









-
As living standards improve and health awareness increases, the functional needs of traditional ornamental plants are changing. Modern consumer preferences have gradually shifted from purely ornamental plants to multifunctional ones, especially those with medicinal or edible properties. These plants not only contribute to environmental aesthetics but also possess significant practical value due to their medicinal and edible functions. For instance, chrysanthemums, roses, peonies, and lilies are rich in polyphenolic compounds, which have numerous health benefits and are widely used in food, medicine, cosmetics, and healthcare[1,2].
In line with this trend, Gesneriaceae plants attract wide attention due to their unique ornamental characteristics, extensive species diversity, and profound medicinal history. Gesneriaceae is a large family of plants, including about 163 genera and more than 4,000 species[3]. China is the center of origin and a major distribution area for the family, boasting a wealth of wild germplasm resources. All species of this family in China belong to the Cyrtandroideae subfamily, comprising 45 genera and 799 species, which are widely distributed from southern Tibet and Yunnan to Hebei and southwestern Liaoning[4]. As a traditional ethnomedicine with a long history in China, the medicinal value of Gesneriaceae plants has been confirmed through extensive use over time, and they hold an important place in traditional folk medicine. Notably, Lysionotus pauciflorus is included in the Chinese Pharmacopoeia, where it is recognized for its ability to resolve phlegm, relieve cough, soften hardness, and disperse nodules[5]. Moreover, studies indicate that the medicinal properties of this family are not confined to China; in the traditional medical systems of other Asian countries and South America, L. pauciflorus is extensively used to treat various inflammatory conditions, gastrointestinal disorders, and snake and insect bites[6−10]. This trans-geographical medicinal practice further validates the scientific basis and universal potential of its biological activity and medicinal value.
Primulina is the genus with the largest number of species and the richest species diversity in Cyrtandroideae[11]. The plants in this genus exhibit a wide range of flower colors, distinctive leaf patterns, and strong environmental adaptability, making many species highly desirable as emerging ornamental floral resources[12]. Additionally, recent studies indicate that the leaves of some Primulina species are rich in soluble calcium[13], a finding that presents new possibilities for their use in the development of high-calcium vegetables, thereby further expanding the application potential of Gesneriaceae.
Although Gesneriaceae demonstrate great potential for ornamental, medicinal, and edible uses, few systematic studies have explored the chemical constituents of their leaves. Recent chemical composition studies reveal that Gesneriaceae leaves primarily contain flavonoids, phenolic acids, quinones, terpenoids, phenylethanol glycosides, and other compounds[14−17]. On this basis, ten Gesneriaceae taxa were selected to comprehensively explore their chemical diversity and potential value. These included L. pauciflorus, the only species in the Gesneriaceae family listed in the Chinese Pharmacopoeia; three wild species (Primulina eburnea, Primulina fimbrisepala, Primulina medica) with a long history of use in folk medicine for treating respiratory diseases and inflammation, representing important genetic resources for medicinal plant research; and six hybrid cultivars (Primulina 'Nandia Dreams', Primulina 'Venus', Primulina 'The Smurfs', Primulina 'Spring of Beilin', Primulina 'Purple Sundae', Primulina 'Four Seasons') derived from crosses involving one of the three wild species, selected for their desirable ornamental traits to explore their multifunctional potential. Both untargeted and targeted metabolomics techniques were employed in this analysis. To further investigate the functional properties of these chemical constituents, the study also offered a comprehensive assessment of their antioxidant activities and explored potential correlations between major differential metabolites and bioactive components. This study aims to offer new scientific insights into the chemical composition and antioxidant activity of Gesneriaceae leaves and to serve as a valuable preliminary reference for exploring the diverse medicinal and edible applications of these plants.
-
All wild species used in this study (L. pauciflorus, P. eburnea, P. fimbrisepala, and P. medica) were originally collected from natural habitats in Guangxi, China, and authenticated by Dr. Fang Wen (Guangxi Institute of Botany, Chinese Academy of Sciences). These accessions were introduced to the greenhouse at Beijing Forestry University in 2007 and propagated clonally through leaf cuttings. Six hybrid cultivars were artificially developed with the following parental combinations: P. 'Nandia Dreams' (Primulina pungentisepala × P. fimbrisepala), P. 'Venus' (P. pungentisepala × P. eburnea), P. 'The Smurfs' (P. fimbrisepala × Primulina linearifolia), P. 'Spring of Beilin' (P. fimbrisepala × P. linearifolia), P. 'Purple Sundae' (P. eburnea × Primulina longii), and P. 'Four Seasons' (a bud mutation of Primulina ronganensis). All six new cultivars have been registered with The Gesneriad Society (International Cultivar Registration Authority for Gesneriaceae) with internationally certified names. Several cultivars also hold Plant Variety Rights from the National Forestry and Grassland Administration of China.
All plant materials (Fig. 1) were cultivated in the greenhouse at Daxing Flower Base, Beijing, China. For each taxon, five healthy and uniform individuals were selected, and mature leaves were collected in November 2023. All samples were immediately frozen in liquid nitrogen. The frozen samples were then stored at −80 °C to preserve their integrity for subsequent analysis and investigation.

Figure 1.
The ten taxa of Gesneriaceae used in this study. (a) L. pauciflorus, (b) P. eburnea, (c) P. fimbrisepala, (d) P. medica, (e) P. 'Nandia Dreams', (f) P. 'Venus', (g) P. 'The Smurfs', (h) P. 'Spring of Beilin', (i) P. 'Purple Sundae', (j) P. 'Four Seasons'.
Metabolome analysis
-
The samples were ground using liquid nitrogen. Subsequently, 1.0 g of the sample powder was extracted with 5.0 mL of 80% aqueous methanol for 30 min in an ultrasonic ice bath. The extraction was then moved to 4 °C and allowed to continue for 10 h. After that, the extract was centrifuged at 13,500 g for 12 min, and the supernatant was taken and filtered through a 0.22 μm Millipore filter. The prepared samples were stored at −80 °C for further analysis. The Agilent UHPLC 1290II system coupled to a G6500 quadrupole time-of-flight mass spectrometer (Q-TOF, Agilent Technologies, Santa Clara, CA, USA) was performed in positive and negative ion modes. The Q-TOF parameters were as follows: sheath gas temperature 350 °C; gas temperature 300 °C; gas flow rate 7 L/min; nozzle voltage 1,500 V; nebulizer pressure 35 psi; vcap 3000 V; and fragmentor 120 V. The Agilent UHPLC 1290II system coupled to a G6400 triple quadrupole mass spectrometer (QQQ, Agilent Technologies, Santa Clara, CA, USA) was utilized to produce product ion (PI) scans and multiple reaction monitoring (MRM). The QQQ parameters were as follows: sheath gas temperature 250 °C; gas temperature 300 °C; gas flow rate 11 L/min; nozzle voltage 500 V; nebulizer pressure 45 psi; vcap 3500 V; and fragmentor 120 V. The liquid chromatographic conditions were as follows: Agilent Eclipse Plus C18 column (2.1 mm × 150 mm, 2.7μm) (Agilent Technologies, Santa Clara, CA, USA); column temperature 35 °C; mobile phase constituting of water containing 0.05% acetic acid, 5 mM ammonium acetate (A), and acetonitrile (B); flow rate 0.3 mL/min; and sample injection volume 3 μL. The solvent gradient was set as follows: 0–2 min: 5% B, 2.5 min: 18.5% B, 10.5 min: 41% B, 11 min: 59% B, 18 min: 77% B, 22 min: 95% B, 24 min: 95% B, 24.1 min: 5% B, 26 min: 5% B. The MRM for each sample was performed in triplicate. Additionally, quality control (QC) samples were prepared by pooling equal volumes of all experimental samples and analyzed at regular intervals throughout the metabolomic analysis. Differentially accumulating metabolites (DAMs) were screened using thresholds (|log2fold change (FC)| > 1.5 or < 0.67, p-value (p) < 0.05, and variable importance projection (VIP) > 1).
