










-
The forkhead box O (FoxO) subfamily is a core component of the forkhead transcription factor family, consisting of evolutionarily conserved members in mammals. This subfamily is multifunctional, participating in various critical cellular processes, for instance, endothelial cell proliferation[1], intestinal stem cell differentiation[2], muscle stem cell macroautophagy/autophagy[3], and naïve pluripotency[4]. Additionally, FoxO proteins are strongly associated with biological processes underlying diseases and aging. FoxO proteins have emerged as drug targets for the treatment of various diseases and the delay of aging[5,6]. Furthermore, several pharmacological approaches for regulating FoxO proteins have been developed[7,8].
FoxO1 is one of the four major components of the FoxO subfamily. It is closely associated with oxidative stress, which impairs cellular functions by disrupting the intracellular redox balance[9−11]. As a well-known effector of oxidative stress, FoxO1 mediates the regulation of various cellular functions through its transcription factor activity[11−13]. In type II alveolar epithelial cells, oxidative stress stimulates the deacetylation and nuclear translocation of the FoxO1 protein, which binds to the ROCK1 promoter to activate its transcription, thereby inducing mitochondrial autophagy and inflammation, ultimately resulting in cell apoptosis[12]. In addition to protein-coding genes, the transcription factor FoxO1 also mediates the cellular response to oxidative stress by activating or suppressing the transcription of non-coding RNAs (ncRNAs), such as miR-424[14] and miR-148-3p[15]. A recent study revealed that high glucose-induced oxidative stress facilitates FoxO1 phosphorylation and mitochondrial entry, increasing its interaction with the D-loop region of mitochondrial DNA, thereby suppressing mitochondrial respiration and inducing apoptosis of retinal pigment epithelium cells[16].
Long ncRNA (lncRNA), a type of transcript with a length exceeding 200 nt but lacking protein-coding potential, has emerged as a major research focus in the field of ncRNA, following microRNA (miRNA). Although oxidative stress has been strongly associated with lncRNA transcription[17], limited research exists on how oxidative stress regulates lncRNA transcription through the transcription factor activity of its effector protein, FoxO1. In this study, we identified lnc2300 (lncRNA LOC102162300) as a new transcriptional target of the transcription factor FoxO1 and participates in FoxO1 mediated oxidative stress induction of sow granulosa cell (sGC) apoptosis.
-
Commercial sow ovaries were provided by Zhushun (Nanjing, China). The following were obtained: pGL3 vector from Promega (Madison, WI, USA), small interfering RNAs (siRNAs) from Generay (Shanghai, China) (Table 1), mediums and fetal bovine serum (FBS) from Gibco (Carlsbad, CA, USA), Lipofectamine 2000 and trypsin from Thermofisher Scientific (Rockford, Waltham, MA, USA), H2O2 from Sigma (St. Louis, MO, USA), TRIzol, HiScript II Q Select RT SuperMix, AceQ qPCR SYBR Green Master Mix and Annexin V-FITC/PI Apoptosis Detection Kit from Vazyme (Nanjing, China), protein A/G agarose beads from Bioworld Biotech (Nanjing, China), and anti-FoxO1 antibody and IgG from Sangon (Shanghai, China).
Table 1. siRNA used in this study.
Name Sequence (5 'to 3') FoxO1-siRNA Sense: GCGGCAAAGAUGGCUUCUATT Antisense: UAGAAGCCAUCUUUGCCGCTT lnc2300-siRNA Sense: GAAUGCAGCGACUAUUAAUTT Antisense: AUUAAUAGUCGCUGCAUUCTT Online tools
-
Online tools used include the NCBI database (
www.ncbi.nlm.nih.gov ) for obtaining promoter sequences and RPKM values, JASPAR (http://jaspar.genereg.net ) for detecting the transcription factor binding sites (TFBSs), Wei Sheng Xin (www.bioinformatics.com.cn ) for GO analysis and KEGG analysis, miRDB (https://mirdb.org ) for detecting the miRNA recognition elements (MREs), and RNAhybrid (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid ) for predicting minimal free energy (mfe).Plasmids
-
The reporter of the lnc2300 promoter was generated by Tsingke (Nanjing, China). Expression vectors pcDNA3.1-FoxO1 and pcDNA3.1-lnc2300 were previously prepared by our laboratory[18,19].
Follicles and cells
-
Fresh ovaries were alternately washed with saline containing gentamicin and 75% alcohol, follicles with a diameter of 3−5 mm were removed, and follicular fluid was extracted. The follicle fluid was centrifuged to collect sGCs, which were then incubated in DMEM medium. KGN cells for reporter assay were revived and incubated in RPMI 1640 medium. Both mediums were supplemented with 15% FBS, and both cell types were incubated in 5% CO2 at 37 °C. All experiments were approved by the Animal Ethics Committee of Nanjing Agriculture University, China.
Cell treatment
-
For transfection, cells were seeded into 12-well culture plates for 36 h (~80% confluency), and transfected with a Lipofectamine 2000 reagent according to the manufacturer's instructions. For H2O2 treatment, cells were exchanged into DMEM medium without FBS for 12 h, and exposed to H2O2 at a final concentration of 150 μM for 2 h.
Reporter assay
-
After co-treating with three plasmids (pcDNA3.1-FoxO1, promoter-reporter, and pRL-TK) for 24 h, KGN cells were collected. A Dual-Luciferase Reporter Assay System was employed for reporter assay. Firefly luciferase activity was normalized to Renilla luciferase activity.
ChIP assay
-
Chromatin immunoprecipitation (ChIP) was conducted as previously described[17]. Chromatin from sGCs was immunoprecipitated with protein A/G agarose beads and anti-FoxO1 antibody. A pair of specific primers (5'-AGGCAGTTGCTTCTTTGT-3' and 5'-AGGAAGGAAAAGAAAGAACA-3') were designed for determining enriched DNA in the lnc2300 promoter containing FoxO1-response element (FRE).
qRT-PCR
-
Quantitative real-time PCR (qRT-PCR) was conducted as previously described[18]. Trizol was employed for RNA isolation, and two kits from Vazyme were used for reverse transcription and quantification. GAPDH and RPLP0 served as controls, and primers are given in Table 2.
Table 2. Primers for qRT-PCR.
Genes Primer sequences (5' to 3') Tm (°C) Size (bp) lnc2300 F: GCAAATCACGGTCAACCTACGA 60 181 R: TCAGCCAAAGAGACACATCTGAAAT FoxO1 F: GCAAATCGAGTTACGGAGGC 60 170 R: AATGTCATTATGGGGAGGAGAGT BAX F: TTTCTGACGGCAACTTCAACTGGG 64 122 R: TGTCCAGCCCATGATGGTTCTGAT BCL2 F: TTCTTTGAGTTCGGTGGGG 57 195 R: CCAGGAGAAATCAAATAGAGGC GAPDH F: GGACTCATGACCACGGTCCAT 60 163 R: TCAGATCCACAACCGACACGT RPLP0 F: TGCCCAGGGAAGACAG 60 119 R: TCTGCTTGGAGCCCACATT Apoptosis assay
-
The apoptosis rate was determined using a flow cytometry assay with a PI/annexin V‐FITC staining according to the manufacturer's instructions. sGCs were sorted with a Fortessa flow cytometer (Becton Dickinson, Franklin Lakes, NJ, USA). Data was analyzed with FlowJo v7.6 software (TreeStar, Ashland, OR, USA).
