










-
In maize (Zea mays L.) seeds, starch, and storage proteins represent the primary components of the endosperm, which is a crucial tissue for nutrient storage and seed development. Starch constitutes the largest proportion, accounting for approximately 70%−75% of the seed dry weight, and serves as the main energy reserve for the germinating seedling. Storage proteins, primarily zeins, make up around 8%−10% of the dry weight and play an essential role in providing amino acids required for early growth. Together, starch and storage proteins support the metabolic needs of the seed during germination and early seedling development. The balance and accumulation of these components are tightly regulated through complex molecular pathways involving enzymes and transcription factors, which ensure that maize seeds have the necessary nutrients to sustain growth and development upon germination. Understanding the genetic and biochemical mechanisms behind starch and protein biosynthesis in the endosperm is critical for improving the yield and quality of maize seeds[1]. Seed texture and quality are determined by the ratio of vitreous to starchy endosperm. The vitreous endosperm, located in the outer layer of seeds, enhances resistance to pests and diseases and protects the grain from mechanical damage during harvest and transport[2]. The abundance of zein gene expression plays a crucial role in forming vitreous endosperm during seed maturation[3−5]. Zeins, which comprise over 60% of maize proteins, are classified into four major groups: α-zeins (19 and 22 kDa), β-zein (15 kDa), γ-zeins (16, 27, and 50 kDa), and δ-zeins (10 and 18 kDa) based on their amino acid sequences[6]. While α-zeins are encoded by a multigene family, other zeins are encoded by single-copy or low-copy genes[7,8]. The expression patterns and regulation of these genes are critical for zein accumulation in maize seeds.
Recent studies have demonstrated that specific transcription factors are central to regulating the biosynthesis of starch and storage proteins in the seed endosperm[9]. These transcription factors function as master regulators, orchestrating the expression of genes involved in the synthesis and accumulation of both starch and storage proteins, such as zeins. The coordinated activity of these factors ensures the proper balance between starch production, which provides the energy reserves for germination, and storage protein synthesis, which supplies essential amino acids. These transcription factors often respond to environmental cues, metabolic signals, and developmental signals, allowing the plant to adjust its seed composition to optimize seed viability and growth potential. For example, factors like WRI1, LEC1, and FUS3 have been implicated in the regulation of genes responsible for storage protein and starch synthesis[10]. By modulating the expression of key enzymes, these transcription factors help control the timing and extent of starch deposition and protein accumulation, which are critical for seed development and subsequent seedling growth. The intricate regulation of these processes by transcription factors is essential for improving crop yield and quality, offering potential targets for genetic engineering to enhance seed nutrient content[9]. The first zein regulator identified in the classic opaque endosperm mutants was the bZIP protein OPAQUE2 (O2), a central regulator of nearly all zein gene families[11−13]. O2 regulates zein gene expression through additive and synergistic interactions with multiple transcription factors (TFs), including O2-heterodimerizing proteins (OHPs), Prolamin-box binding factor 1 (PBF1), bZIP transcription factor 22 (bZIP22), and MADS-box transcription factor 47 (MADS47)[14−17]. ZmNAC128 and ZmNAC130, in coordination with O2, activate the transcription of γ-zein genes (50, 27, and 16 kDa) and starch metabolism genes, including Brittle2, pullulanase-type starch debranching enzyme, granule-bound starch synthase I, starch synthase I, starch synthase II, and sucrose synthase I, thereby regulating the accumulation of storage proteins and starch in maize seeds[18,19]. Additionally, Opaque11 (O11), a key TF in endosperm development and nutrient metabolism, directly regulates other key TFs involved in protein deposition, such as O2 and PBF1[20]. ZmABI19 also directly regulates several essential grain-filling TFs, including O2, O11, NAC130, ZmbZIP22, and PBF1, in the endosperm nutrient reservoir[21]. However, the signaling pathways that regulate these TFs and zein genes remain unclear.
Sucrose plays a dual role in plant growth and development, serving as both a vital source of energy and carbon and as a key signaling molecule that regulates various developmental and metabolic processes[22,23]. It has been shown to synergistically regulate starch synthesis, storage protein accumulation, and grain filling[24−27]. In maize, sucrose and ABA jointly regulate the starch biosynthesis-related gene ZmSSIIIa via the transcription factor ZmEREB156, driving starch accumulation in the endosperm[28]. Sucrose also promotes the expression of O2, a hub gene that coordinates the accumulation of storage reserves in the endosperm[29]. Similarly, in potato, sucrose induces the expression of the patatin gene, which encodes a storage protein constituting 30%−40% of the total soluble proteins[30]. However, whether sucrose signaling influences zein protein accumulation in maize endosperm remains unclear. In this study, we identified the gene underlying the ven2 mutant and confirmed it as a mutant allele of Mn1/Incw2, which encodes a cell wall invertase expressed in the basal endosperm[31]. Beyond its role in seed size regulation, we demonstrate that Mn1/Incw2 also affects seed texture and quality through sucrose metabolism.
-
The mutants ven2 and ven2-1 were identified in the ethyl methane sulfonate (EMS) mutagenesis populations of the inbred lines B73 and HuangZao4, respectively. The fine mapping population was developed using the F2 progenies of C733 × ven2. OE-Incw2 is a gift from Prof. Guangtang Pan (Sichuan Agricultural University, Sichuan, China), which is Incw2 gene driven by 27 kDa γ-zein promoter specifically over-expressed in the endosperm cells[32]. The ven2-2 mutant was in W22 inbred line background obtained from the Maize Genetic Cooperation Stock Center.
Measurement of starch and protein contents
-
Twelve mature seeds of wild type (WT) and mutants were selected for further research, respectively. The pericarp and embryo of seeds were removed and the endosperm were ground into fine flour. Then, 50 mg of each sample was used for starch and proteins measurements according to previous described[33]. SDS-PAGE gel with Coomassie Brilliant Blue (CBB)-staining was performed to analyze the accumulation patterns of zein proteins in WT, ven2, ven2-1, and OE-Incw2.
Scanning electron microscopy (SEM)
-
For scanning electron microscopy, mature WT and ven2 seeds were rifted longitudinally with a razor. Samples were critical-point dried, spray-coated with gold and then observed under a scanning electron microscopy (JSM-6610LV) at Shandong Agricultural University (Shandong, China).
Map-based cloning
-
To clone the causative gene for ven2 phenotype, the homozygous mutants were out-crossed with maize inbred line C733, and then self-pollinated the offspring to generate the F2 segregating population. A total of 1,547 homozygous ven2 mutant seeds from F2 segregating population with vitreous endosperm phenotype were used for gene mapping. Polymorphic molecular markers listed in Supplementary Table S1 were used to localize the Ven2 locus to a 280-kb region between the molecular markers 28z-27 and c-3799-2 containing four annotated genes. The corresponding DNA fragments of candidate genes were amplified from WT and ven2 seeds using KOD DNA polymerase (Toyobo, Japan) and sequenced. Finally, allelism tests were carried out among ven2, ven2-1, and ven2-2.