Determination of total phenolic and total flavonoid content
-
The total phenolic content (TPC) was determined using a modified Folin-Ciocalteu reduction (FCR) method[18]. First, 100 μL of supernatant was accurately pipetted, and 100 μL of 0.25 mol/L Folin-Ciocalteu reagent was added, mixed thoroughly, and left to react for 30 min. Then, 200 μL of 15% aqueous sodium carbonate solution was added, mixed thoroughly, and left for another 30 min, protected from light. After the color development stabilized, the mixture was centrifuged at 3,500 rpm for 3 min, and the absorbance was measured at 760 nm. The standard curve was plotted based on gallic acid concentration and absorbance at 760 nm. Each group was tested in triplicate. The TPC (gallic acid) was calculated from the standard curve using the equation y = 0.0167x + 0.0486, R2 = 0.9972, where 'y' represents absorbance and 'x' represents gallic acid concentration. Results were expressed as gallic acid equivalents (GAE) per gram of fresh weight (mg GAE/g FW).
The total flavonoid content (TFC) was determined using the aluminum chloride colorimetric method[19]. First, 60 μL of the sample solution was accurately pipetted, followed by the addition of 30 μL of 5% sodium nitrite solution. The mixture was shaken thoroughly and left to stand for 6 min. Next, 30 μL of 10% aluminum nitrate solution was added, shaken thoroughly, and left to stand for another 6 min. Finally, 400 μL of 4% sodium hydroxide solution was added, mixed thoroughly, and left to stand for 15 min, protected from light. The absorbance of the sample solution was measured at 510 nm, and the standard curve was plotted based on rutin concentration and absorbance. Each group was tested in triplicate. The TFC (rutin) was calculated using the standard curve with the equation: y = 0.7308x + 0.0546, R2 = 0.9935, where 'y' represents absorbance and 'x' represents rutin concentration. The TFC was expressed as mg of rutin equivalents per gram of fresh weight (mg RE/g FW).
Determination of antioxidant activity
-
2,2-Diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging assay. The DPPH radical scavenging assay was performed according to a previously described method[20] with slight modifications. A total of 0.4 mL of the sample and 1.8 mL of 100 μmol/L DPPH solution were added to test tubes, left to stand for 30 min away from light, and the absorbance was measured at 517 nm. The DPPH radical scavenging ability of the samples was calculated using the standard curve equation y = 0.5426x − 1.2917, R2 = 0.9932, where 'y' represents the DPPH scavenging rate and 'x' represents the Trolox concentration. The DPPH scavenging capacity was expressed as milligrams of Trolox equivalents (TE) per gram of fresh weight (mg TE/g FW).
Determination of ABTS radical scavenging activity[21]. In a 96-well plate, 10 μL of the sample solution was added to each well, followed by 100 μL of ABTS working solution. The reaction was conducted at 30°C for 6 min, after which the absorbance was measured at 734 nm. The standard curve equation was y = 0.7189x + 0.0489, R2 = 0.999, where 'y' represents absorbance and 'x' represents the Trolox concentration. ABTS radical scavenging activity was expressed as milligrams of Trolox equivalents per gram of fresh weight (mg TE/g FW).
Cupric reducing antioxidant capacity (CUPRAC) assay. Following previous research[22], 0.1 mL of the sample extract was pipetted, and 0.5 mL of 5 mmol/L CuSO4, 0.5 mL of 3.75 mmol/L neocuproine, 1 mL of 1 mol/L ammonium acetate, and 1 mL of water were added sequentially, mixed thoroughly, and then allowed to react in the dark for 30 min. The absorbance was then measured at 450 nm. The standard curve equation was y = 0.4521x + 0.0383, R2 = 0.9987, where 'y' represents absorbance and 'x' represents the Trolox concentration. CUPRAC activity was expressed as milligrams of Trolox equivalents per gram of fresh weight (mg TE/g FW).
Determination of ferric ion reducing antioxidant power (FRAP)[23]: 1 mL of the sample extract was pipetted into a small test tube, 4.5 mL of TPTZ working solution was added, vortexed, and then allowed to react in the dark at 25 °C for 30 min. The absorbance was measured at 593 nm. The standard curve equation was y = 0.007x + 0.1376, R2 = 0.997, where 'y' represents absorbance and 'x' represents the Trolox concentration. Iron reducing capacity was expressed as milligrams of Trolox equivalents per gram of fresh weight (mg TE/g FW).
Comprehensive evaluation of antioxidant activity
-
The antioxidant potential of the ten samples was comprehensively evaluated using the weighted membership function method. First, each indicator was standardized according to the following formula Eqn (1):
$ \mu ({X}_{j})=\dfrac{{X}_{j}-{X}_{min}}{{X}_{max}-{X}_{min}} $ (1) Where j = 1,2,3, …, n; Xj represents the value of the j-th indicator; Xmin is the minimum value of the j-th indicator; and Xmax is the maximum value of the j-th indicator.
The standard deviation coefficient (Vj), weight coefficient (Wj), and weighted membership function value (D) were calculated according to Eqns (2)−(4). A larger D value indicates stronger antioxidant activity.