Statistics
-
Values were shown as the mean ± standard error of three independent experiments, and statistically analysed with GraphPad Prism v9 software (San Diego, CA, USA). Significance between two groups was compared with a Student's t-test (two-tailed), and significance among three groups was compared with an one-way ANOVA.
-
To characterize the lnc2300 promoter, a 1,457-bp sequence was obtained from the Yorkshire pig through amplification and sequencing. Its identity with the reference genome sequence of Sus scrofa (Duroc pig) is 98.97%, with only 15 bases differing, such as g.-534A > C, g.-540A > T, and g.-1454C > A (Supplementary Fig. S1). The lnc2300 promoter sequence of the Yorkshire pig is rich in adenine and thymine, with an A + T content (56.28%, 820/1457) exceeding the G + C content (43.72%, 637/1457) (Fig. 1a). Interestingly, a TATA box was identified at −779/−775 nt in the lnc2300 promoter (Fig. 1b), indicating that it is a TATA promoter. However, no CpG island was found in the lnc2300 promoter.
A CAAT box was identified at −73/−69 nt in the lnc2300 promoter (Fig. 1b). In addition to classic cis-elements, recent evidence suggests that MREs in promoters play a critical role in activating gene transcription[20]. Notably, 324 MREs of seven miRNAs were detected in the lnc2300 promoter (Fig. 1c, Supplementary Table S1). Of them, two miRNAs, including miR-214 and miR-92a, were down-regulated during sow follicular atresia (Fig. 1d). RNAhybrid observed a strong potential interaction between miR-214 and two DNA regions from −109 to −105 nt and −735 to −728 nt (Fig. 1e, f), or miR-92a and a DNA region from −62 to −54 nt (Fig. 1g). Moreover, the sequences of miR-214, mi-92a were found to have high identity across humans, mouse, and pigs, respectively (Fig. 1h, i). Collectively, the lnc2300 promoter contains abundant cis-elements that may contribute to its transcription.
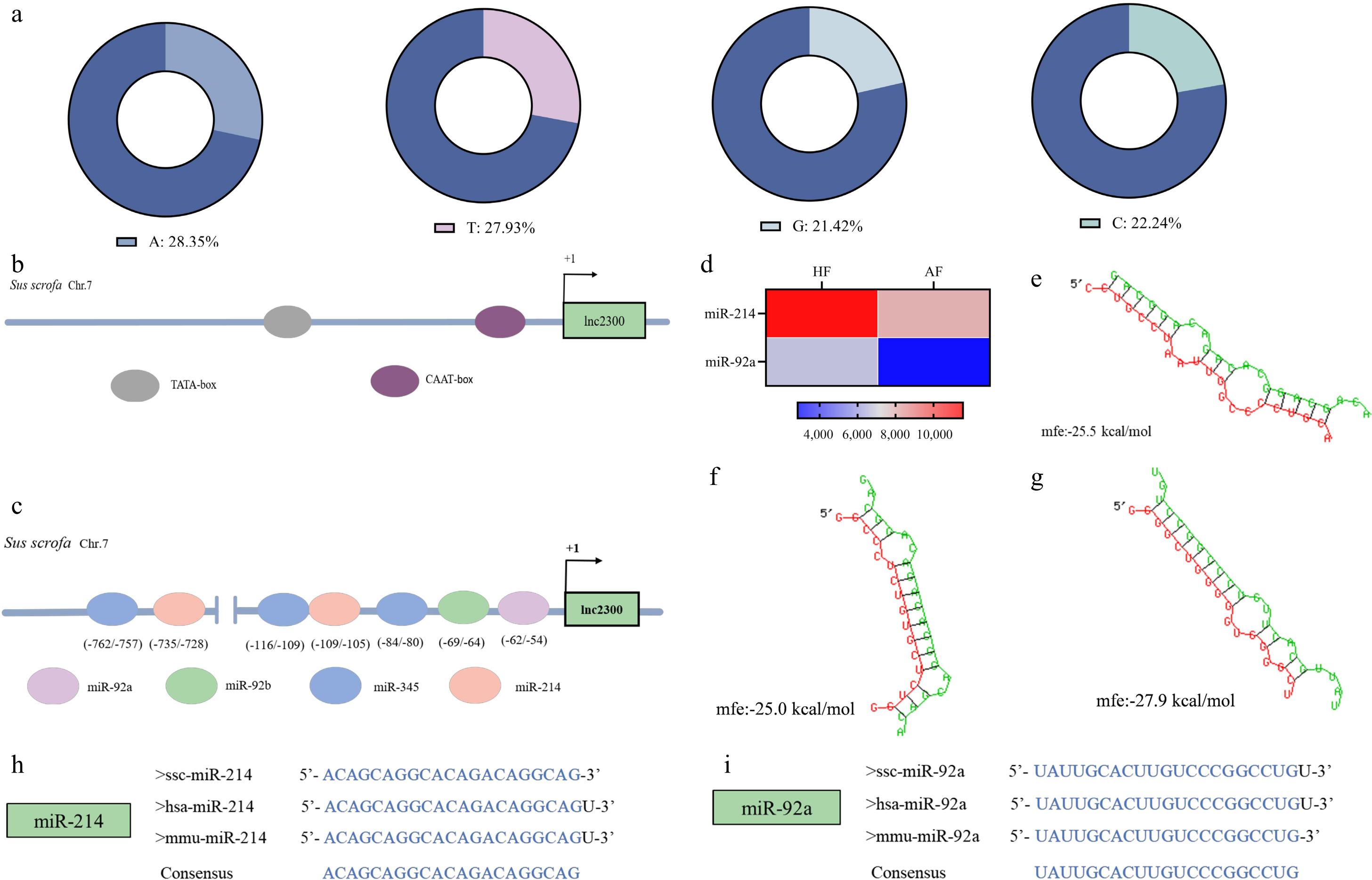
Figure 1.
Isolation and characterization of lnc2300 promoter. (a) Base composition of the lnc2300 promoter. (b) TATA-box and CAAT-box in the lnc2300 promoter are shown schematically. (c) miRNAs and MREs with strong binding capacity in the lnc2300 promoter are shown schematically. (d) Two miRNAs differentially expressed during follicular atresia. Chip data were obtained from a previous study[21]. The MRE motifs of (f) miR-214, (g) miR-92a, in the lnc2300 promoter were predicted by RNAhybrid. Alignment of mature sequences of (h) miR-214 and (i) miR-92a among humans, mouse, and pigs.
