








-
RNA interference (RNAi) is a crucial mechanism that regulates gene expression at both transcriptional and post-transcriptional levels[1]. It is widely observed in mammals, insects, plants, and microbes, including fungi and oomycetes[1−4]. RNAi depends on the RNA-induced silencing complex (RISC)[1−5]. RISC mainly consists of small RNAs (sRNAs), Argonaute (AGO) proteins, and Dicer-like (DCL) proteins[5]. The main types of sRNAs include microRNAs (miRNAs), small interfering RNAs (siRNAs), phased secondary siRNAs (phasiRNAs), heterochromatic siRNAs (hc-siRNAs)[5,6], and epigenetically activated small interfering RNAs (easiRNAs)[7−9].
In plants, RNAi mediated by endogenous sRNAs regulates growth, stress responses, and immunity[2,10]. It also serves as a well-established antiviral defense. The functions of each RISC component in this process have been thoroughly reviewed[11]. A recent study revealed that the plant RNAi system can disrupt vertical virus transmission via seeds, addressing a longstanding question in the field[12]. Virus-derived sRNAs are critical for repressing viral replication by cleaving nascent viral RNAs[13,14]. Invasion by bacteria, fungi, or oomycetes can also induce sRNAs. These sRNAs target negative immune regulators and enhance plant immunity[15−17].
Beyond native sRNAs, a pioneering study on erythrocyte–Plasmodium falciparum interaction demonstrated that non-self sRNAs (specifically, two erythrocyte-derived miRNAs) could trigger RNAi in P. falciparum and inhibit infection[18]. Later studies confirmed that trans-kingdom RNAi occurs in many systems. These include interactions between plants and fungi, oomycetes, bacteria, and even other plants[19−24]. Growing evidence suggests that trans-kingdom RNAi plays a major role in the evolutionary arms race between plants and microbes[25,26]. Pathogens can export sRNAs into host cells. These sRNAs suppress immune gene expression and promote infection. In defense, plants produce extracellular vesicles (EVs). These vesicles deliver sRNAs or mRNAs into pathogen cells to inhibit virulence[27,28].
Plants can be engineered to express artificial sRNAs that target pathogen virulence genes. This strategy, known as host-induced gene silencing (HIGS), relies on transgenic approaches[29−32]. Alternatively, spray-induced gene silencing (SIGS) offers a transgene-free method. In SIGS, sRNAs are loaded onto nanoparticles and sprayed onto plants. This protects the sRNAs from degradation and helps suppress pathogen infection[33,34]. A novel strategy, microbe-induced gene silencing (MIGS), has recently been developed. In this approach, beneficial rhizospheric fungi such as Trichoderma harzianum are engineered to produce sRNAs targeting pathogen virulence genes. These fungi can effectively suppress a wide range of pathogens. MIGS does not involve host plant transformation. It offers a promising and sustainable tool for crop protection[35,36].
However, early studies on plant–virus interactions identified several viral proteins as viral suppressors of RNA silencing (VSRs). These include 2b from cucumber mosaic virus, P19 from tomato bushy stunt virus, and HC-Pro from potyviruses[11]. These proteins impair host RNAi by disrupting RISC function. More recently, a VSR was found to block RNAi initiation by interfering with calcium signaling in plants[37]. Such proteins are now termed suppressors of trans-kingdom RNAi (STRs). STRs may weaken RNAi-based strategies such as HIGS, SIGS, and MIGS. As a result, researchers have focused on identifying RNAi suppressors in viruses, fungi, and oomycetes. Notably, fungal and oomycete STRs seem to act through mechanisms distinct from those of viral suppressors[38,39].
In this review, we explore the current understanding of trans-kingdom RNAi-mediated defense and counter-defense mechanisms in plant–pathogen interactions and discuss their potential applications in crop protection.
-
Double-stranded RNA (dsRNA) is a key molecule in triggering RNAi across eukaryotic cells, as first demonstrated by dsRNA injection in Caenorhabditis elegans[40]. RNAi can also be induced by feeding C. elegans with Escherichia coli expressing dsRNA, paving the way for RNAi-based crop protection strategies against insect pests[41]. Leveraging the conserved RNAi machinery in eukaryotes, researchers began engineering plants to produce dsRNAs targeting pathogen genes. These dsRNAs can be transferred into pathogen cells, silencing virulence-related genes and enabling HIGS-mediated defense.