$ {V}_{j}=\dfrac{\sqrt{\dfrac{\sum _{i=1}^{n}{\left({X}_{ij}-\overline{{X}_{j}}\right)}^{2}}{n-1}}}{{\overline{X}}_{j}} $ (2) $ {W}_{j}=\dfrac{{V}_{j}}{\sum _{j=1}^{n}{V}_{j}} $ (3) $ D=\sum _{j=1}^{n}\left[\mu ({X}_{j})*{W}_{j}\right]$ (4) Where
$ \overline{{X}_{j}} $ Statistical analysis
-
One-way analysis of variance (ANOVA) was conducted using IBM SPSS 26. Histograms were created using GraphPad Prism 9.5 software. Correlation and clustering heat map analysis was conducted using the ChiPlot website (
www.chiplot.online ). Principal component analysis (PCA) and orthogonal partial least squares-discriminant analysis (OPLS-DA) were conducted using SIMCA 14.1 software. -
By querying and summarizing information about various chemical components of Gesneriaceae from CNKI and SciFinder, a first-order mass spectrometry database of Gesneriaceae components was established. The database includes key information such as compound names, molecular formulas, and relative molecular masses, providing critical data support for subsequent compound identification. Subsequently, raw UPLC-ESI-MS/MS data were processed, and compounds were extracted based on the self-constructed database and public mass spectrometry databases, a total of 48 metabolites were tentatively identified in the leaf samples of ten Gesneriaceae taxa. These metabolites included flavonoids (10), phenolic acids (2), terpenoids (4), lipids (15), phenylethanoid glycosides (7), quinones (1), coumarins (2), saccharides (4), alkaloids (1), and others (2) (Supplementary Tables S1, S2). Three compounds were definitively identified by comparison with standards, while the structures of the remaining compounds were tentatively assigned based on their primary and secondary mass spectra, fragmentation patterns, and literature references.
By mass spectrometry analysis, ten flavonoids were tentatively identified. These were primarily characterized by polyhydroxyl and polymethoxy structures, and sugar groups were mainly lost during mass spectrometry fragmentation, showing a unique aglycone structure. Compounds 1 and 5 were tentatively identified as 5,7-dihydroxy-8-C-β-D-glucosyl-chromone and phylloflavanine, with parent ions ([M-H]−) observed at m/z 339.0722 and m/z 659.1738, respectively, and molecular formulas presumed to be C15H16O9 and C35H32O13. Major fragment ions were observed at m/z 177.0193 and m/z 461.1662, possibly resulting from the loss of the glucosyl group from the parent ion. Compounds 2, 3, 4, 6, and 7 were tentatively identified in negative ion mode as 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone (m/z 667.188), 6'-methoxypseudobaptigenin 7-O-glucoside (m/z 473.1088), hesperetin 7-(2,6-dirhamnosylglucoside) (m/z 755.2401), tetramethylquercetin 3-rutinoside (m/z 665.2087), and lethedoside B (m/z 519.1503). The major fragment ions observed were m/z 505.1345, m/z 293.0462, m/z 593.2087, m/z 461.1672, and m/z 357.1171, which are likely due to the loss of a dehydrated glucose molecule from the parent ion. Compounds 2 and 6 further lost one hexose, yielding m/z 341.0870 ([M-H-CH3-C6H10O5-C6H12O5]−), m/z 315.1095 ([M-H-CH3-C6H10O5-C6H10O4]−) separately; Compound 4 continued to lose two hexose, resulting in m/z 447.1510 ([M-H-CH3-C6H10O5-C6H10O4]−) and m/z 315.1082 ([M-H-CH3-C6H10O5-C6H10O4-C5H8O4]−). Compounds 8 and 9 were tentatively identified as hispidulin and 5,7-dihydroxy-6,4'-dimethoxyflavone, with parent ions at m/z 299.0561 ([M-H]−) and m/z 315.0863 ([M+H]+), respectively, and presumed molecular formulas C16H12O6 and C17H14O6. The major fragment ions were m/z 284.0326 and m/z 300.0630, resulting from the loss of a methyl group from the parent ion. Hispidulin, on the other hand, yielded a fragment ion at m/z 184.0527 ([M-H-CH3-C5H8O2]−), suggesting that the parent ion lost a methyl group followed by an acyl group (C5H8O2). Based on a literature search, compounds 8 and 9 have been reported in Aeschynanthus superbus[24] and Corallodiscus kingianus[25], respectively. Compound 10 was identified as 5,7-dihydroxy-6,8,4'-trimethoxy flavone through standard comparison. Ten flavonoids were tentatively identified using the aforementioned identification method.
Phenolic acids are more responsive in negative ion mode and tend to lose H2O and -COOH during secondary fragmentation. For example, compound 11 has a parent ion ([M-H]−) at m/z 193.0506, with a presumed molecular formula of C10H10O4. The fragmentation ion at m/z 161.0238 ([M-H-H2O-CH3]−) corresponds to the removal of a methyl group, and the subsequent fragmentation to form m/z 133.0293 ([M-H- CH3-COOH]−) corresponds to the further loss of the carboxylic acid group. Based on a search of the MASSBANK database, compound 11 was tentatively identified as isoferulic acid, which has been reported in Aeschynanthus bracteatus[14]. Compound 12 was tentatively identified as [7]-paradol, with a parent ion ([M-H]−) at m/z 291.1966 and a presumed molecular formula of C18H28O3. The fragment ion at m/z 247.2068 ([M-H-CO2]−) results from the loss of one CO2 from the parent ion, followed by the loss of an acyl group and a molecule of H2O, yielding m/z 165.1291. Using the aforementioned identification method, two phenolic acids were tentatively identified.
Phenylethanol glycosides contain multiple functional groups, such as glucose and rhamnose, which are easy to combine with sugar components to form an aglycone structure. In secondary fragmentation, it is also easy to lose glucose residues, -OH and-COOH structures, generating numerous fragment ions. Compound 32 was tentatively identified as beta-hydroxyacteoside, with a parent ion ([M-H]−) at m/z 639.1931 and a presumed molecular formula of C29H36O16. The major fragment ions produced were m/z 477.1400 and m/z 315.1085, which were likely derived from the parent ion by the loss of two dehydrated glucose molecules, consistent with previous reports on olive[26]. Compound 33 was tentatively identified as phlinoside A. The parent ion ([M-H]−) was observed at m/z 785.2508, with a presumed molecular formula of C35H46O20. The fragmentation ion at m/z 753.2233 may result from the loss of a methoxy group (-OCH3) from the parent ion, followed by the subsequent loss of a dehydrated glucose molecule, resulting in m/z 591.1922 ([M-H-OCH3-C6H10O5]−). Fragment ion m/z 427.1212 ([M-H- OCH3- C6H10O5- C6H12O5]−) may correspond to the further loss of a rhamnose unit. Compound 34 was tentatively identified as plantamajoside, which has been previously reported in P. eburnea[27]. The parent ion ([M-H]−) was observed at m/z 639.1931, with a presumed molecular formula of C29H36O16. The fragmentation ion at m/z 607.1647 may result from the loss of a methoxy group (-OCH3) from the parent ion, followed by the subsequent loss of a dehydroglucose molecule and a glucuronide unit, yielding m/z 445.1348 ([M-H- C6H10O5]−) and m/z 251.0546 ([M-H- C6H10O5- C6H10O7]−). Furthermore, a possible further loss of CO, CO2, and H2O results in m/z 161.0244. Compound 35 was tentatively identified as isonuomioside A, with a parent ion ([M-H]−) at m/z 609.1825 and a presumed molecular formula of C28H34O15. The fragment ion at m/z 447.1515 ([M-H- C6H10O5]−) corresponds to the loss of a molecule of dehydrated glucose from the parent ion, followed by another molecule of sugar to give m/z 315.1092([M-H- C6H10O5- C5H8O4]−). Compounds 36 and 37 were identified by standard comparison as isoacteoside and calceolarioside B, respectively. Compound 38 was tentatively identified as aeschynanthoside B, with a parent ion ([M-H]−) at m/z 665.2087 and a presumed molecular formula of C31H38O16. The fragment ion at m/z 503.1769 ([M-H- C6H10O5]−) results from the loss of a molecule of dehydrated glucose from the parent ion, followed by the loss of another molecule of dehydrated rhamnose, giving m/z 357.1133 ([M-H- C6H10O5- C6H10O4]−). According to the above identification method, seven phenylethanol glycosides were tentatively identified.