FoxO1 is a potential transcription factor of lnc2300
-
TFBSs are one of the most abundant cis-elements distributed on the promoter[22]. To understand the distribution of TFBSs in the lnc2300 promoter, we predicted potential TFBSs in this region by using an online tool, JASPAR, with a relative score of 90.0 as the threshold. A total of 2,075 putative TFBSs of 254 transcription factors were detected (Fig. 2a, Supplementary Table S2). In terms of molecular functions, the putative transcription factors associated with the lnc2300 promoter are involved in DNA-binding, transcription factor activity, sequence-specific DNA binding, and transcription regulator complex (Fig. 2b). Additionally, KEGG analysis revealed the enrichment of these transcription factors in pathways, including transcriptional misregulation in cancer, signaling pathways regulating pluripotency of stem cells and human T-cell leukemia virus 1 infection (Fig. 2c).
The electronic tissue expression profile revealed that among 254 transcription factors, 22 transcription factors, including NR1H2, NOBOX, and WT1, exhibited higher abundance in sow ovary (Fig. 2d). Interestingly, six transcription factors were differentially expressed in oxidative stress-stimulated sGCs, with five (e.g., FoxO1, NFIC, and FoxO3) being transcriptionally up-regulated, and 1 (NFIX) was transcriptionally down-regulated (Fig. 2e). Notably, as an effector of oxidative stress and a critical pro-apoptotic transcription factor in sGCs, FoxO1 exhibited the most significant fold change in sGCs with and without H2O2 treatment (Fig. 2e). Furthermore, we observed a strong negative correlation between FoxO1 levels and lnc2300 levels in follicles of sow ovary in vivo (Fig. 2f). Collectively, FoxO1 is a critical candidate transcription factor for regulating lnc2300 in the sow ovary.
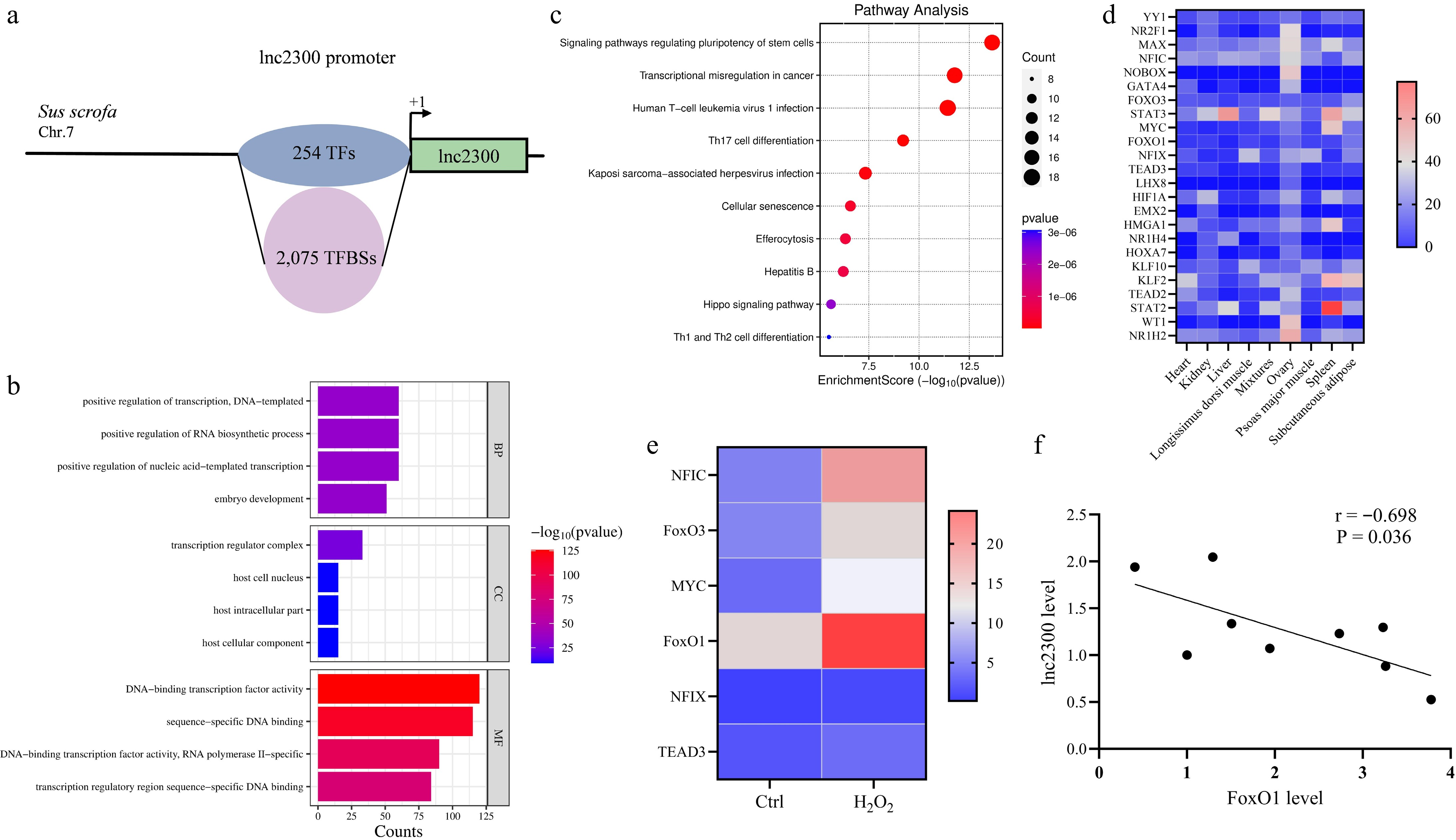
Figure 2.
FoxO1 is a potential transcription factor of lnc2300. (a) Schematic presentation of TFBSs in the lnc2300 promoter. GO and KEGG analysis of transcription factors that potentially bound to the lnc2300 promoter. (b) GO analysis. (c) KEGG analysis. (d) Heatmap indicating the high abundance of transcription factors in sow ovary. Data were obtained from NCBI database. (e) Heatmaps showing the expression of transcription factors with highly abundant in oxidative stress-stimulated sGCs. Data for RNA-seq were obtained from our previous study[23]. (f) Correlation analysis between FoxO1 and lnc2300 levels in sow antral follicles. n = 9.
FoxO1 binds to the lnc2300 promoter to suppress its transcriptional activity
-
A FRE motif (−270 TCCTGTTTACA −259) was identified in the lnc2300 promoter, with a relative score of 0.9 (Fig. 3a). To investigate whether transcription factor FoxO1 regulates the transcriptional activity of lnc2300, a reporter of promoter containing the FRE motif was prepared and co-treated with a FoxO1 expression vector in KGN cells cultured in vitro (Fig. 3b). The relative luciferase activity of this reporter decreased remarkably in response to overexpressed FoxO1 (Fig. 3c), suggesting that transcription factor FoxO1 suppresses lnc2300 transcriptional activity.