A well-known example is transgenic cotton expressing dsRNAs targeting the P450 monooxygenase gene of Helicoverpa armigera (cotton bollworm), which significantly reduced pest damage[42]. Similarly, silencing plant oncogenes reduced crown gall formation caused by Agrobacterium tumefaciens[43]. These findings encouraged further research into HIGS applications in plant-fungal interaction. For instance, targeting the CYP51 gene in Fusarium graminearum (involved in ergosterol biosynthesis) impaired fungal growth and infection[44]. HIGS has also been successfully applied to combat Blumeria graminis infection[45], and Verticillium dahliae invasion[31]. These examples highlight the potential of properly designed RNAi constructs in enhancing plant resistance through HIGS.
Unlike these cases involving artificial sRNAs, the role of native sRNAs in HIGS was discovered more recently (Fig. 1). Two endogenous cotton miRNAs were found to accumulate during V. dahliae infection and to target the pathogen virulence genes. These two miRNAs were found in V. dahliae cells to turn down virulence-related gene expression, which successfully disturbed the pathogen invasion in cotton[20]. Similarly, phasiRNAs derived from pentatricopeptide-repeat (PPR) protein gene loci in plants were shown to silence virulence genes in Phytophthora, enhancing plant immunity[22]. Despite the effectiveness of both artificial and native sRNAs in HIGS, the mechanisms by which these molecules are taken up by pathogen cells remain under investigation.
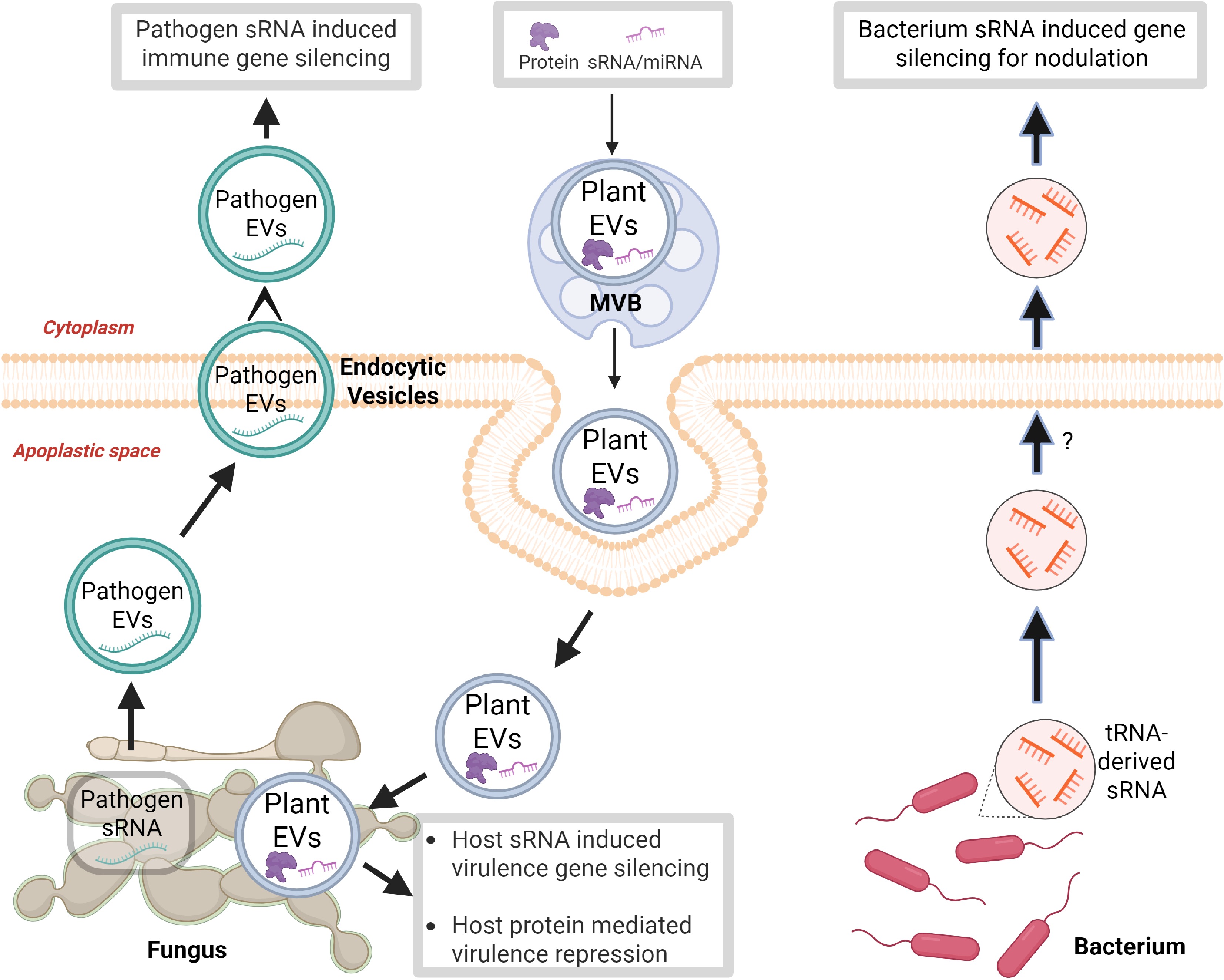
Figure 1.
RNA communication between plants and microbes. Plants deliver small RNAs (sRNAs) and proteins via extracellular vesicles (EVs) to silence virulence genes and suppress pathogenicity in microbes. In return, fungal and bacterial pathogens release EVs containing sRNAs that target and suppress plant immune genes. Additionally, rhizobial bacteria use tRNA-derived sRNAs to modulate plant gene expression for nodulation. This figure illustrates a bidirectional trans-kingdom RNA interference (RNAi) mechanism in plant–microbe interactions. This figure was created in BioRender by Zhu C, 2025,