The mass spectrometry conditions were optimized on the basis of tentative qualitative results. Full-scan mode was used for 48 compounds in both positive and negative ion modes to optimize mass spectral conditions. The results indicated that certain compounds exhibited superior sensitivity in both ion modes, and MRM conditions were optimized for the 48 compounds based on the highest sensitivity and specific ion pairs (Supplementary Table S3). Relative contents of the target metabolites were determined using a targeted metabolomics approach on the LC-QQQ-MS system. Quantitative results revealed significant differences in the metabolite distribution patterns among the leaves of different Gesneriaceae taxa. Hierarchical cluster analysis (HCA) was conducted to summarize the distribution patterns. Based on the HCA results (Fig. 2), the biological replicates for each taxa clustered closely together, indicating high reproducibility. Additionally, the QC samples showed a good fit, as evidenced by the high degree of overlap of the total ion chromatograms (TICs) (Supplementary Fig. S1). This indicates minimal instrumental variability throughout the experiment. The ten taxa were categorized into two groups. P. medica, P. 'Purple Sundae', and P. 'Four Seasons', L. pauciflorus formed Group 1, while P. fimbrisepala, P. 'Spring of Beilin', P. 'The Smurfs', P. eburnea, P. 'Nandia Dreams', and P. 'Venus' formed Group 2. The PCA and OPLS-DA analysis were conducted (Fig. 3a & d), yielding an explanatory R2X of 0.424 for the X variable, an explanatory R2Y of 0.992 for the Y variable, and a predictability Q2 of 0.979. As assessed by the permutation test (Fig. 3b), the model was not overfitted and was credible enough to be used in the subsequent analysis of metabolite differential analyses among the ten taxa.
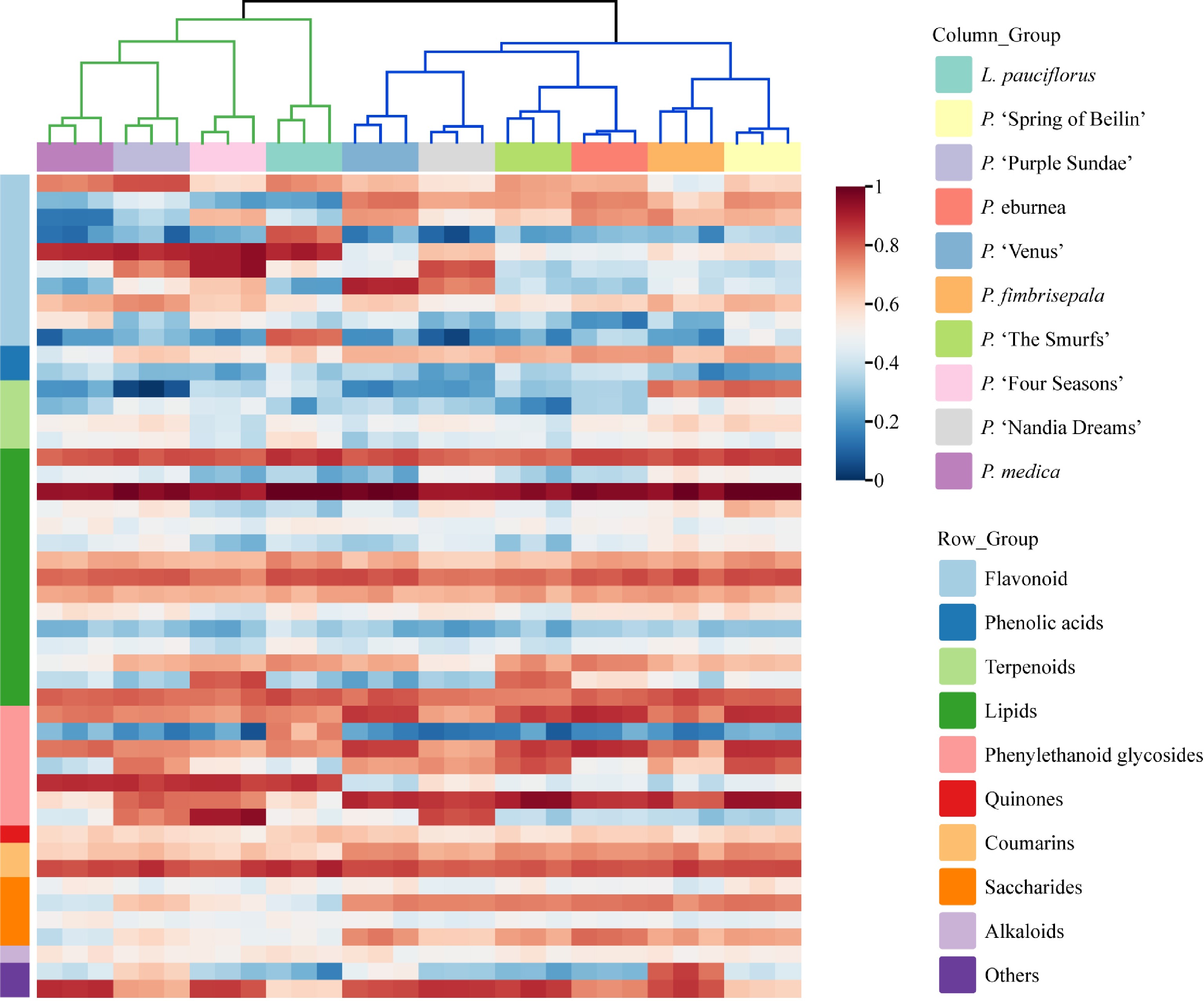
Figure 2.
Heat map analysis of metabolite clustering in the leaves of ten Gesneriaceae taxa.