To further clarify whether transcription factor FoxO1 suppresses lnc2300 transcriptional activity through the FRE motif on its promoter, a reporter of promoter with a mutated FRE motif was prepared (Fig. 3b). The relative luciferase activity of this mutated reporter was no significant change in response to overexpressed FoxO1 (Fig. 3c), suggesting that transcription factor FoxO1 suppresses lnc2300 transcriptional activity via the FRE motif in its promoter. Furthermore, a direct interaction between the FoxO1 protein and the FRE motif of the lnc2300 promoter was observed in sGCs (Fig. 3d, e). Collectively, transcription factor FoxO1 binds to the lnc2300 promoter via the FRE motif, thereby suppressing its transcriptional activity.
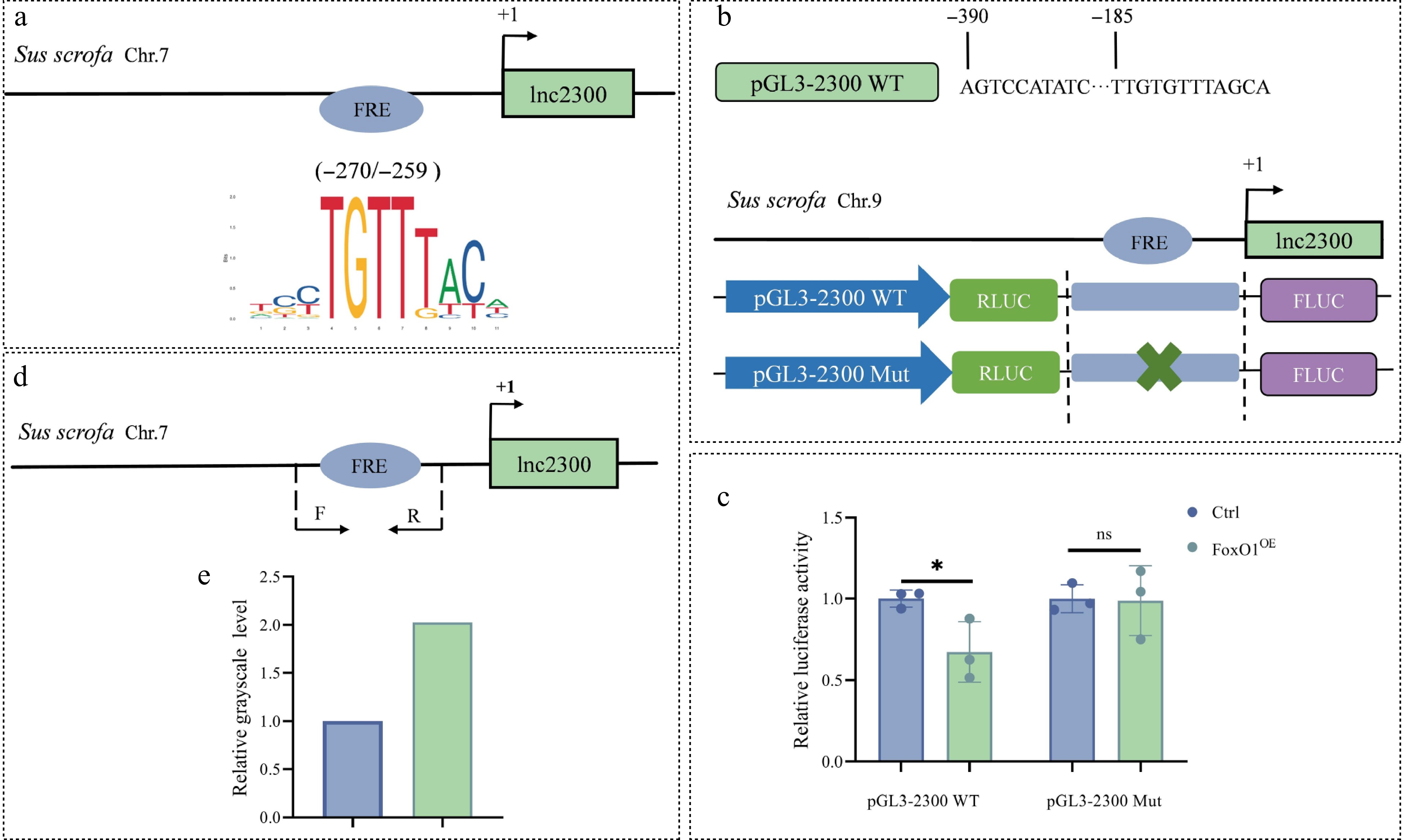
Figure 3.
FoxO1 binds to the lnc2300 promoter to suppress its transcriptional activity. (a) Schematic presentation of FRE motif in the lnc2300 promoter. FRE, FoxO1 response element. (b) Reporters of the lnc2300 promoter with FRE motif and its mutated version are shown schematically. (c) Reporter assay. pcDNA3.1-FoxO1 and reporters were co-transfected into KGN cells, luciferase activity was determined. (d) ChIP-PCR primers located in the lnc2300 promoter are shown schematically. (e) Enriched FoxO1 protein at the lnc2300 promoter was determined with ChIP. Data are expressed as mean ± SEM (n = 3). * p < 0.05. ns, no significance.
FoxO1 suppresses transcription and function of lnc2300 in sGCs
-
To investigate the regulatory effect of transcription factor FoxO1 on endogenous lnc2300 expression, we determined lnc2300 levels in FoxO1-overexpressing sGCs. As expected, lnc2300 levels significantly decreased in response to overexpressed FoxO1 (Fig. 4a). Conversely, lnc2300 levels significantly increased in FoxO1-silenced sGCs (Fig. 4b). These findings suggest that transcription factor FoxO1 suppresses lnc2300 transcription in sGCs.
FoxO1 and lnc2300 have been shown to play opposite roles in sow follicular atresia and sGC apoptosis[18,19]. However, whether FoxO1 induces sGC apoptosis via lnc2300 remains unclear. FACS analysis showed that FoxO1 significantly increases the proportion of apoptotic cells, while overexpression of lnc2300 alleviated this trend (Fig. 4c−f). Similarly, FoxO1 resulted in a remarkable decrease in the anti-apoptotic marker BCL2 levels in sGCs, while overexpression of lnc2300 rescued this decrease (Fig. 4g). In contrast, FoxO1 significantly increased pro-apoptotic marker BAX levels in sGCs, while overexpression of lnc2300 reversed this effect (Fig. 4h). Furthermore, FoxO1 significantly reduced the BCL2/BAX ratio, a hallmark indicator of cell apoptosis, while overexpression of lnc2300 rescued this decline (Fig. 4i). Collectively, FoxO1 induces sGC apoptosis by suppressing the lnc2300 transcript.