https://BioRender.com/26gx2vx .In mammals, RNAs are known to mediate intercellular communication via extracellular vesicles (EVs) such as exosomes[46,47]. A landmark study in the Arabidopsis-Botrytis cinerea system demonstrated that plant-derived miRNAs, phasiRNAs, and hc-siRNAs are delivered into fungal cells via EVs[27]. These sRNAs efficiently suppressed virulence gene expression in B. cinerea, enhancing plant resistance (Fig. 1). More recently, studies in the Oryza sativa–Rhizoctonia solani system revealed that rice EVs not only carry sRNAs but are also enriched with defense proteins upon infection. Uptake of these proteins by the fungus or their overexpression in rice significantly enhanced disease resistance[28].
In summary, HIGS represents a powerful and feasible approach for enhancing crop resistance through trans-kingdom RNAi. Its integration into crop breeding strategies holds great promise for sustainable agricultural disease management.
-
Effector proteins and secondary metabolites are well-known virulence factors that pathogens use to suppress plant immunity[45,48]. Recently, sRNAs have emerged as potential virulence factors in plant–pathogen interactions.
The first evidence came from the Botrytis cinerea-plant system (Fig. 1). Plant AGO1 immunoprecipitation identified B. cinerea sRNAs that suppressed host resistance genes, demonstrating their role in pathogenicity[19]. Similarly, a 22-nt miRNA from the parasitic plant Cuscuta pentagona was shown to move into host plants, aiding parasitism[21]. Moreover, three tRNA-derived sRNAs from Rhizobium interacted with soybean AGO1 to regulate root nodulation genes, highlighting sRNA function in trans-kingdom communication (Fig. 1)[23]. In Verticillium dahliae-infected plants, a 24-nt rRNA-derived sRNA upregulated MIR157d, delaying flowering and benefiting pathogen nutrient uptake[49].
Some pathogen sRNAs promote virulence without AGO1 binding. For example, B. cinerea sRNA Bc-siR37 suppresses three immune genes, increasing host susceptibility[50]. In wheat, the stripe rust pathogen Puccinia striiformis f. sp. tritici (Pst) produced a microRNA-like sRNA, Pst-milR1, that repressed PR2 gene expression, weakening plant immunity[51]. Despite their clear impact, the delivery methods of these sRNAs remained unknown.