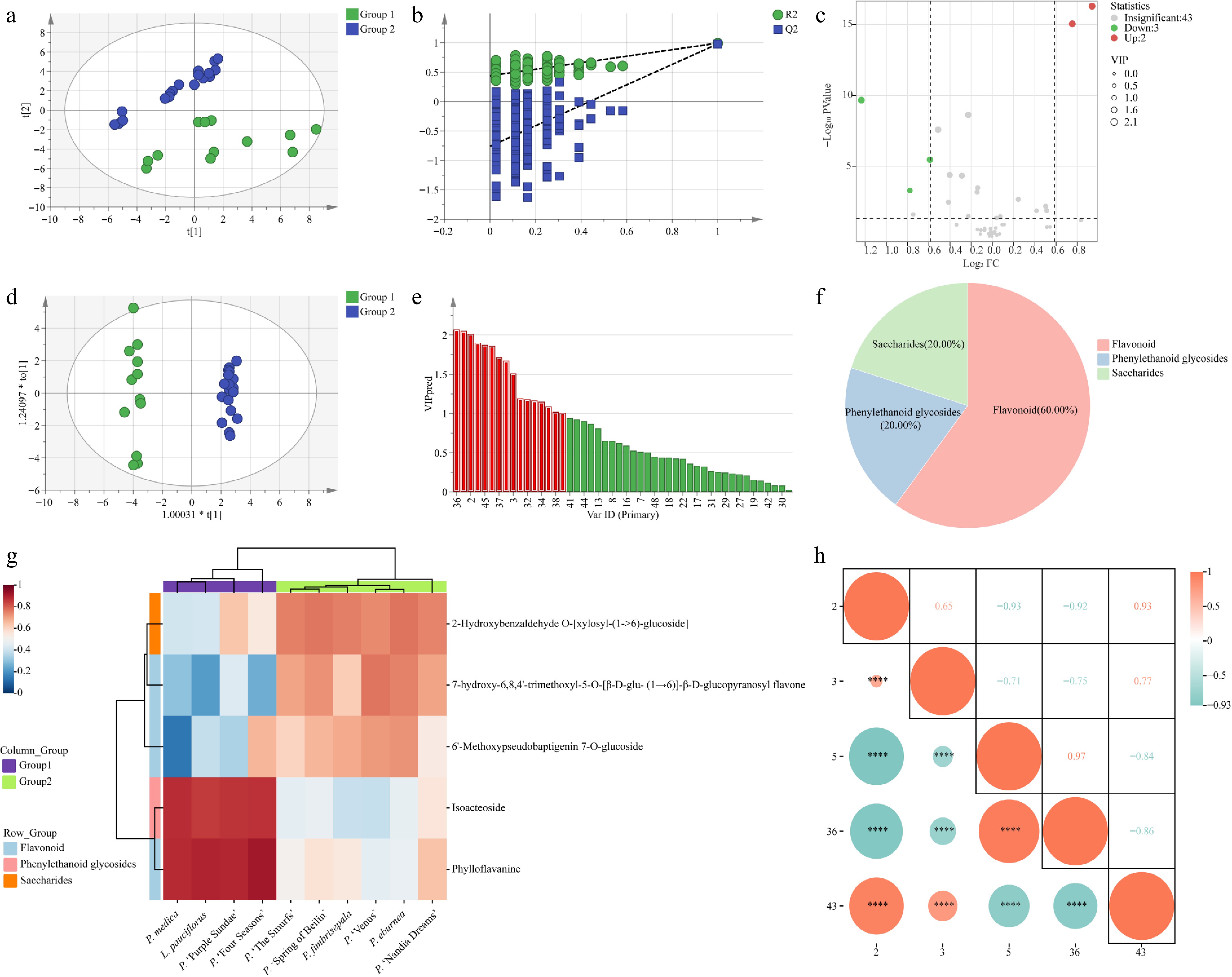
Figure 3.
Multivariate statistical analyses of metabolites in the leaves of ten Gesneriaceae taxa. (a) Scatter plot of PCA scores. (b) Model replacement test plot. (c) Volcano diagram. (d) Scatter plot of OPLS-DA scores. (e) VIP values of metabolites. (f) DAMs pie chart. (g) DAMs clustering heat map. Group 1: P. medica, P. 'Purple Sundae,' P. 'Four Seasons', L. pauciflorus; Group 2: P. fimbrisepala, P. 'Spring of Beilin', P. 'The Smurfs', P. eburnea, P. 'Nandia Dreams', and P. 'Venus'. (h) DAMs correlation analysis. 2: 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone; 3: 6'-Methoxypseudobaptigenin 7-O-glucoside; 5: Phylloflavanine; 36: Isoacteoside; 43: 2-Hydroxybenzaldehyde O-[xylosyl-(1→6)-glucoside].
VIP > 1, p < 0.05, and FC > 1.5 or < 0.67 were set as the screening criteria. A total of five DAMs were identified through VIP and FC screening, with two upregulated and three down-regulated. These DAMs consisted of three (60.00%) flavonoids, 1 (20.00%) phenylethanol glycoside, and 1 (20.00%) saccharide (Fig. 3c, e−f; Supplementary Table S4). Through clustering heatmap analysis, distinct metabolite expression patterns across different species were observed (Fig. 3g). Phylloflavanine and isoacteoside showed significant upregulation in Group 1. However, phylloflavanine showed some expression in 'Nandia Dreams' from Group 2. This result suggests that, although these metabolites are significantly upregulated in Group 1, they may still play certain biological roles in other species. 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone, 6'-Methoxypseudobaptigenin 7-O-glucoside, and 2-Hydroxybenzaldehyde O-[xylosyl-(1→6)-glucoside] were significantly downregulated in Group 1. However, 2-Hydroxybenzaldehyde O-[xylosyl-(1→6)-glucoside] still showed some expression in 'Purple Sundae' in Group 1, and 6'-Methoxypseudobaptigenin 7-O-glucoside was expressed at high levels in 'Four Seasons'. To further explore the potential connections between the differential metabolites, correlation analysis was performed (Fig. 3h). The results showed that the upregulated differential metabolites were significantly positively correlated, with a correlation coefficient of 0.97, suggesting that they might be involved in the same metabolic pathways or biological processes. The downregulated metabolites also showed significant positive correlations, with a correlation coefficient of 0.93 between 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone and 2-Hydroxybenzaldehyde O-[xylosyl-(1→6)-glucoside], indicating potential synergistic roles in metabolic regulation. Moreover, the analysis revealed that 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone was significantly negatively correlated with both phylloflavanine and isoacteoside, with correlation coefficients of −0.93 and −0.92, respectively. This negative correlation likely reflects their antagonistic roles in the metabolic network, suggesting an interaction across metabolic pathways.
Determination of bioactive components and antioxidant activity
Determination of total phenolic and total flavonoid content
-
L. pauciflorus exhibited the highest TPC and TFC, which were significantly different (p < 0.05) from those of other Primulina taxa. The TPC of P. 'Spring of Beilin' and P. 'Purple Sundae' was significantly higher than that of P. medica (p < 0.05), and both taxa also had relatively high TFC values. The TPC of P. fimbrisepala, P. 'The Smurfs', and P. 'Four Seasons' was lower, with no significant differences (p > 0.05), while P. 'Venus' exhibited the lowest TPC (Fig. 4). Significant differences in TFC were observed among different genera of Gesneriaceae, which corresponded to the differences in TPC.