RNA extraction and reverse-transcription quantitative PCR (qRT-PCR)
-
Total RNA was extracted from plant tissues using the Ultrapure RNA Kit (cwbiotech, CW0581M). After DNase treatment to remove genomic DNA, 2-μg of total RNA per sample was subjected to synthesize first-strand cDNA using FastQuant RT Kit (TIANGEN) according to the instructions. Next, qRT-PCR was performed using SYBR Green qRT-PCR kit (TIANGEN) on a Light Cycler 96 (Roche Diagnostics) detection system. The gene expression was normalized to the maize Actin gene (Zm00001d010159) with three biological replicates. The primers used in qRT-PCR are listed in Supplementary Table S1.
Measurement of sucrose, glucose, and fructose
-
The developing seeds of WT, ven2, CK, and OE-Incw2 at 6, 12, 18, and 24 DAP were cut longitudinally transversely along the base of the embryo into two parts (upper and basal seeds section). Each sample was ground into a fine flour with liquid nitrogen. First, the total soluble sugar in the sample was extracted three times by 80% ethanol at 80 °C. And then, the resulting soluble sugar solution was dried into flour at 60−80 °C. The fructose, glucose, and sucrose were differentiated by HPLC systems (LC-20AT, Ultimate 3000) using XBridge® Amide Column (4.6 mm × 250 mm, 3.5 μm, Waters) at Shandong Analysis Test Center. The operating temperature was 25 °C, and mobile phases A and B were acetonitrile and H2O, respectively. And the flow rate was 0.8 mL/min.
Transient expression assays, exogenous sugar induction, and LUC activity assay in maize endosperm
-
Approximately 1,500 bp regions region of 19 kDa α-zein and 22 kDa α-zein were amplified with the primers listed in Supplementary Table S2. Then these two sequences were introduced into the pGreenII0800-LUC vector, respectively, yielding the construct pro19 kDa-LUC and pro22 kDa-LUC. Endosperms at 16 DAP were excised and sterilized with 75% (v/v) ethanol and cultivated on MS medium at 28 °C, with a 4-h dark photoperiod. Then the prepared endosperm was subject to bombardment using a helium biolistic gun transformation system (PDS-1000, Bio-Rad) according to the manufacturer's instructions and the methods previously described[34]. The bombarded endosperm is cultivated on MS medium supplied with 200 nM mannitol (as the control), 200 nM sucrose, 200 nM glucose, and 200 nM fructose, respectively, at 28 °C, for 24-h dark photoperiod to analyze LUC activity. For the LUC activity assay, both LUC and REN activities were measured using the Dual-Luciferase Reporter Assay System (Promega, USA) according to the manufacturer's instructions on the Enzyme label instrument (PerkinElmer EnSpire). Relative LUC activity was calculated by normalizing the LUC activity to the REN activity.
Transient expression assay in leaves of N. benthamiana
-
Approximately 1,500 bp regions of the 19 kDa and 22 kDa α-zein gene promoter were cloned into the vector pGreenII0800-LUC to generate a reporter. The full-length coding sequences of SRF1 (Zm00001d021947) were inserted into vector pGreenII62-SK under the control of the 35S promoter to generate the effectors. pGreenII62-SK under the control of the 35S promoter was used as the negative control effector. The reporter and the effector were introduced into Agrobacteriumtumefaciens GV3101 strain (pSoup-p19, Catalog No. AC1003, Shanghai Weidi Biotechnology Co., Ltd., Shanghai, China) and then were co-infiltrated into the leaves of N. benthamiana as described previously[35]. The LUC/REN activity ratio was measured using a Dual-luciferase Reporter Assay System (Promega).
Recombinant protein expression and purification
-
The full coding sequences of SRF1 were fused with GST in the expression vectors pGEX-4T-1, and expressed in E. coli BL21 (DE3) strain. The recombinant proteins were induced with 0.2 mM isopropyl-β-D-thiogalactoside (IPTG) for 16 h induction at 16 °C and were purified with glutathione beads from GE Healthcare. Primers are listed in Supplementary Table S2.
Electrophoretic mobility shift assay (EMSA)
-
Oligonucleotide probes of the 19 kDa and 22 kDa α-zein gene promoter was synthesized and labeled with biotin at the 5' end by Sangon Biotech. Biotin-labeled DNA was detected according to the instructions of the LightShift Chemiluminescent EMSA Kit (Thermo Fisher Scientific, Catalog No. 20158) according to previous studies[36,37].
-
A mutant was derived from an EMS-mutagenized B73 population. Homozygous mutants were crossed with B73 and C733 inbred lines, respectively. Phenotypic segregation in the F2 seeds followed a 3:1 ratio of wild-type (WT, +/+ and ven2/+) to mutant (ven2) phenotypes, confirming that it is a recessive, nuclear monogenic mutation (Supplementary Table S2). The mutant seeds were smaller, shrunken, and vitreous compared to WT seeds (Fig. 1a; Supplementary Fig. S1a, b), exhibiting a harder endosperm phenotype. So, the mutant was named vitreous endosperm 2 (ven2). Longitudinal dissection showed that ven2 seeds had an increased ratio of hard to soft endosperm (Fig. 1b). The hundred-seed weight of ven2 was only 23% of WT (Supplementary Fig. S1c). Starch content in ven2 seeds was 32% lower than that in WT seeds (Supplementary Fig. S1d), while zein protein levels were significantly higher in ven2 seeds, with non-zein proteins remaining unchanged (Fig. 1c−e). Correspondingly, transcript levels of most zein genes, particularly those encoding 19-kDa and 22-kDa α-zeins, were markedly elevated in the ven2 endosperm at 15 DAP (Supplementary Fig. S2). The results of SEM showed that the central region of ven2 endosperm was densely packed with smaller, smooth starch granules (SGs) embedded in a protein matrix. In contrast, WT endosperm was loosely packed with spherical SGs and lacked the protein matrix seen in ven2 (Fig. 1f−i, Supplementary Fig. S3).