A breakthrough came with the isolation and sequencing of B. cinerea EVs from infected plant tissues, which revealed that sRNA effectors are packaged and transported via EVs (Fig. 1)[52]. This trans-kingdom RNAi-based mechanism, termed pathogen-induced gene silencing (PIGS), is a key virulence strategy in phytopathogens (Fig. 1).
-
Plant viruses must replicate inside host cells to survive, but host RNAi machinery often blocks this process[14]. To counteract this, viruses produce viral suppressors of RNA silencing (VSRs) protein that inhibit RNAi pathways[11]. VSRs were first identified using a reporter system, where co-expression of a VSR and a GFP reporter in plants restored GFP fluorescence by preventing RNAi-based silencing[53,54]. Later studies showed that VSRs target multiple RNAi steps, including sRNA biogenesis, transport, and AGO loading[11].
While viral RNAi suppressors are well studied, RNAi suppressors in eukaryotic pathogens remained unknown until the discovery of Phytophthora suppressors PSR1 and PSR2[38]. Expression of either PSR1 or PSR2 significantly increased Phytophthora sojae infectivity, highlighting their role in pathogenicity (Fig. 2). The target of PSR1, PSR1-Interacting Protein 1 (PINP1), is a key pre-mRNA splicing factor that regulates alternative splicing of resistance-related sRNAs and mRNAs. This interaction explains how PSR1 impairs plant RNAi immunity[55−57] (Fig. 2). PSR2 interferes with pathogen-induced phasiRNA biosynthesis. In the Phytophthora capsici-host system, phasiRNAs derived from plant PPR genes (e.g., siR1310 and siR0513) target the virulence gene for degradation. PSR2 binds to dsRNA-binding protein 4 (DRB4), suppressing phasiRNA production and weakening host defense[22] (Fig. 2). Structural analysis identified a conserved L-W-Y motif in PSR2 essential for its virulence function[58].
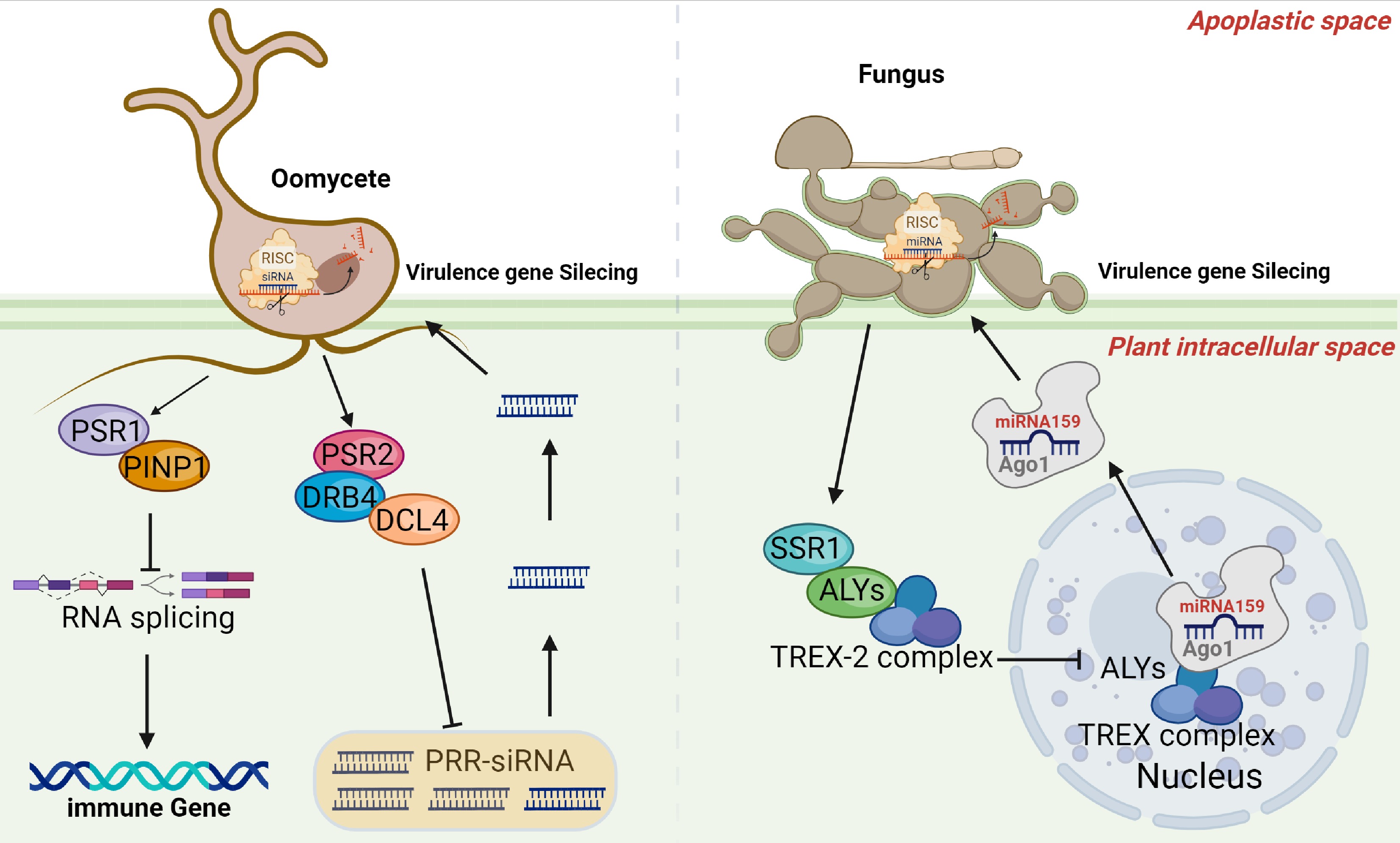
Figure 2.
Suppressor of trans-kingdom RNAi proteins. Oomycetes and fungi deploy STR proteins to block plant RNAi-based immunity. In oomycetes, PSR1 interferes with host RNA splicing, while PSR2 inhibits DRB4-DCL4 complex activity, reducing production of PRR-derived siRNAs which trigger HIGS defense. In fungi (V. dahliae), SSR1 disrupts the TREX complex by binding ALY proteins, impairing nuclear export of AGO1–miRNA complexes and reducing miRNA accumulation in fungal cells, thereby enhancing pathogen virulence. This figure was created in BioRender by Zhu C, 2025,
https://BioRender.com/s4von3m .An antagonistic mechanism is also found in the phytopathogenic fungus V. dahliae. A non-canonical effector named VdSSR1 (secretory silencing repressor 1) shows strong RNAi suppressor activity (Fig. 2). VdSSR1 interacts with ALY proteins, adaptors of the TREX complex, disrupting nuclear export of AGO1–miRNA complexes. This reduces cytoplasmic AGO1 and sRNA levels, suppressing mobile plant miRNA accumulation in fungal cells and enhancing pathogen virulence[39].
Collectively, these findings emphasize that the suppression of plant sRNA-based trans-kingdom RNAi immunity through VSR-like proteins during phytopathogen-host interaction.