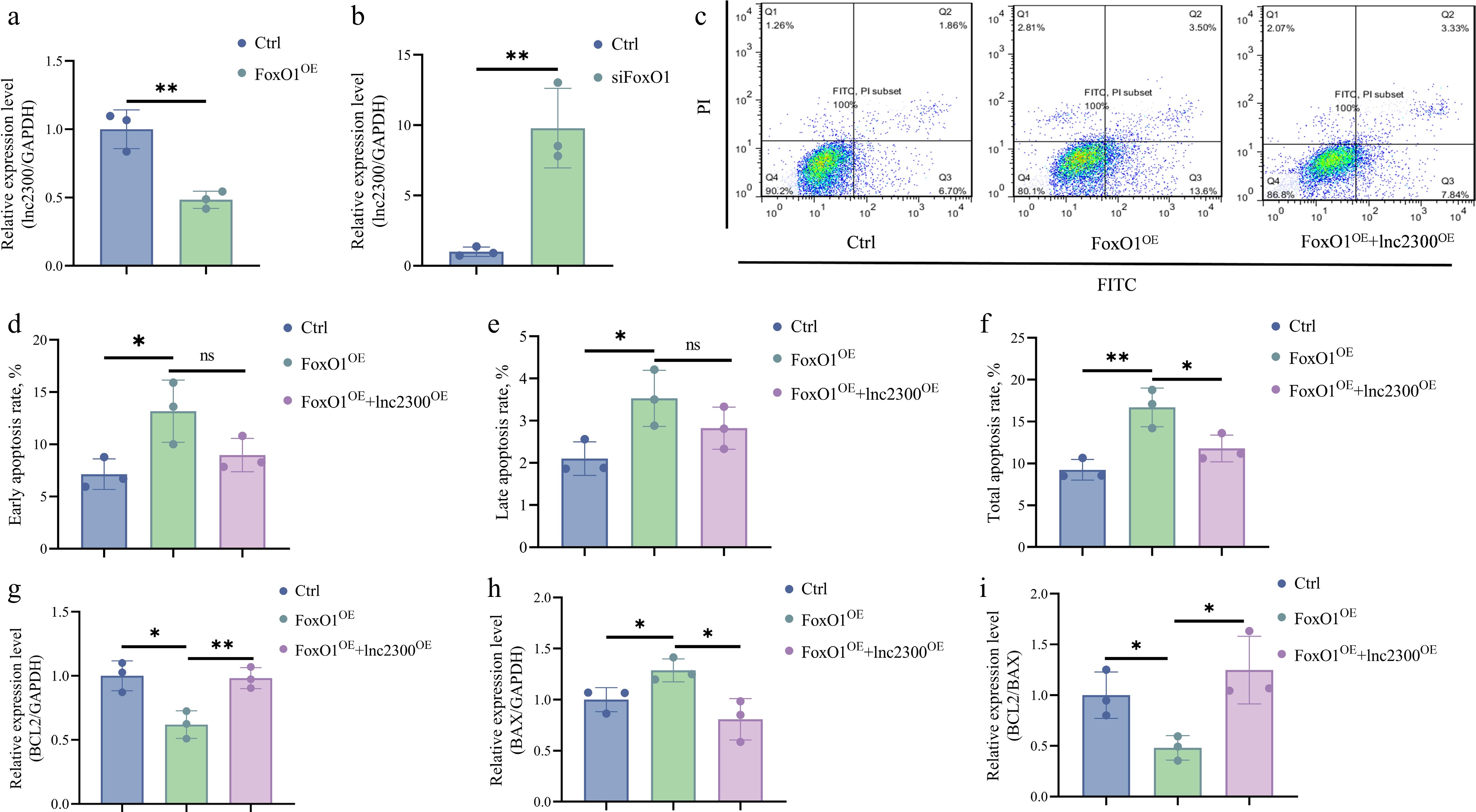
Figure 4.
FoxO1 suppresses transcription and function of lnc2300 in sGCs. sGCs were transfected with (a) pcDNA3.1-FoxO1 or (b) FoxO1-siRNA, lnc2300 levels were determined using qRT-PCR. (c)−(f) Apoptosis assay. sGCs were co-transfected with pcDNA3.1-FoxO1 and pcDNA3.1-lnc2300, apoptosis rate was determined by FACS. sGCs were co-transfected with pcDNA3.1-FoxO1 and pcDNA3.1-lnc2300, mRNA levels of (g) BCL2 and (h) BAX were determined, and the (i) BCL2/BAX ratio was analyzed. Data are expressed as mean ± SEM (n = 3). * p < 0.05. ** p < 0.01. ns, no significance.
FoxO1 mediates oxidative stress inhibition of lnc2300 transcription
-
As an effector of oxidative stress, FoxO1 mediates the regulation of its transcription targets in mammalian GCs[13]. Interestingly, we noticed a decrease in lnc2300 reads from our previous RNA-seq data for oxidative stress-stimulated sGCs (Fig. 5a). To verify this result, an oxidative stress model was established using sGCs exposed to H2O2. As expected, lnc2300 levels significantly decreased in response to oxidative stress in sGCs (Fig. 5b). Furthermore, silencing of FoxO1 rescued the decline in lnc2300 transcription caused by oxidative stress (Fig. 5c), suggesting that oxidative stress suppresses lnc2300 transcription in sGCs via FoxO1.
Next, we plan to analyze whether oxidative stress suppresses lnc2300 transcription depending on the transcription factor activity of FoxO1. A reporter assay demonstrated that lnc2300 promoter activity significantly decreased in response to oxidative stress, while silencing of FoxO1 rescued this decline (Fig. 5d), revealing that oxidative stress suppresses lnc2300 transcriptional activity in a FoxO1-dependent manner. Furthermore, oxidative stress increased the enrichment of transcription factor FoxO1 at the FRE motif of the lnc2300 promoter in sGCs (Fig. 5e, f). Collectively, oxidative stress suppresses lnc2300 transcription in sGCs in a FoxO1-dependent manner.