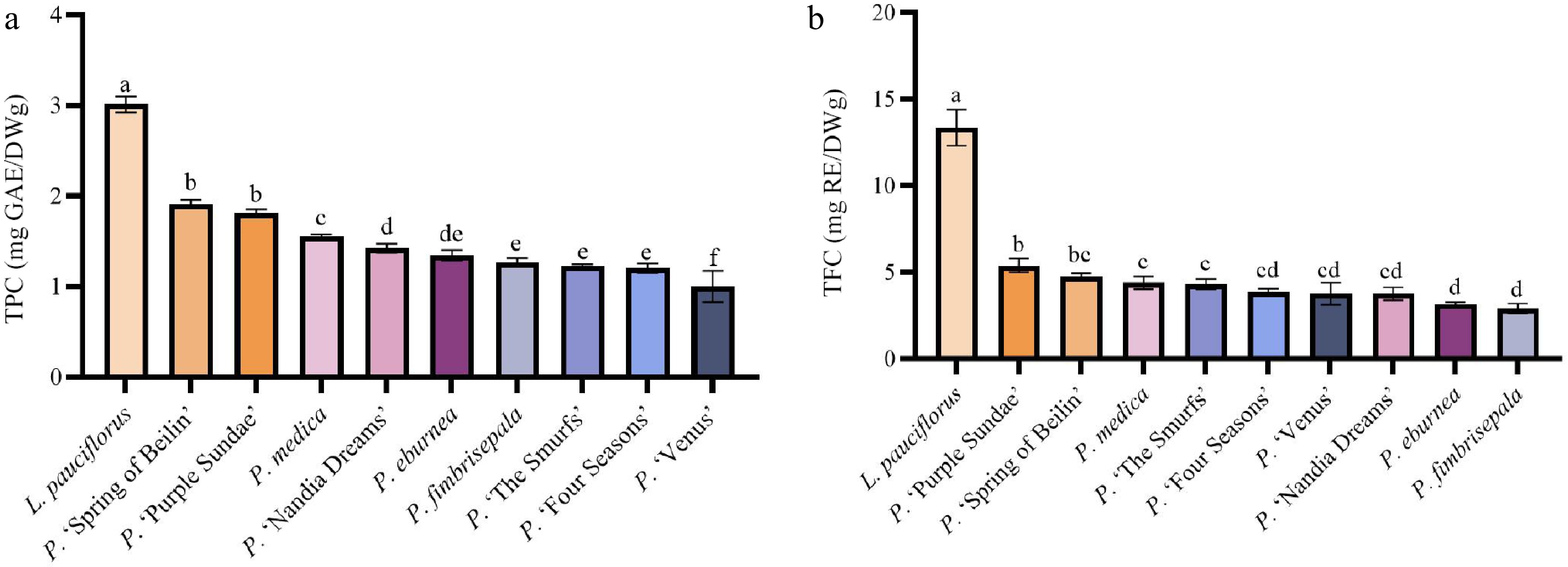
Figure 4.
Bioactive components in the leaves of ten Gesneriaceae taxa. (a) Total phenolic content (TPC) results. (b) Total flavonoid content (TFC) results. Data with no or the same letter indicate no significant difference at p > 0.05; those with different letters indicate a significant difference at p < 0.05.
Determination of antioxidant activity in the leaves of Gesneriaceae using four methods
-
The antioxidant capacity of leaves from different taxa was evaluated using four methods: DPPH, ABTS, CUPRAC, and FRAP (Fig. 5).
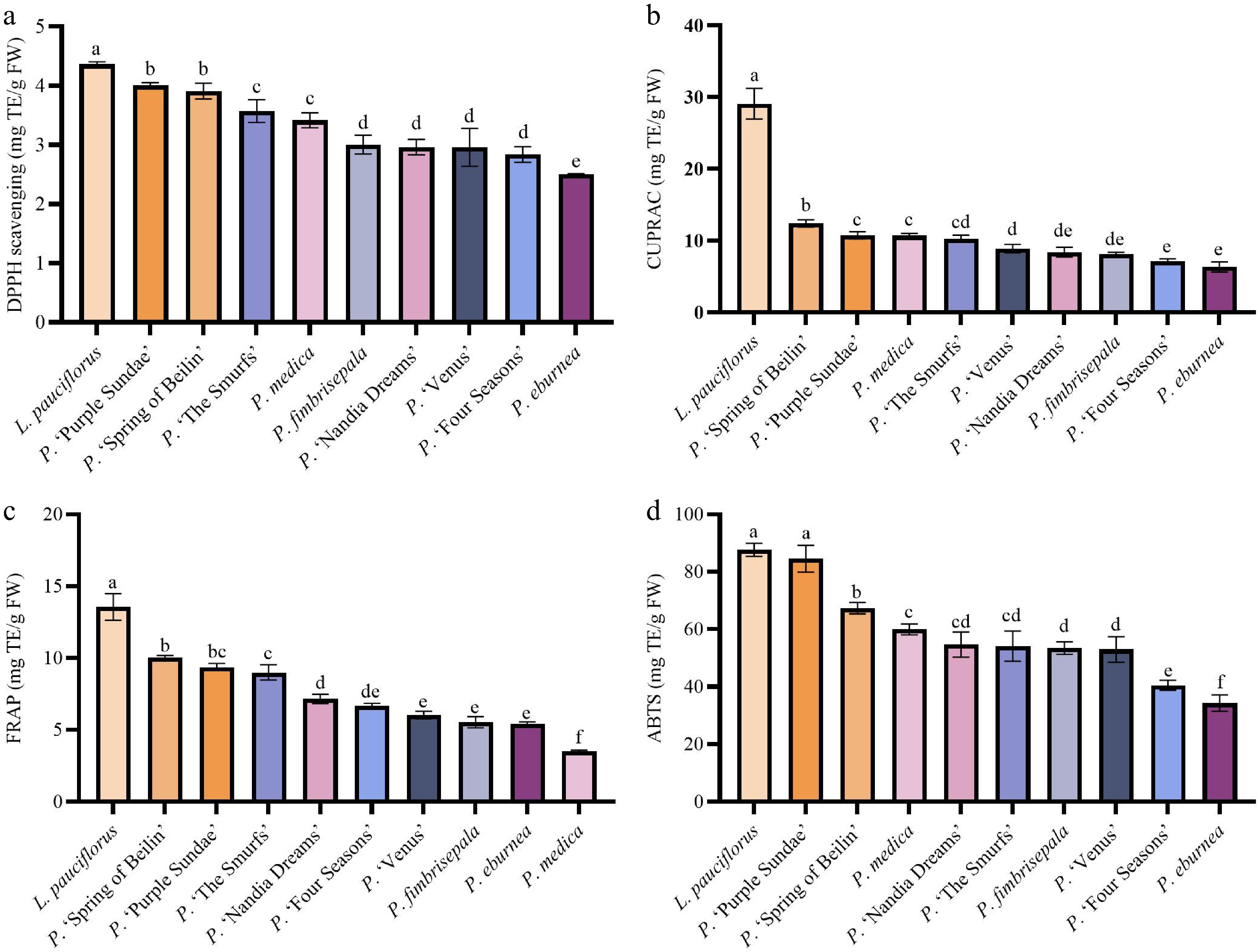
Figure 5.