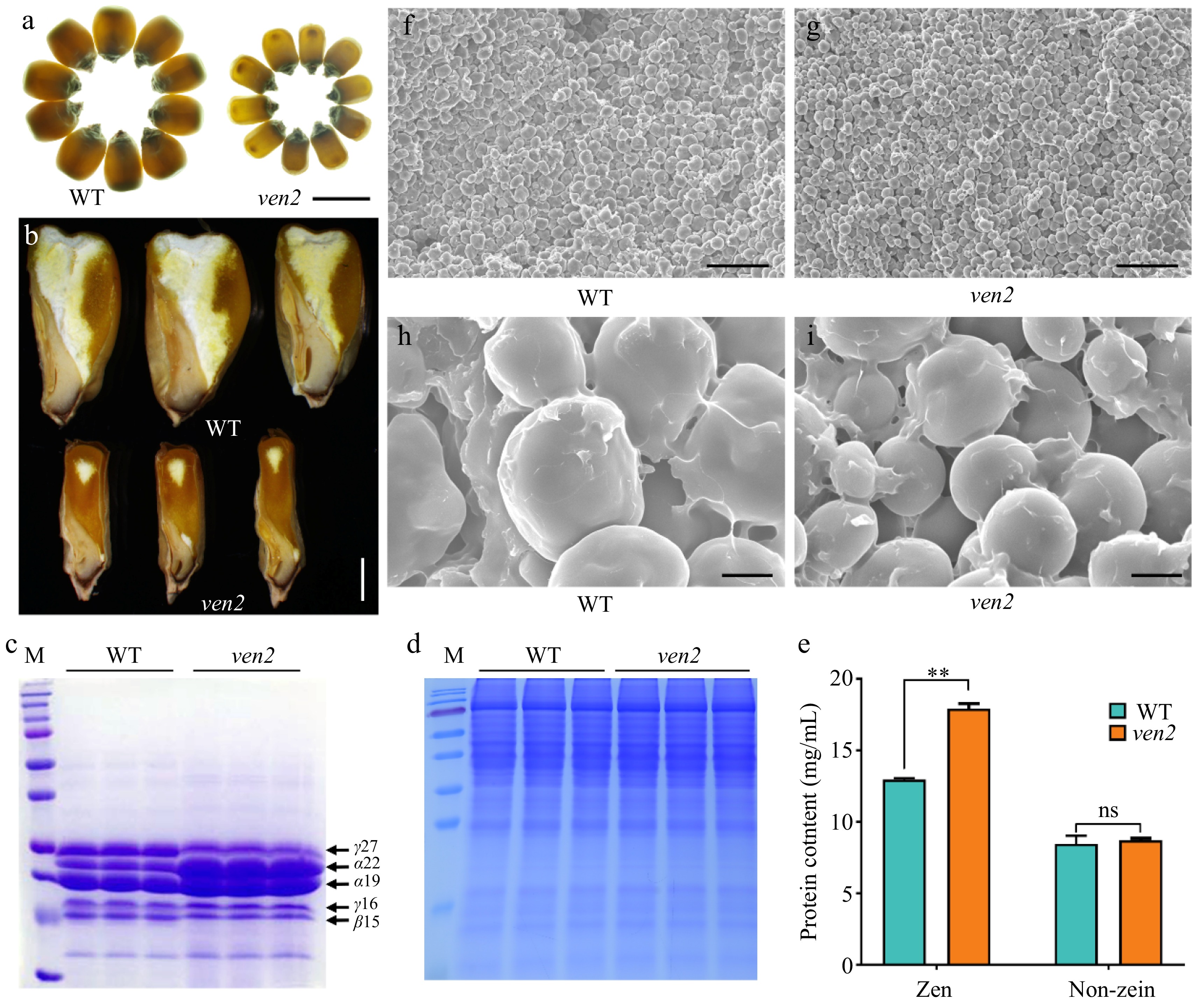
Figure 1.
Seed phenotypes of the WT and ven2 mutant. (a) WT and ven2 mature seeds viewed on a light box. Scale bars, 2 mm. (b) Sagittal sections of WT and ven2 mature seeds. Scale bars, 2 mm. SDS-PAGE analysis of (c) endosperm zein and (d) non-zein protein accumulation in WT and ven2 mutant. (e) Quantification of zein, non-zein protein contents in WT and ven2 mature endosperm. (f), (g) SEM of the WT and the ven2 endosperm cells at maturity. Scale bars, 50 μm. (h), (i) The magnified sections from the (f) and (g) panels. Scale bars, 5 μm.
Map-based cloning of Ven2 gene
-
To identify the gene responsible for the phenotype of ven2 mutant, we conducted genetic mapping using an F2 population derived from a cross between C733 and ven2. After analyzing 1,547 F2 plants displaying the ven2 phenotype with molecular markers (Supplementary Table S2), we narrowed the locus to a 0.28 Mb genomic interval on Chromosome 2, located between the markers 28z-27 and c-3799-2. According to the maize version 4 database, four open reading frames (ORFs) were identified within this mapped region (Supplementary Fig. S4). Sequencing revealed an EMS-induced G/A transition in the second exon of the Zm00001d003776 gene, leading to a conversion from aspartate (Asp) to asparagine (Asn) (Fig. 2a). Thus, the Zm00001d003776 gene is the candidate gene and designed as Ven2, which was the same gene as cell wall invertase II (INCW2) in maize.
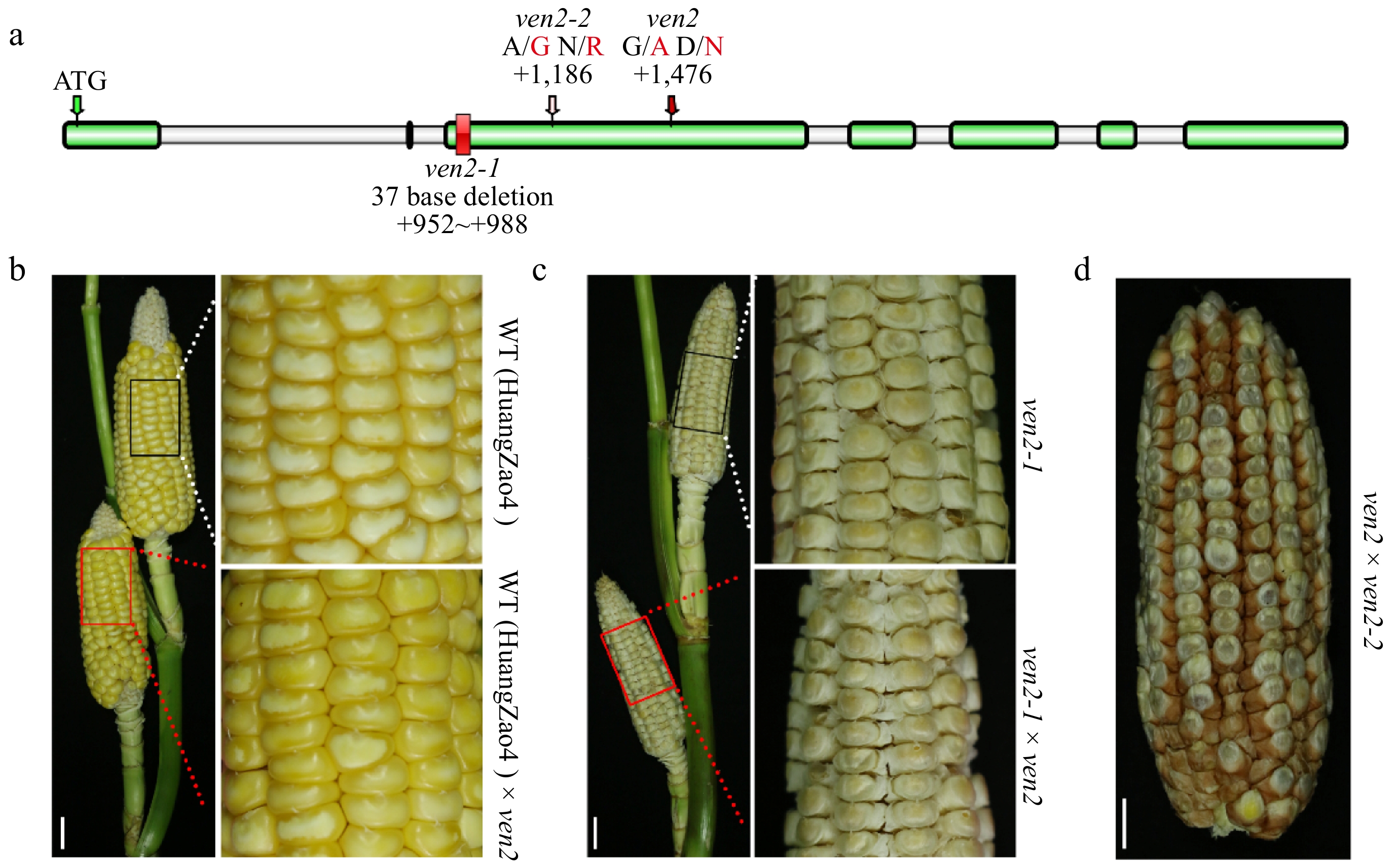
Figure 2.