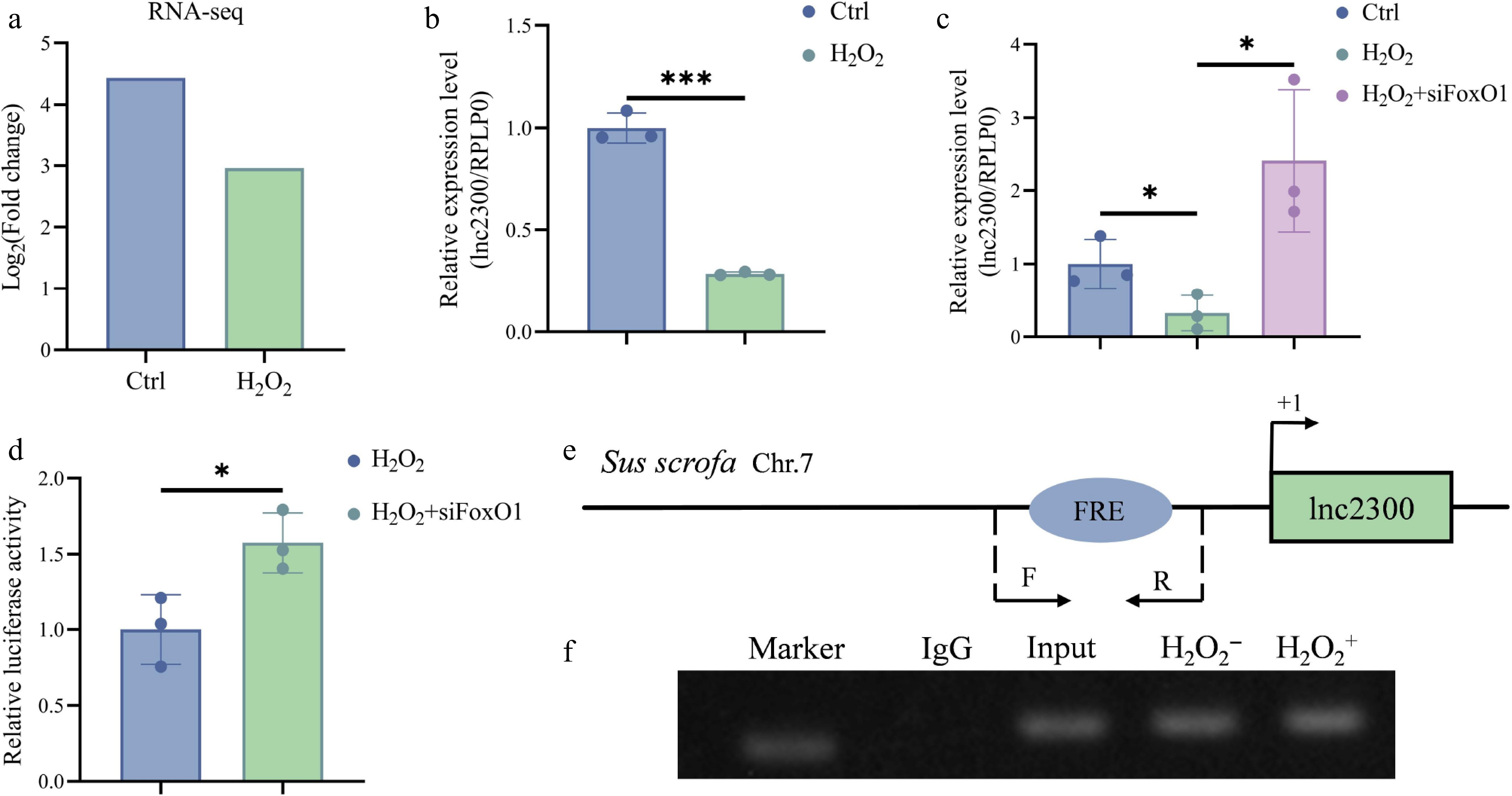
Figure 5.
FoxO1 mediates oxidative stress inhibition of lnc2300 transcription in sGCs. (a) lnc2300 was transcriptionally downregulated in oxidative stress-stimulated sGCs. Data was obtained from our previous RNA-seq[23]. (b) sGCs were exposed with H2O2, lnc2300 levels were determined with qRT-PCR. (c) sGCs were co-treated with H2O2 and FoxO1-siRNA, lnc2300 levels were determined with qRT-PCR. (d) sGCs were co-treated with reporter of the lnc2300 promoter, H2O2 and FoxO1-siRNA, luciferase activity was determined. (e) Specific primers are designed, to determine the (f) enriched FoxO1 protein on the lnc2300 promoter with ChIP assay, (f) in sGCs with or without H2O2 expose. Data are expressed as mean ± SEM (n = 3). * p < 0.05. *** p < 0.001.
lnc2300 mediates oxidative stress induction of sGC apoptosis
-
Excessive oxidative stress has been implicated in the induction of follicular atresia and GC apoptosis[24]. FACS analysis revealed that the proportion of apoptotic sGCs significantly increased in response to oxidative stress induced by H2O2 (Fig. 6a−d), indicating that oxidative stress acts as a stimulus for sGC apoptosis. Furthermore, overexpression of lnc2300 significantly alleviated the increase in apoptotic sGCs caused by oxidative stress (Fig. 6a−d), indicating that oxidative stress induces sGC apoptosis via lnc2300.
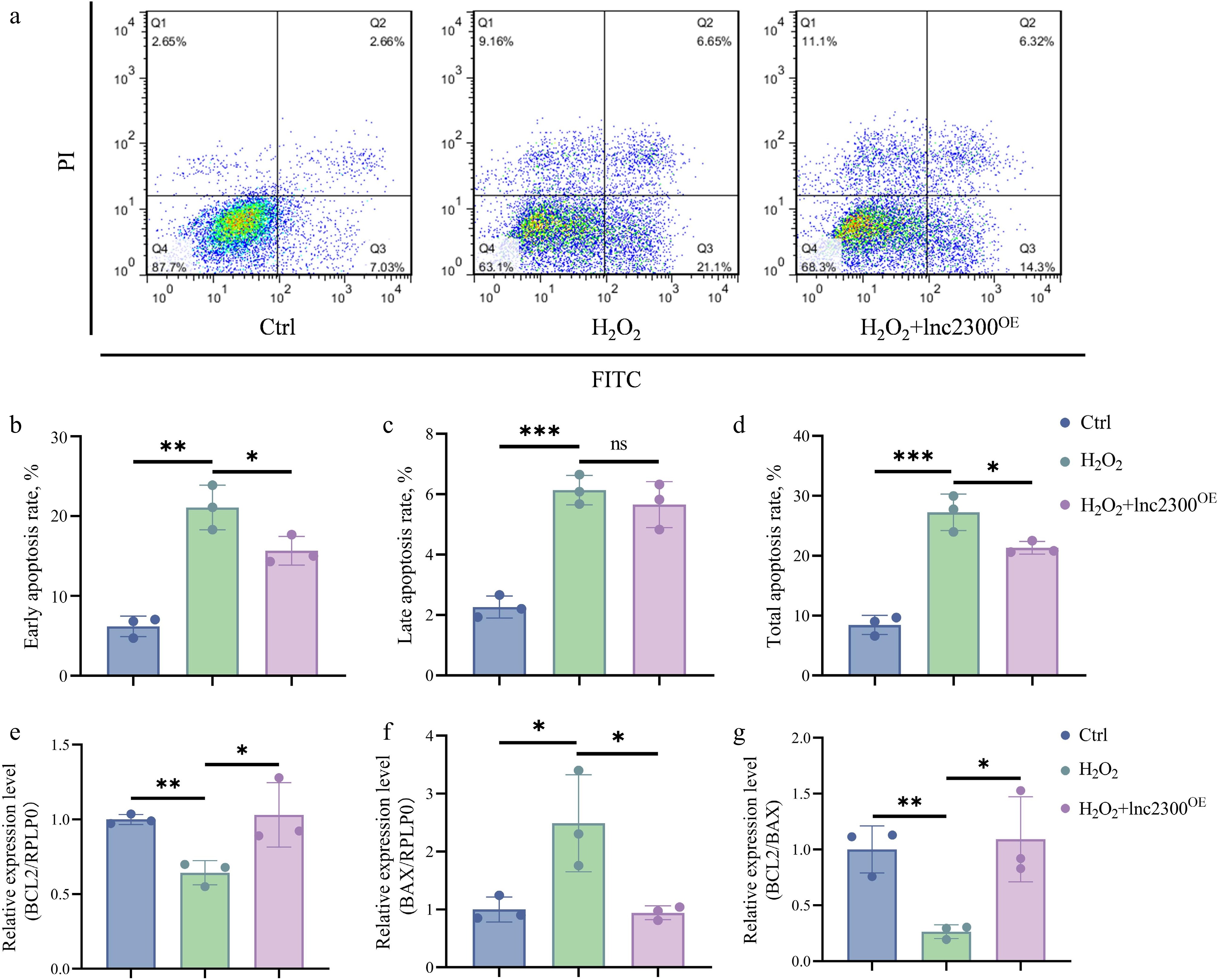
Figure 6.