Bioactive components in the leaves of ten Gesneriaceae taxa. (a) DPPH results. (b) CUPRAC results. (c) FRAP results. (d) ABTS results. Data with no or the same letter indicate no significant difference at p > 0.05; those with different letters indicate a significant difference at p < 0.05.
All results were expressed as Trolox equivalents (mg TE/g FW), revealing significant differences in the antioxidant capacity of leaves across different taxa, which reflects the diversity of their antioxidant potential.
DPPH scavenging activity ranged from 2.50 to 4.36 mg/g. L. pauciflorus exhibited the strongest antioxidant activity at 4.36 mg/g, significantly higher than that of other samples (p < 0.05), followed by P. 'Purple Sundae' (4.01 mg/g) and P. 'Spring of Beilin' (3.91 mg/g). The lowest antioxidant capacity was found in P. eburnea at 2.5 mg/g.
ABTS scavenging activity ranged from 34.27 to 87.65 mg/g. L. pauciflorus and P. 'Purple Sundae' exhibited stronger antioxidant activity, at 87.65 mg/g and 84.52 mg/g, respectively, significantly higher than other samples (p < 0.05). This was followed by P. 'Spring of Beilin' at 67.35 mg/g, while P. eburnea showed the lowest activity at 34.27 mg/g.
CUPRAC reduction activity ranged from 6.35 to 29.09 mg/g. L. pauciflorus exhibited the strongest antioxidant activity at 29.09 mg/g, significantly higher than other samples (p < 0.05). This was followed by P. 'Spring of Beilin' at 12.40 mg/g, approximately 50% of L. pauciflorus' antioxidant activity. The remaining samples showed little variation in activity, ranging from 6.35 to 10.76 mg/g.
FRAP reduction activity ranged from 3.53 to 13.55 mg/g. L. pauciflorus showed the highest antioxidant activity at 13.55 mg/g, significantly higher than the other samples (p < 0.05). This was followed by P. 'Spring of Beilin' (10.03 mg/g), P. 'Purple Sundae' (9.37 mg/g), and P. 'The Smurfs' (8.99 mg/g). The lowest antioxidant activity was observed in P. medica at 3.53 mg/g.
The results of antioxidant activity determined by the four methods were largely consistent, with L. pauciflorus showing the strongest activity, indicating significant differences among genera. Significant differences were also observed among the different taxa of Primulina, with the antioxidant activity of P. 'Purple Sundae' and P. 'Spring of Beilin' being particularly prominent.
Correlation analysis between bioactive components and antioxidant activity
-
To further investigate the potential relationship between antioxidant activity, TPC, and TFC, a correlation analysis was performed (Fig. 6). Both TFC and TPC showed strong positive correlations (0.72 < R < 0.99, p < 0.05) with scavenging and reducing abilities. Plants with high phenolic and flavonoid contents generally exhibit strong antioxidant effects, and previous studies have shown that polyphenolic compounds in plant extracts contribute to their antioxidant activity and reducing capacity[28,29]. The results of this study are consistent with this view. In addition, the correlation statistics of the antioxidant results showed R > 0.94 and p < 0.05, indicating a highly significant positive correlation between the four selected methods, confirming their suitability for measuring antioxidant activity in Gesneriaceae.
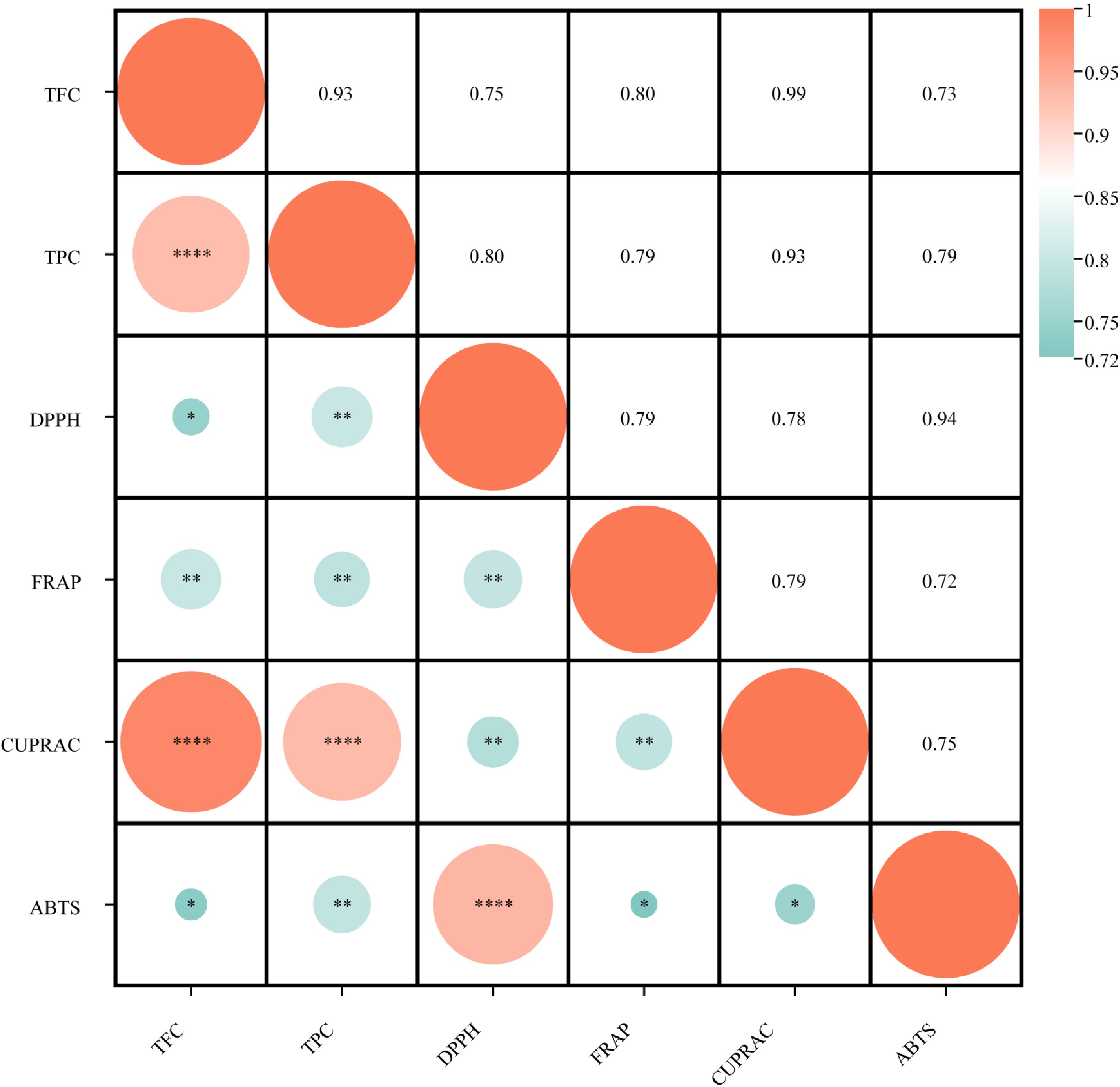
Figure 6.