-
HIGS has been widely applied to defend against insects, fungi, and oomycetes, with numerous successful cases reported (Fig. 3)[30,31,59]. The development and implementation of HIGS technology follows several key principles:
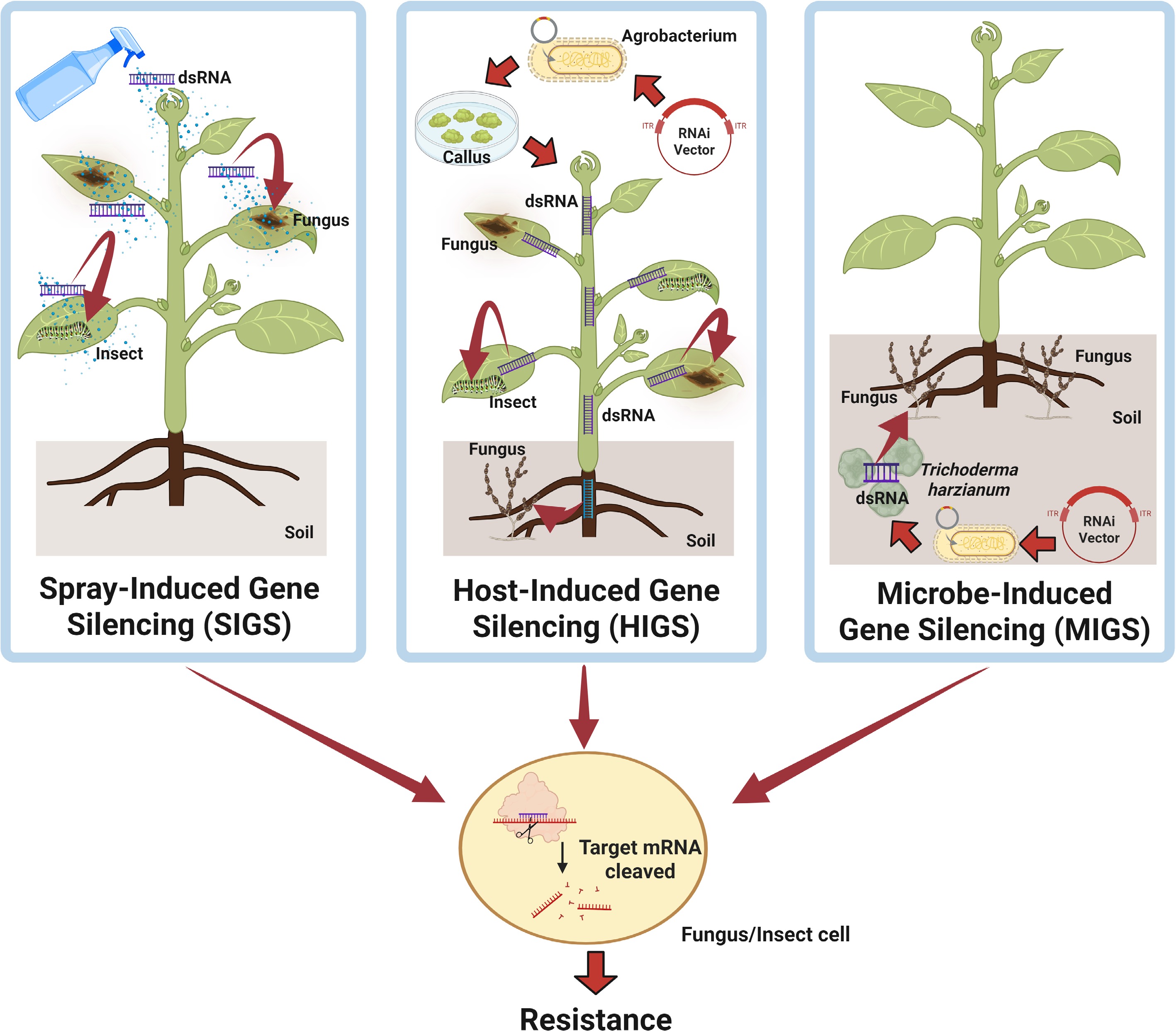
Figure 3.
Trans-kingdom RNAi in plant breeding. This diagram illustrates three RNAi-based approaches used in plant disease management: (1) SIGS: Externally applied dsRNAs or sRNAs are absorbed by pathogens on plant surfaces, leading to gene silencing without transgenic modification. (2) HIGS: Transgenic plants express dsRNAs targeting pathogen virulence genes, which are taken up by invading pathogens. (3) MIGS: Beneficial fungi such as Trichoderma harzianum deliver sRNAs into soil-borne pathogens, silencing virulence-related genes and offering a non-transgenic, environmentally friendly solution. This figure was created in BioRender by Zhu C, 2025,
https://BioRender.com/fvszcsn .Target gene selection
-
The chosen target gene should be essential for pathogen or insect development or pathogenicity. For fungi, genes involved in RNAi pathways, vesicle trafficking, ergosterol biosynthesis, chitin synthesis, or lipid catabolism are effective targets[60]. Proteins related to host tissue penetration and other disease-associated proteins are also ideal candidates[60]. For insects, genes conferring tolerance to plant-produced insecticides such as gossypol or Bacillus thuringiensis toxins serve as good targets. The target gene should be highly expressed across the lifespan and tissues. Designing dsRNA chimeras that target multiple genes or different regions of one gene can improve silencing efficiency.