Positional cloning and identification of Ven2. (a) Structure and mutation site of the Zm00001d003776 gene. Gray and green boxes represent introns and exons, respectively. (b) Allelism test using homozygous WT, ven2, ven2-1, and ven2-2. Scale bars, 3 cm.
To confirm that the identified mutation in Ven2 is the causal gene for the ven2 phenotype, we obtained two additional mutant alleles: a spontaneous small kernel mutation in maize, ven2-1 (from the HuangZao4 background) and ven2-2 (from the W22 background) from the Maize Genetic Cooperation Stock Center, which are new allelic mutants. In the ven2-1 mutant, a deletion of 37 bp from positions 952 to 988 in Ven2 gene leads to early termination of protein translation, resulting in significantly increased zein protein levels (Fig. 2a; Supplementary Fig. S5). The mutant ven2-1 exhibited a higher ratio of vitreous endosperm, smaller starch granules (SGs), and more protein bodies compared to WT (HuangZao4) (Supplementary Fig. S6). Thus, ven2-1 shares a similar phenotype with ven2. In the ven2-2 mutant, a single nucleotide change (A to G) occurs in the Ven2 gene, resulting in a codon change from CAG (Gln/N) to CGG (Arg/R) (Fig. 2a). Allelism tests were conducted by crossing ven2-1 and ven2-2 with the ven2 mutant. All F1 seeds exhibited the ven2 mutant phenotype (Fig. 2b−d). These results confirm that the loss of function in Ven2 is responsible for the ven2 mutant phenotype.
Ven2/INCW2 is primarily functions to hydrolyze sucrose into hexoses[31,38,39]. To evaluate sugar content in WT and ven2 seeds, we employed a sensitive HPLC-based detection method. Our results showed that fructose (Fru) and glucose (Glc) levels decreased in the basal sections of ven2 seeds at 6, 12, 18, and 24 DAP. Conversely, sucrose (Suc) levels increased in the basal ven2 seeds (Supplementary Fig. S7). Interestingly, Glc and Fru levels increased in the upper sections of ven2 seeds at 6, 12, and 18 DAP. However, Suc levels remained unchanged at 6 and 12 DAP. Overall, there was no significant change in Glc and Fru levels in the entire seeds, but Suc levels increased in ven2 seeds (Supplementary Fig. S7). These findings confirmed that the loss of function of Ven2/Incw2 results in sucrose accumulation during early seed development.
19 kDa α-zein and 22 kDa α-zein are regulated by a sucrose-inducible promoter
-
Sugars play a dual role in plants, serving as both energy sources and signaling molecules that regulate gene expression and development[40]. To investigate whether the accumulation of zein in the ven2 mutant is linked to the changes in sugar content, we analyzed the promoters of the 19 kDa α-zein and 22 kDa α-zein genes. Notably, the promoters of both genes contain several W-box cis-elements that respond to sugar signals (Fig. 3a).
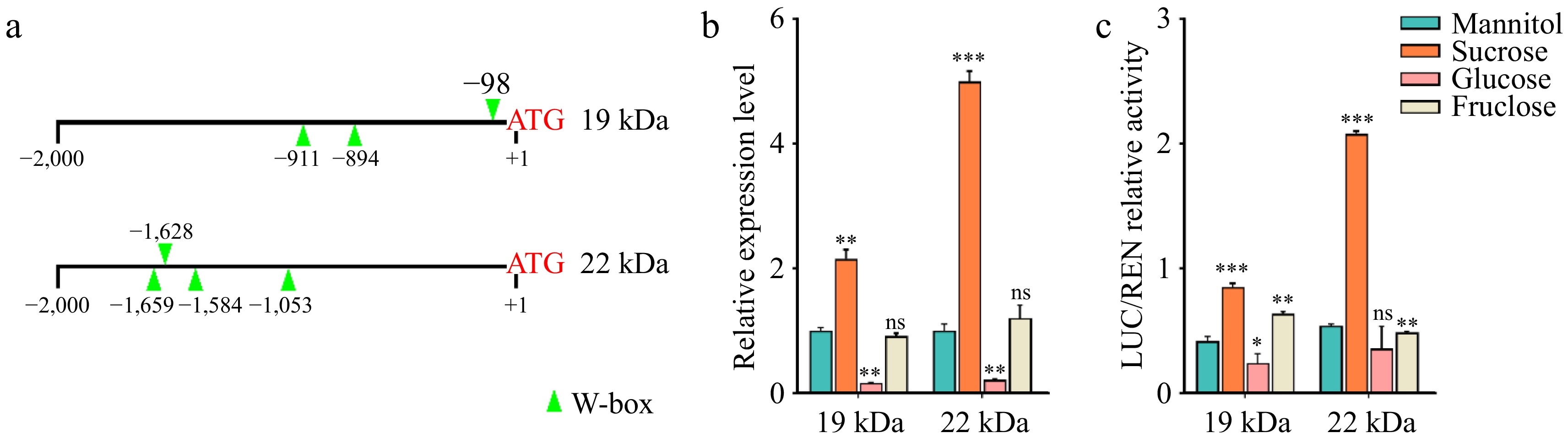
Figure 3.
Sucrose induces 19 kDa α-zein and 22 kDa α-zein gene expression. (a) Promoter analysis of 19 kDa α-zein and 22 kDa α-zein gene. W-box is a cis-acting element that responds to sugar signals. (b) Expression analysis of 19 kDa α-zein and 22kDa α-zein in different sugar treatment by qRT-PCR. Normalization was performed against the maize Actin gene (Zm00001d010159). Values represent the mean and SD of three biological replicates. **, p < 0.01; ***, p < 0.001; ns, no significant difference; student's t-test. (c) The LUC/REN relative activity of the promoter fragments including W-box of 19 kDa α-zein and 22 kDa α-zein gene. The expression level of REN was used as an internal control. Values represent the mean and SD of three biological replicates. *, p < 0.05; **, p < 0.01; ***, p < 0.001; ns, no significant difference; student's t-test.
To determine if sugar induces the expression of these genes, we cultured 16 DAP endosperm from WT in Murashige and Skoog (MS) medium supplemented with mannitol, sucrose, glucose, and fructose, respectively. The expression levels of the 19 kDa and 22 kDa α-zein genes were subsequently measured. The results indicated that transcripts for both genes accumulated in the endosperm treated with sucrose (Fig. 3b). To further explore the role of the W-box element in regulating zein gene expression, we conducted transient assays using chimeric promoters of the 19 kDa and 22 kDa α-zein genes, fused to the luciferase (LUC) reporter gene in transformed maize endosperm. The total length of about 1.5 kb promoters was linked to the LUC reporter, and the transformed endosperm was cultured on various sugar-treated media. Results revealed a significant increase in relative LUC activity in endosperm treated with sucrose (Fig. 3c). Collectively, these findings suggest that sucrose acts as a signaling molecule regulating the expression of the 19 kDa and 22 kDa α-zein genes, consistent with the observed over accumulation of zein in the ven2 mutant.
