findings including ppe-miR171b/f/h-SCL6s in cell division. In summary, peach miRNAs primarily regulate early fruit growth and development by modulating auxin signaling and mediating cell division and differentiation. These findings provide critical insights into miRNA-mediated mechanisms underlying peach fruit development and establish a foundation for future research on drupe fruit crops.
-
Key words:
- MiRNA /
- Fruit /
- Early development /
- Targets /
- Peach