Expression system design
-
RNAi constructs must be optimized for each crop. Hairpin constructs (sense-intron-antisense) are more effective at producing dsRNAs than single antisense constructs. Promoters should either drive expression constitutively across tissues or spatiotemporally in tissues attacked by fungi or insects. Using native promoters such as those from UBIQUITIN or GAPDH genes helps reduce epigenetic silencing in progeny. The intron in hairpin constructs should be efficiently recognized by crop cells to facilitate dsRNA processing. Expression levels of dsRNA must be balanced to avoid stress responses that could reduce yield.
Off-target effect
-
Although gene silencing by RNAi is highly specific, off-target effects still occur[61]. A recent study[62] suggests criteria to minimize these effects: (1) target sequence identity should exceed 80%; (2) dsRNA must match at least 16 base pairs perfectly without mismatches; and (3) continuous mismatches within 19 base pairs pose a high risk of target failure. Online databases for designing low off-target dsRNAs have been documented by Mosquera et al.[60]. Off-target effects on beneficial fungi or insects are rarely addressed but may exist. Therefore, further ecotoxicological studies on HIGS are necessary.
MIGS in plant resistance breeding
-
MIGS has recently emerged as a promising strategy in plant disease management (Fig. 3). The beneficial fungus Trichoderma harzianum is widely used as a soil biocontrol agent and is genetically tractable. These features make it well-suited for delivering small RNAs (sRNAs). A recent study demonstrated that T. harzianum can transmit sRNAs into the cells of two major soil-borne pathogens, V. dahliae and Fusarium oxysporum, resulting in the silencing of virulence-related genes[35]. MIGS provides a non-transgenic approach for managing soilborne pathogens. Moreover, recent findings have confirmed the effectiveness of MIGS against Fusarium graminearum, further supporting its potential in protecting a broad range of crops from diverse fungal pathogens[36].
MIGS combines the advantages of HIGS and SIGS: (1) It avoids the need for plant transformation, unlike HIGS; (2) It overcomes the field application challenges faced by SIGS; and (3) It provides an efficient and environmentally friendly solution for controlling soilborne diseases without reducing crop productivity.
-
Based on the principles of HIGS, efficient delivery and uptake of small RNAs (sRNAs) targeting pathogen virulence genes are critical for enhancing plant immunity. A long-standing question, however, was whether exogenously applied artificial sRNAs could be taken up by pathogens in the absence of transgenic plants.
Pioneering studies addressed this question by showing that Fusarium graminearum can take up externally applied dsRNAs. Spraying the fungus with artificial long dsRNAs led to their processing by FgDCL1 into three sRNAs. These sRNAs targeted key genes in ergosterol biosynthesis and effectively suppressed infection in barley[30]. Similarly, topical application of sRNAs or dsRNAs targeting Botrytis cinerea DCL1 and DCL2 genes significantly reduced disease symptoms in fruits, vegetables, and flowers[63]. Foliar spraying of dsRNAs also provided strong protection against Sclerotinia sclerotiorum in multiple crops[64]. These findings confirmed spray-induced gene silencing (SIGS) as a promising method for fungal disease control in both pre- and post-harvest stages (Fig. 3)[60]. SIGS offers key advantages. It allows precise targeting of virulence genes and minimizes off-target effects. It also avoids the time and cost of producing transgenic plants.
Further investigations revealed that dsRNA uptake efficiency varies considerably across fungal species. Most pathogenic fungi showed high uptake capacity, while beneficial fungi like Trichoderma virens absorbed dsRNA poorly[65]. To enhance RNA stability in the environment, SIGS has adopted nanovesicles—also called artificial vesicles (AVs)—to encapsulate RNA. This approach improves RNA durability and increases disease control effectiveness[34]. However, key challenges remain. The high production cost of RNA-loaded AVs and the potential ecological risks must be addressed before SIGS can be widely applied in the field.