Overexpression of Ven2/Incw2 gene decreased the contents of sucrose and zein
-
To further explore the role of sucrose in regulating zein, we examined seed storage proteins in the Ven2/Incw2 overexpression transgenic line controlled by the 27 kDa γ-zein promoter (OE-Incw2). Longitudinal analysis of dry seeds from both the control (CK) and OE-Incw2 lines revealed a decreased ratio of hard to soft endosperm in the OE-Incw2 transgenic seeds (Fig. 4a, b). Transverse sections of mature seeds displayed a significantly thinner layer of vitreous endosperm in OE-Incw2 compared to CK (Fig. 4c, d). CBB-staining of protein gels indicated a marked reduction in the levels of 19 kDa α-zein and 22 kDa α-zein proteins in OE-Incw2, a finding confirmed by quantitative measurements of protein content (Fig. 4e, f). In line with these results, qRT-PCR analysis revealed that the transcript levels of both 19 kDa α-zein and 22 kDa α-zein were significantly down-regulated in the endosperm of OE-Incw2 compared to CK (Fig. 4g).
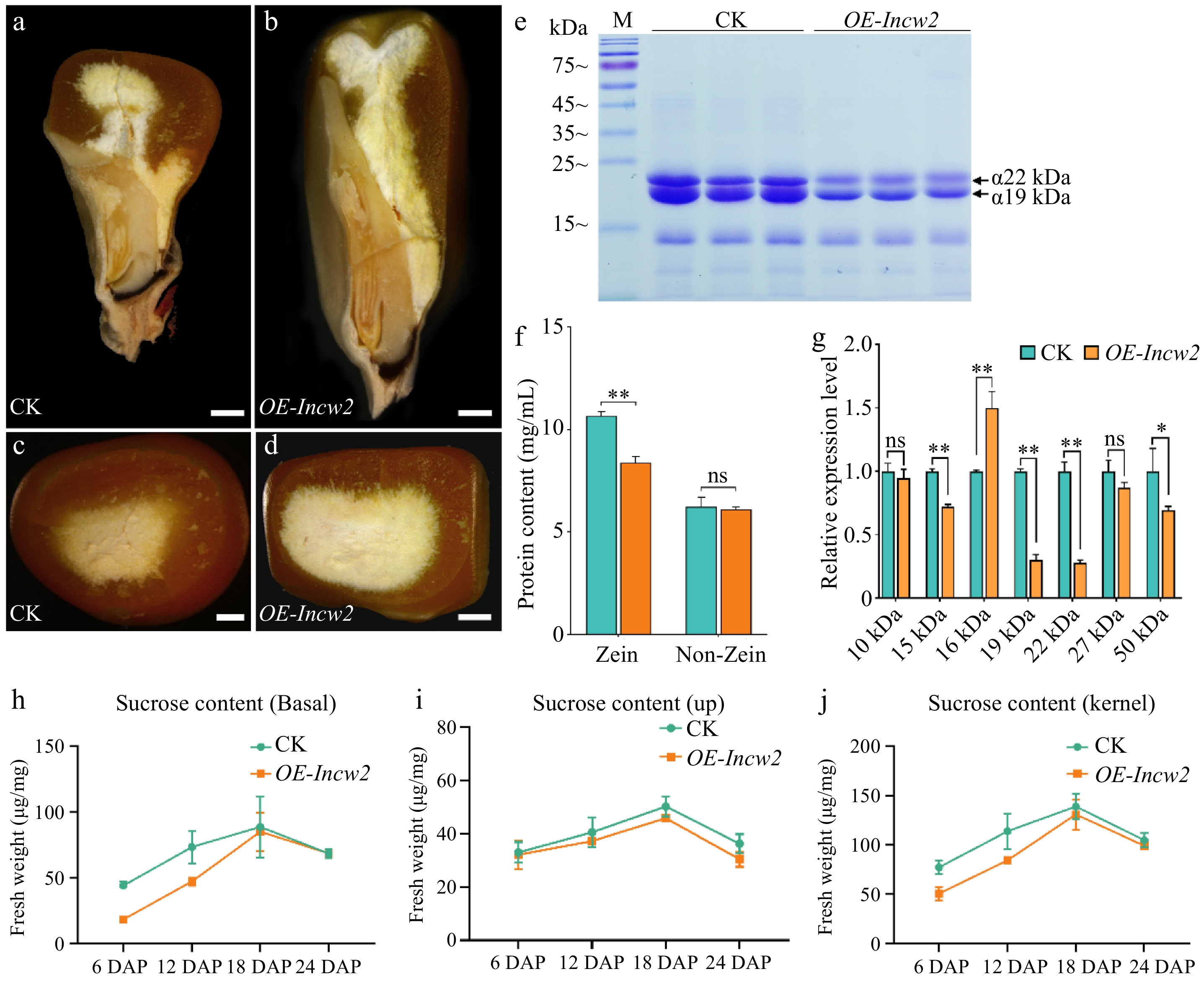
Figure 4.
Seed phenotypes of the CK and OE-Incw2. (a), (b) Longitudinal dissection of CK and OE-Incw2 mature seeds. CK, non-transgenic control. Scale bars, 2 mm. (c), (d) Transection of CK and OE-Incw2 mature seeds. Scale bars, 2 mm. (e) SDS-PAGE analysis of zein in CK and OE-Incw2. (f) Quantification of zein and non-zein contents in CK and OE-Incw2 mature endosperm. Values represent the mean and SD of three biological replicates. **, p < 0.01; ns, no significant difference; student's t-test. (g) qRT-PCR analysis of the expression of zein genes. *, p < 0.05; **, p < 0.01; ns, no significant difference; student's t-test. (h)−(j) Sucrose contents of CK and OE-Incw2 developing seeds. Values represent the mean and SD of three biological replicates.
Moreover, we analyzed the sucrose content of seeds at various developmental stages. The results showed a significant decrease in sucrose levels in OE-Incw2 seeds during early development (Fig. 4h−j). This reduction in sucrose may explain the decreased zein content and lower transcript levels of 19 kDa α-zein and 22 kDa α-zein observed in OE-Incw2 seeds.
SRF1 responds to sugar signals
-
To investigate the developmental defects in the ven2 mutant and the regulatory relationship between sugar metabolism and storage proteins, we conducted RNA sequencing (RNA-seq) on endosperm samples from both WT and ven2 at 13 DAP. We identified a total of 2,501 significant differentially expressed genes (DEGs) between WT and the ven2 mutant, with 1,481 genes upregulated and 1,020 genes downregulated in ven2 (Supplementary Fig. S8a). Gene ontology (GO) enrichment analysis showed that DEGs were primarily associated with metabolic, catalytic, and cellular processes (Supplementary Fig. S8b). Additionally, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis revealed that DEGs were mainly clustered in the biological processes of starch and sucrose metabolism and amino acid biosynthesis (Supplementary Fig. S8c). Notably, the ven2 mutant endosperm exhibited upregulation of genes related to the synthesis and metabolism of storage proteins, while genes involved in starch biosynthesis were downregulated. This pattern aligns with the observed phenotype of reduced starch and increased protein content in the mutant (Supplementary Fig. S9). These findings suggest that the altered starch and protein contents in the ven2 mutant may be partially due to the differential regulation of genes associated with starch and protein biosynthesis.