lnc2300 mediates oxidative stress induction of sGC apoptosis. (a)−(d) Apoptosis assay. sGCs were co-treated with H2O2 and pcDNA3.1-lnc2300, apoptosis rate was determined by FACS. sGCs were co-treated with H2O2 and pcDNA3.1-lnc2300, mRNA levels of (e) BCL2 and (f) BAX were determined, and the (g) BCL2/BAX ratio was analyzed. Data are expressed as mean ± SEM (n = 3). * p < 0.05. ** p < 0.01. *** p < 0.001. ns, no significance.
To further validate the mediating role of lnc2300 on oxidative stress-induced sGC apoptosis, we also determined the levels of the anti-apoptotic marker BCL2 and the pro-apoptotic marker BAX in co-treating sGCs. As shown in Fig. 6e, oxidative stress significantly reduced BCL2 levels, while overexpression of lnc2300 rescued this decrease. By contrast, oxidative stress significantly increased BAX levels, while overexpression of lnc2300 reversed this effect (Fig. 6f). Furthermore, oxidative stress significantly reduced the BCL2/BAX ratio, while overexpression of lnc2300 rescued this decline (Fig. 6g). Collectively, oxidative stress induces cell apoptosis by suppressing the lnc2300 transcript in sGCs.
-
Promoters are critical for activating, repressing, or maintaining transcription of transcripts, as they contain abundant cis-elements, such as TFBSs for transcription factors and MRE motifs for miRNAs[20,22]. These transcripts include both mRNAs and ncRNAs. Lnc2300 is a recently identified lncRNA down-regulated during sow follicular atresia[19]. In this study, we identified the transcription factor FoxO1 as a novel transcription repressor of lnc2300 in sGCs. Interestingly, FoxO1 is up-regulated during sow follicular atresia[18], in contrast to lnc2300. Furthermore, FoxO1 can also act as a transcription factor to activate its transcription in mouse GCs[25]. Several differentially expressed genes during follicular atresia have been identified as transcriptional targets of FoxO1 in mammalian follicular cells, such as the CCND2 gene in rat GCs[26], and LHCGR in rodent GCs[27]. However, no ncRNA targets of transcription factor FoxO1 have been identified in the mammalian ovary, despite thousands of differentially expressed ncRNAs, including miRNAs[21], lncRNAs[18], and circRNAs[28], have been identified during sow follicular atresia. Collectively, our findings reveal the mechanism underlying the transcriptional downregulation of lnc2300 during sow follicular atresia, and identify a novel ncRNA target of FoxO1 in the mammalian ovary. In the future, additional targets of FoxO1, including mRNAs and ncRNAs, should be identified through high-throughput technologies such as ChIP-seq to comprehensively elucidate the role of transcription factor FoxO1 in follicular atresia. Ultimately, this would enable the assessment of therapeutic targets for rescuing follicular atresia and improving follicular utilization efficiency.
Oxidative stress is widely recognized as a key factor in sow follicular atresia, as it significantly increases oxidative stress levels and activates oxidative stress-related pathways in atretic follicles[28]. Oxidative stress also induces sGC apoptosis, which subsequently triggers follicular atresia[17]. Meanwhile, the levels of FoxO1, an effector of oxidative stress, are increased in sow atretic follicles[17,29]. In mammalian GCs, including sGCs, evidence suggests that oxidative stress stimulates FoxO1 expression and activity through multiple mechanisms, such as activating its transcription via transcription factors[25], controlling its expression at the post-transcriptional level via miRNAs[18], and maintenance of protein stability through deacetylation[29]. Notably, oxidative stress also enhances the nuclear translocation of the FoxO1 protein, thereby mediating transcription factors and regulating cellular functions in mammalian GCs, including sGCs[29,30]. However, limited studies have explored the involvement of lncRNAs in the oxidative stress-FoxO1 system in mammalian GCs. In this study, we showed that FoxO1 mediates the regulatory effect of oxidative stress on its transcriptional target lnc2300 transcript and sGC apoptosis. In human GCs, FoxO1 interacts with NCOA4 promoter to mediate oxidative stress induction of cell ferroptosis[13]. In rodent GCs, FoxO1 binds to the promoters of autophagy-related genes, including ATG5 and ATG12, to mediate oxidative stress induction of cell autophagic death and inhibition of cell viability[31]. Our findings provide new evidence that FoxO1 regulates sGC functions by controlling lncRNA transcription in a transcription factor activity-dependent manner. Combined with our previous study[18], a novel signaling pathway, FoxO1-lnc2300-CYP11A1, for oxidative stress-induced apoptosis in sGCs has been defined. Importantly, we demonstrated for the first time that lnc2300 exhibits antioxidant stress resistance in mammalian GCs, providing a potential non-hormonal target for future therapeutic strategies to mitigate female reproductive decline and treat ovarian diseases caused by oxidative stress, and lnc2300 is expected to be developed as an endogenous antioxidant for regulating sow reproduction.
-
In summary, this study describes a mechanism underlying the transcriptional downregulation of lnc2300 during sow follicular atresia. Lnc2300 was identified as a novel transcriptional target of FoxO1, and the signaling axis formed by these two mediates oxidative stress-induced apoptosis in sGCs (Fig. 7). Our findings propose a novel model for oxidative stress-induced GC apoptosis and suggest a potential non-hormonal antioxidant for regulating female reproduction.