The three main RNAi-based methods for cross-kingdom gene silencing have been described. Although the principle of CRISPR (Clustered Regularly Interspersed Short Palindromic Repeats) based gene editing does not involve sRNA delivery, it still shares some similarities with HIGS, SIGS, and MIGS. Therefore, key features of these approaches are compared and summarized in Table 1.
Table 1. The comparison between trans-kingdom RNAi technology and CRISPR-based technology.
Name Plant transgene dependent Target type Off-target effect sRNA delivery Application stage Cultivation time CRISPR-based Yes DNA/RNA Medium No Breeding Long HIGS Yes mRNA Low Yes Breeding Long SIGS No mRNA Low Yes Growth Very short MIGS No mRNA Low Yes Growth Short -
The demand for environmentally sustainable and safe biocontrol strategies is growing rapidly in modern agriculture. sRNAs and dsRNAs are now recognized as promising biocontrol agents. Strategies such as HIGS, SIGS, and MIGS have been developed based on different sRNA delivery mechanisms. However, the success of these methods depends on several factors. These include RNA uptake efficiency, transport, and molecular stability within the host–pathogen system. All of these processes require further investigation through interdisciplinary research.
Cost is another major challenge for the large-scale application of HIGS, SIGS, and MIGS. In addition, the presence of suppressors of trans-kingdom RNAi (STRs), which have evolved during plant–pathogen coevolution, reduces the effectiveness of RNAi-based strategies. Targeting STR-encoding genes, together with essential virulence genes, may enhance RNAi efficiency. However, this approach depends on the identification of STRs in different phytopathogens.
Moreover, studies have shown that HIGS triggers only a limited and non-amplified RNAi response in pathogen cells[66], suggesting the presence of STRs. Whether plants also produce STR-like proteins to counter pathogen interference remains unknown. Discovering such proteins could help overcome current limitations of RNA-based disease control.
Further research is needed to clarify the molecular properties, delivery vehicles, mobility, and lifespan of RNA molecules during plant–pathogen interactions. These insights will be critical for improving the effectiveness of RNAi-based plant protection strategies.
-
Not applicable.
This research was funded by the Natural Science Research Project of Anhui Educational Committee (2022AH030026), the Scientific Research Foundation for High-level Talents of Anhui Normal University (762208), and The University Synergy Innovation Program of Anhui Province (GXXT-2022-067). The illustrations used in this manuscript were created using BioRender.com.
-
The authors confirm their contributions to the paper as follows: study conception and design: Duan CG; draft manuscript preparation: Luo M, Zhu C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Luo M, Zhu C, Duan CG. 2025. Trans-kingdom RNAi in plant–pathogen interactions: mechanisms and biocontrol strategies. Epigenetics Insights 18: e011 doi: 10.48130/epi-0025-0010 

Trans-kingdom RNAi in plant–pathogen interactions: mechanisms and biocontrol strategies
- Received: 01 May 2025
- Revised: 04 July 2025
- Accepted: 23 July 2025
- Published online: 04 August 2025
Abstract: The exchange of secondary metabolites or proteins between plants and phytopathogens has long been a central topic in plant–pathogen interaction. In recent decades, small RNAs (sRNAs) have emerged as key effectors during plant–pathogen interaction. RNA interference (RNAi) induced by foreign sRNA across species boundaries, known as trans-kingdom RNAi, plays crucial roles in regulating both plant defense and pathogen virulence. Beyond its biological significance, trans-kingdom RNAi is gaining attention as a promising strategy for sustainable crop disease management through RNAi-based biocontrol. In this review, we summarize recent advances in understanding the mechanisms of trans-kingdom RNAi and three major RNAi-based technologies for pathogen control, along with their practical applications. We also discuss the definition and functional mechanism of suppressor of trans-kingdom RNAi (STR) proteins. A deeper understanding of these mechanisms holds great potential for developing innovative approaches to crop protection.
-
Key words:
- Trans-kingdom RNAi /
- RNAi suppressor /
- HIGS /
- SIGS /
- MIGS /
- EVs