To investigate the mechanism by which sugar signals regulate zein expression, we analyzed the expression levels of transcription factors from the transcriptome data (Supplementary Fig. S9). We utilized the PLACE database (
https://sogo.dna.affrc.go.jp/cgi-bin/sogo.cgi ) to examine the promoters of these transcription factors and discovered that the promoter of the WRKY transcription factor SRF1 contains several cis-acting elements responsive to sugar signals (Supplementary Fig. S10a). To confirm that the expression of SRF1 is regulated by sugar signaling, we treated wild-type endosperm with sucrose, using mannitol as a control. The results indicated that SRF1 expression was significantly elevated in endosperm treated with sucrose (Supplementary Fig. S10b). Additionally, the ven2 mutant seeds, which exhibited increased sucrose content compared to WT, also showed a significant increase in SRF1 expression (Supplementary Fig. S10c). Conversely, the decrease of sucrose content in endosperm was accompanied by the decrease of SRF1 gene expression in OE-Incw2 endosperm (Supplementary Fig. S10d). These findings suggest that sugar signals positively regulate the expression of SRF1 in maize.SRF1 transcriptionally regulates 19 kDa α-zein and 22 kDa α-zein gene expression
-
To determine whether the transcription factor SRF1 directly activates the expression of the α-zein genes, we performed a dual-luciferase transcriptional activity assay. This assay utilized a firefly luciferase (LUC) reporter system, specifically designed with the promoters of the 19 kDa and 22 kDa α-zein genes. By co-expressing SRF1 along with the LUC reporter in tobacco leaves, we aimed to assess any resultant changes in luciferase activity. The experimental results revealed a significant increase in LUC activity in the presence of SRF1, indicating that this transcription factor indeed enhances the transcriptional activity of the α-zein promoters (Fig. 5a).
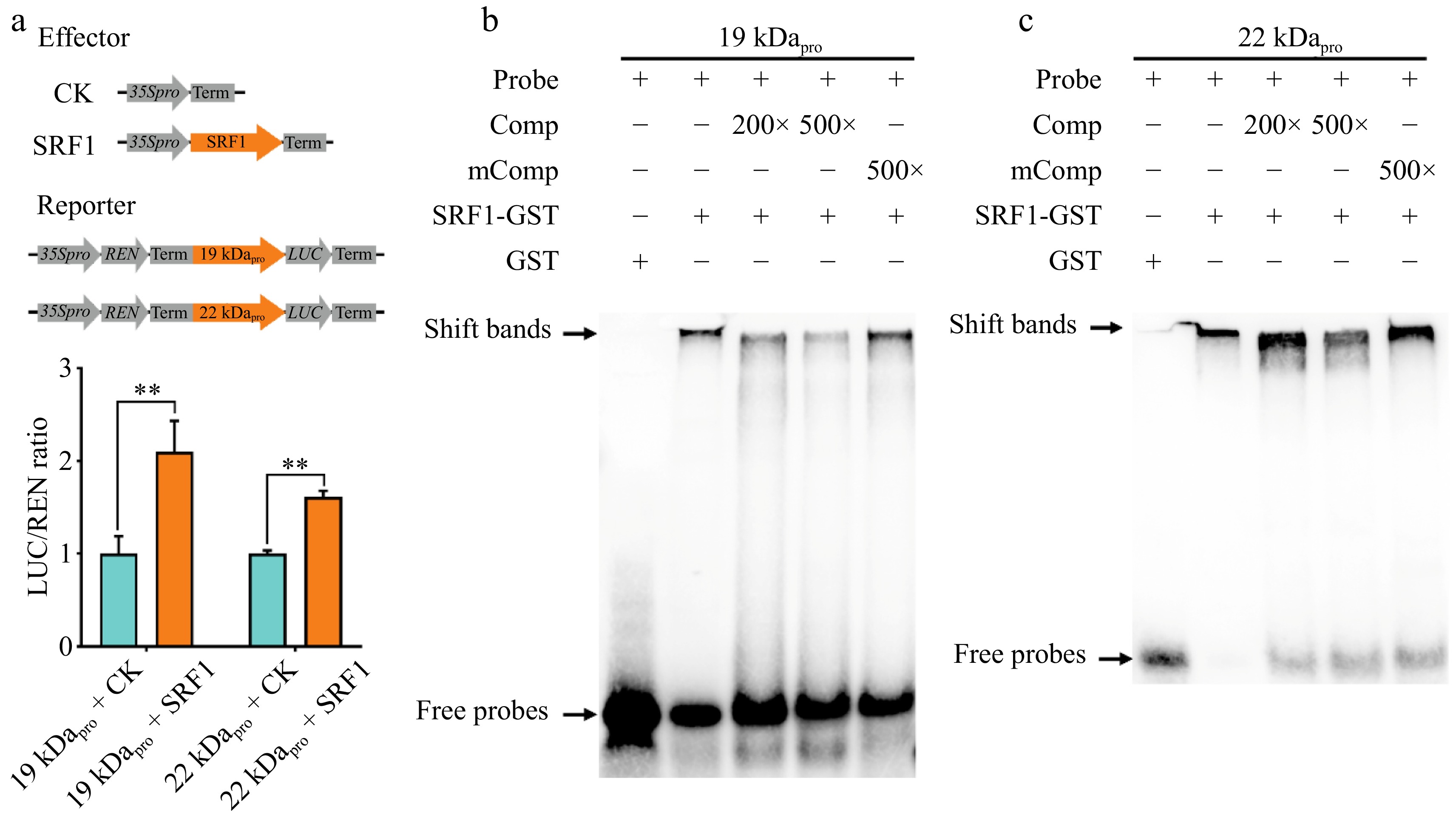
Figure 5.
Identification of cis-elements and the associated SRF1 for 19 kDa and 22 kDa promoter transactivation. (a) DLR assay of transactivation of the 19 kDa and 22 kDa zein genes promoter by SRF1 in tobacco leaves. (b), (c) EMSA of the specific binding of SRF1-GST to the W-box motifs in the 19 kDa and 22 kDa zein genes promoter. The normal and mutant probes were labeled with biotin. Unlabeled intact probes were used for competition. Comp, competing probes unlabeled with biotin.
To further elucidate the binding interaction between SRF1 and the W-box element within the promoters, we employed eight biotin-labeled probes that were derived from the specific promoter regions of the 19 kDa and 22 kDa α-zein genes in EMSA. It clearly demonstrated that SRF1 can bind directly to the W-box in the promoters of both α-zein genes in vitro (Fig. 5b, c). This finding supports the hypothesis that SRF1 plays a critical role in regulating the transcription of α-zein genes through direct interaction with their promoter regions.