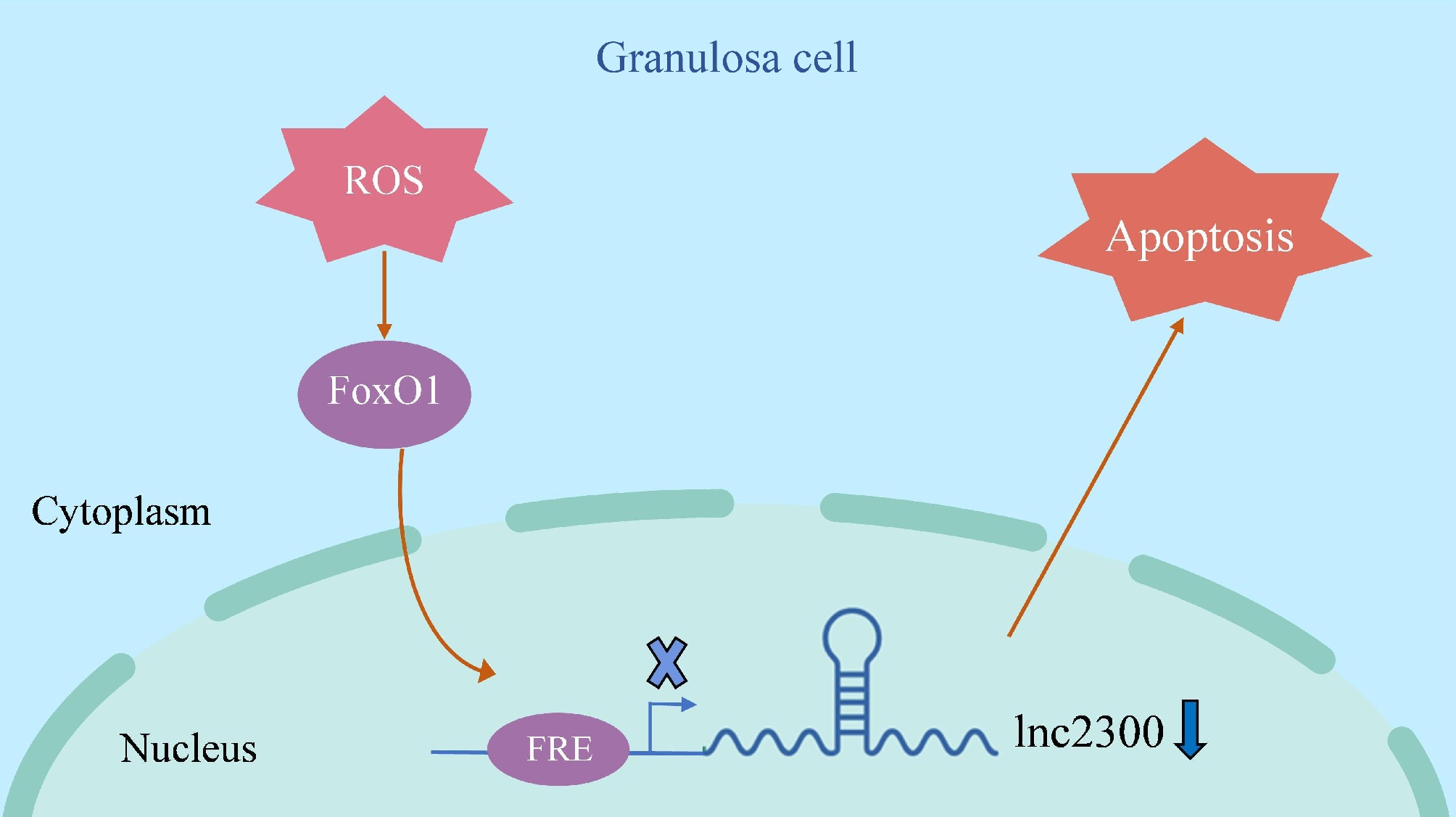
Figure 7.
Schematic illustration of lnc2300-mediated oxidative stress induction of sGCs. When sGCs undergo oxidative stress, its effector FoxO1 levels increase, which leads to transcription repressor FoxO1 enrichment in the promoter of transcriptional target lnc2300, thereby inhibiting lnc2300 transcription and causing cell apoptosis.
This work was supported by the National Key R&D Program of China (2022YFD1600903), the Fundamental Research Funds for the central universities (KYLH2025010), and Student Research Training for the Nanjing Agricultural University (2025).
-
All procedures were reviewed and preapproved by the Animal Ethics Committee of Nanjing Agricultural University (Identification No. 20223024059, approval date: Dec 2021.
-
The authors confirm contribution to the paper as follows: conception and design: Li Q, Li Y, Sheng W; collection and assembly of data: Sheng W, Xu Z; data analysis and interpretation: Sheng W, Fu Z, Zhang J, Du X; draft manuscript preparation: Sheng W, Li Q. All authors reviewed the results and approved the final version of the manuscript.
-
All data during this study are included in this published article.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 miRNAs and MREs ofthe lnc2300 promoter.
- Supplementary Table S2 The putative TFBSs in the lnc2300 promoter.
- Supplementary Fig. S1 Sequence amplification of the lnc2300 promoter region. lnc2300 promoter region identity with the reference genome sequence of Sus scrofa (Duroc pig).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Nanjing Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sheng W, Xu Z, Fu Z, Zhang J, Du X, et al. 2025. lnc2300 mediates the induction of granulosa cell apoptosis by oxidative stress. Animal Advances 2: e015 doi: 10.48130/animadv-0025-0013 

lnc2300 mediates the induction of granulosa cell apoptosis by oxidative stress
- Received: 10 December 2024
- Revised: 14 March 2025
- Accepted: 24 March 2025
- Published online: 04 June 2025
Abstract: lnc2300 (LOC102162300) is a newly characterized lncRNA that is down-regulated during sow follicular atresia and oxidative stress-stimulated sow granulosa cells (sGCs). However, the mechanism underlying its reduced abundance in atretic follicles and oxidative stress-stimulated sGCs remains unclear. Here, we reported that lnc2300 is transcriptionally suppressed by oxidative stress in a FoxO1-dependent manner. The lnc2300 promoter was characterized, and multiple cis-elements were identified, including the binding site of transcription factor FoxO1, an effector of oxidative stress. ChIP combined with reporter assays demonstrated that transcription factor FoxO1 directly binds to the lnc2300 promoter, thereby suppressing its activity. Furthermore, FoxO1 suppresses the lnc2300 transcript in sGCs by acting as a transcription repressor. Functionally, FoxO1 induces sGC apoptosis by suppressing the lnc2300 transcript. Additionally, FoxO1 mediated the promotion of oxidative stress on the lnc2300 transcript and its anti-apoptotic function in sGCs. Overall, these findings provide new insights into the mechanisms underlying the transcriptional regulation of lnc2300 and oxidative stress-induced sGC apoptosis.
-
Key words:
- lnc2300 /
- Granulosa cell apoptosis /
- FoxO1 /
- Oxidative stress