-
Impaired INCW2 function disrupts normal sugar metabolism and storage protein synthesis, leading to significant alterations in seed development. INCW2, which encodes a cell wall invertase, plays a crucial role in the breakdown of sucrose into its component hexoses, which are essential for energy metabolism and proper biosynthesis of starch and proteins. When INCW2 is nonfunctional, sucrose accumulation in the basal endosperm is elevated, while the conversion of sucrose to hexoses is impaired[39]. This disruption not only affects energy availability for storage but also leads to an imbalance in the metabolic pathways responsible for synthesizing storage proteins, such as zeins, which are critical for seed structure and nutrient storage. As a result, the altered sugar and protein metabolism negatively impacts seed size, quality, and overall development, highlighting the essential role of INCW2 in maintaining metabolic homeostasis during seed maturation.
Dysfunction of INCW2 leads to reduced invertase activity, resulting in impaired metabolic flux of sugar utilization in the developing endosperm[39,41]. The altered sugar profile in ven2 is attributed not only to the loss of catalytic activity of the cell wall invertase but also to the regulation of sugar transport-related genes. During the filling stage, sucrose can be directly transferred to the endosperm through the BETL via specific sucrose transporters, such as SUGCAR1, ZmSUT1, and ZmSWEET11/13a[42−44]. In the ven2 endosperm, several DEGs associated with starch biosynthesis, including Brittle2 (Bt2), Amylose extender1 (Ae1), and Shrunken2 (Sh2)[45], were significantly downregulated (Supplementary Fig. S9). This downregulation inhibits the conversion of sugar to starch, resulting in elevated sugar levels in the endosperm of ven2. Additionally, the increase in sugar content in ven2 may also be partially due to the upregulation of sugar transport-related genes, such as Sucrose transporter1 (SUT1) (Supplementary Fig. S9). In summary, INCW2 plays a crucial role in regulating seed weight and starch content through its influence on sugar metabolism.
Previous research indicated that mutations in INCW2, localized in the basal endosperm transfer cell layer, resulted in a loss of over 70% of seed weight at maturity due to reductions in both cell number and size during endosperm development[31,46−48]. Further investigations revealed that the decreased cell number and size in the mn1 mutant were attributed to deficiencies in various phytohormones[47,49]. In our study, we observed that the zein protein content in the ven2 and ven2-1 mutants was significantly increased, particularly for the 19 kDa and 22 kDa zein proteins, compared to the WT (Fig. 1c−e; Supplementary Fig. S5). In contrast, zein protein content decreased when INCW2 was specifically overexpressed in maize BETLs (Fig. 4e, f). Unlike previous studies, our findings demonstrate that INCW2 influences not only seed weight but also storage protein levels. Collectively, these results indicate that INCW2 has a pleiotropic effect on seed development.
Transcriptional regulation of zein genes by SRF1
-
Transcriptional regulation of zein genes by SRF1 involves a complex mechanism by which this transcription factor modulates the expression of genes responsible for synthesizing storage proteins in maize seeds. SRF1 is a key regulator that responds to changes in sugar levels, particularly sucrose, which acts as both an energy source and a signaling molecule during seed development. When sucrose levels increase, SRF1 is activated and binds directly to the promoter regions of the 19 kDa and 22 kDa α-zein genes, which encode major storage proteins in the endosperm. Through this binding, SRF1 enhances the transcription of these genes, facilitating the production of α-zeins, which are critical for seed protein storage. This regulatory pathway ensures that adequate storage proteins are synthesized to support seed growth and development. In addition to its role in regulating zein gene expression, SRF1 also integrates signals from other metabolic pathways, providing a coordinated response to nutrient availability and developmental cues, which is essential for optimizing seed quality and nutrient content.
Several transcription factors, including O2, PBF1, MADS47, and others have been reported to directly activate 19 kDa and 22 kDa α-zein proteins genes and have been shown to play crucial roles in regulating the biosynthesis of maize endosperm storage proteins[9,12,15,16]. However, the external signals that influence the storage protein biosynthesis remain largely unknown. In this study, we identified a new allelic mutant of INCW2, characterized by increased levels of the 19 kDa and 22 kDa α-zein proteins, along with elevated sugar content. During seed development, approximately 90% of carbon is derived from sucrose, whose levels exhibit diurnal variations that lead to the diurnal transcription of O2-regulated zein genes[25]. Collectively, these findings suggest that sucrose serves not only as a primary carbon source but also as a critical signal in regulating starch and storage protein synthesis in plants. In ven2 mutant, transcription factors, such as, O2, PBF1, and MADS47 show no significant changes in transcriptome data. This suggests that, in addition to the known transcription factors such as O2, PBF1, and MADS47, there may be other potential transcription factors that are regulated by the sucrose signal and are involved in the regulation of zein protein synthesis.
Over the past few decades, various transcription factors, including MYB domain proteins, basic leucine zippers (bZIPs), and WRKY proteins, have been identified as key regulators of sugar responses[40,50−52]. In the ven2 mutant, elevated sucrose levels lead to increased expression of SRF1, which subsequently upregulates the transcription of genes involved in zein biosynthesis. Sequence analysis indicates that SUSIBA2 in barley and SRF1 in maize are closely related members of the WRKY superfamily of plant transcription factors. SUSIBA2 plays a role in sugar signaling in barley by binding to sugar-responsive (SURE) and W-box elements within the iso1 promoter[52,53]. Our promoter sequence analysis revealed multiple W-box cis-elements in the α-zein gene, which are bound by the transcription factor SRF1 (Fig. 3a). Additionally, the results from dual-luciferase transcriptional activity assays and electrophoretic mobility shift assays confirmed that SRF1 is essential for activating the expression of the 19 kDa and 22 kDa α-zein genes (Fig. 5). In conclusion, we proposed a model for sucrose content affects zein biosynthesis mediating by transcription factor SRF1 (Fig. 6). These findings establish SRF1 as a potential candidate regulatory transcription factor in the synthesis of storage proteins, underscore the role of a WRKY protein in the sucrose response. Therefore, further exploration of SRF1 and its regulatory mechanisms will help to deepen our understanding of the regulatory network of maize zein protein synthesis and pave the way for its future application in maize improvement.
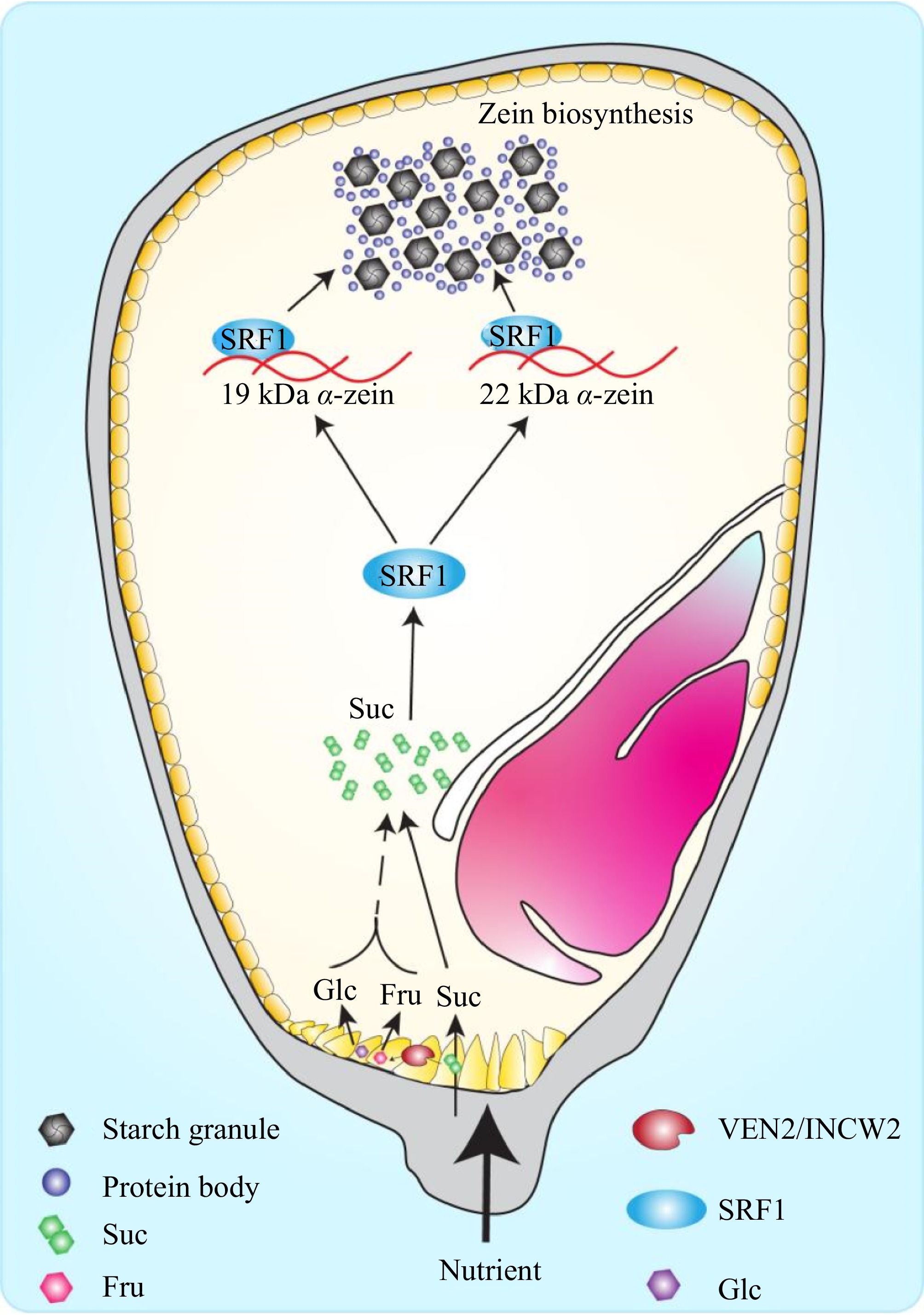
Figure 6.
A proposed model of sucrose content affects zein biosynthesis mediating by transcription factor SRF1. In the cell wall ingrowths, sucrose is hydrolyzed into hexoses (glucose and fructose) by VEN2/INCW2 and the apoplasmic sucrose and hexoses are translocated into endosperm cell. The transcription factor SRF1 responds to sucrose and regulates the expression of 19 kDa and 22 kDa α-zein by binding directly to their promoters, thereby mediating zein biosynthesis. Suc, sucrose; Fru, fructose; Glc, glucose.
-
In this work, we characterize a WRKY transcription factor SRF1, which responds to sucrose signal and regulates 19 kDa and 22 kDa α-zein gene expression, thereby affecting zein protein synthesis during seed development.
This work was supported by the National Natural Science Foundation of China (32071921), the Natural Science Foundation of Shandong (ZR2023QC162 and ZR2024QC152), Shandong Provincial University Youth Innovation and Technology Program, China (2023KJ284), and the Taishan Scholars Project. We acknowledge Professor Guangtang Pan (Sichuan Agricultural University) for providing seeds of OE-Incw2; Professor Yongcai Huang (Sichuan Agricultural University) for technical support on maize endosperm protein extraction and analysis; and The Maize Genetics Cooperation Stock Center for the seed stocks.
-
The authors confirm contribution to the paper as follows: study conception and design: Zhao X; data collection: Wei Y, Zang J, Shao D, Wang B, Zheng G, Yan R, Wu J; analysis and interpretation of results: Wei Y, Zang J, Zhao Y, Zhang C, Zhou C; draft manuscript preparation: Wei Y, Zhao X, Zhang X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files. Further information is available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yiming Wei, Jie Zang
- Supplementary Table S1 List of primers used in this study.
- Supplementary Table S2 Genetic analysis of the seeds in F2 population of the segregating ears.
- Supplementary Fig. S1 Phenotypes of the WT and ven2 mutant.
- Supplementary Fig. S2 Zein genes expression analysis by qRT-PCR.
- Supplementary Fig. S3 SEM analysis of endosperm in WT and ven2.
- Supplementary Fig. S4 Positional cloning and identification of VEN2.
- Supplementary Fig. S5 Analysis of zein and non-zein in WTHuangZao4 seeds and ven2-1 seeds.
- Supplementary Fig. S6 SEM analysis of endosperm in WTHuangZao4 and ven2-1.
- Supplementary Fig. S7 Contents of fructose, glucose, and sucrose of WT and ven2 developing seeds.
- Supplementary Fig. S8 Analysis of transcriptome data.
- Supplementary Fig. S9 Heat map analysis of differentially expressed genes.
- Supplementary Fig. S10 SRF1 responses to sugar signals.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan YazhouBay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wei Y, Zang J, Shao D, Wang B, Zheng G, et al. 2025. Sucrose signaling regulates storage protein synthesis in maize endosperm. Seed Biology 4: e010 doi: 10.48130/seedbio-0025-0008 

Sucrose signaling regulates storage protein synthesis in maize endosperm
- Received: 06 February 2025
- Revised: 14 April 2025
- Accepted: 21 April 2025
- Published online: 24 June 2025
Abstract: Sucrose plays a crucial role as a primary carbon source during seed development. In addition to this metabolic function, sucrose and its breakdown products (hexoses) serve as important signaling molecules that regulate cell fate and seed development in maize. However, the precise molecular mechanisms through which sucrose influences storage protein synthesis during seed development remain poorly understood. In this study, we identified a novel allelic mutant of Incw2, named vitreous endosperm 2 (ven2). The loss-of-function mutation in Incw2 led to a significant reduction in grain weight, accompanied by a marked increase in sucrose levels in the basal endosperm. This was also associated with substantial increases in the accumulation of the 19 kDa and 22 kDa α-zein proteins. In contrast, overexpression of Incw2 resulted in lower sucrose and zein contents. Furthermore, we discovered that the transcription factor Sugar Response Factor 1 (SRF1), which is responsive to sucrose, regulates the expression of the 19 kDa and 22 kDa α-zein genes by directly binding to their promoters. These findings suggest that disruption of sucrose metabolism due to the Incw2 mutation modulates α-zein gene expression and affects protein synthesis during seed development.
-
Key words:
- Maize /
- Storage protein /
- Endosperm /
- Sugar Response Factor 1 /
- Sucrose