Correlation between TPC, TFC, and DPPH, ABTS, CUPRAC, and FRAP (* p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001).
Comprehensive evaluation of antioxidant activity
-
To eliminate the bias of single-indicator evaluation, the weighted membership function method was employed to calculate the standard deviation coefficients for each indicator, and weights (D values) were assigned to the membership function values of each indicator, as shown in Table 1. Ten taxa of Gesneriaceae were comprehensively evaluated and ranked based on the comprehensive evaluation value. Within the genus Primulina, P. 'Purple Sundae' and P. 'Spring of Beilin' were identified as the top three excellent resources.
Table 1. Membership function values and composite evaluation values of ten Gesneriaceae taxa.
Name Membership function values D value Rank TFC TPC DPPH FRAP CUPRAC ABTS L. pauciflorus 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1 P. 'Purple Sundae' 0.24 0.40 0.81 0.58 0.19 0.94 0.43 2 P. 'Spring of Beilin' 0.18 0.45 0.76 0.65 0.27 0.62 0.41 3 P. 'The Smurfs' 0.13 0.11 0.58 0.54 0.17 0.37 0.27 4 P. medica 0.14 0.27 0.49 0.00 0.19 0.48 0.22 5 P. 'Nandia Dreams' 0.08 0.21 0.25 0.36 0.09 0.38 0.20 6 P. 'Venus' 0.08 0.00 0.25 0.25 0.11 0.35 0.15 7 P. fimbrisepala 0.00 0.13 0.27 0.20 0.08 0.36 0.13 8 P. 'Four Seasons' 0.09 0.10 0.18 0.31 0.03 0.12 0.12 9 P. eburnea 0.02 0.17 0.00 0.19 0.00 0.00 0.06 10 Weight 0.25 0.15 0.07 0.16 0.24 0.12 Correlation analysis of major metabolites and antioxidant activity
-
The metabolites present in the leaves of Gesneriaceae play a crucial role in contributing to its antioxidant activity. To identify the key antioxidant components in Gesneriaceae leaves, Pearson correlation analysis was performed to elucidate the relationship between metabolites and antioxidant activity (Fig. 7). Among the DAMs identified, phylloflavanine and isoacteoside were positively correlated with the antioxidant activity of Gesneriaceae leaves. Phylloflavanine has been reported in Phyllocladus trichomanoides[30], and isoacteoside can repair free radical-induced oxidative damage and have antioxidant functions[31−33], which is consistent with the results of this study. In addition to antioxidant activity, isoacteoside has many biological effects, such as neuroprotection, antitumor, hepatoprotection, and antiapoptosis[34−36]. The effects of these compounds are consistent with the main functions and indications of Gesneriaceae in the history of traditional Chinese national medicine[37]. In differential metabolite analysis, these two compounds were upregulated in P. 'Purple Sundae', P. medica, P. 'Four Seasons' and L. pauciflorus.
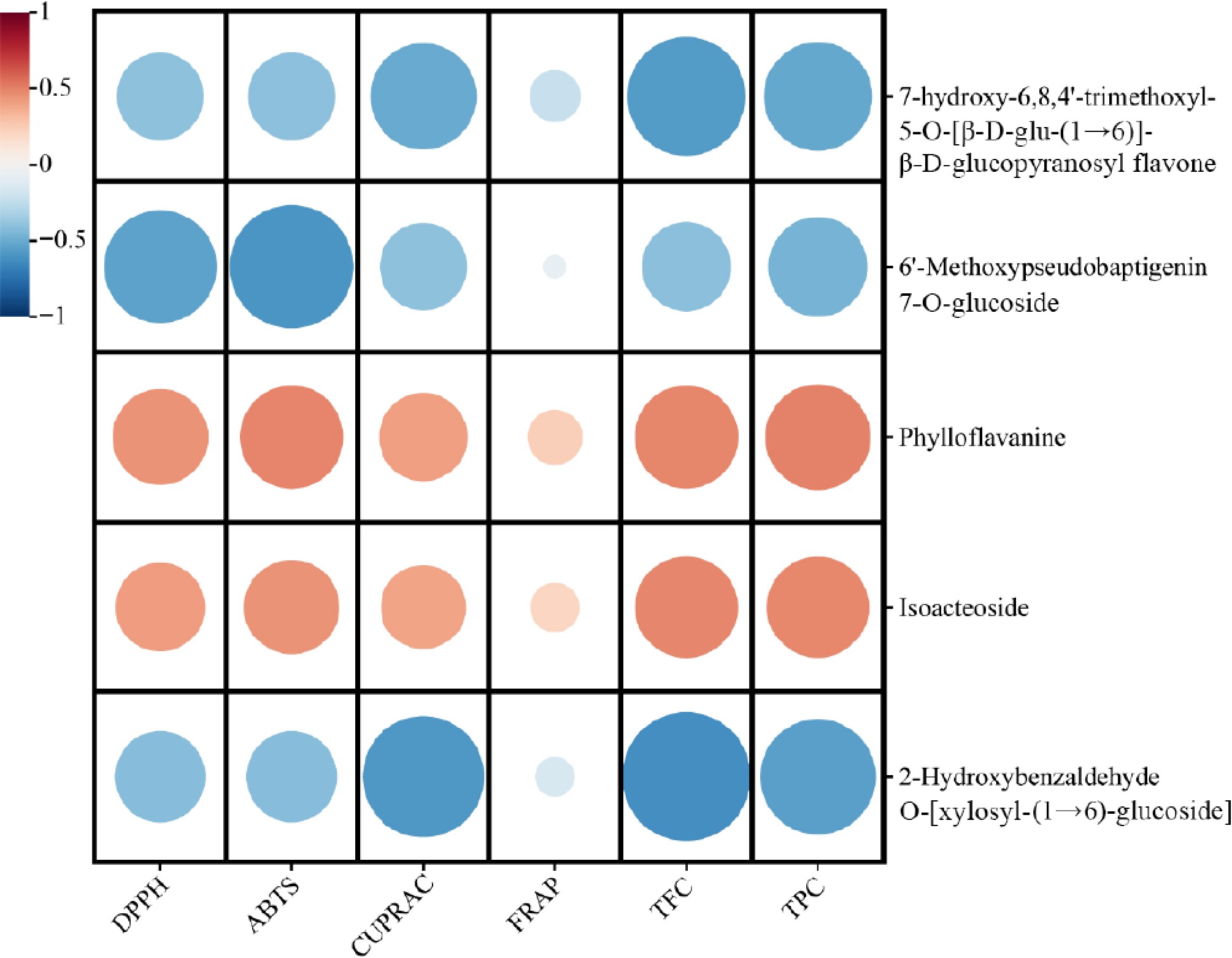
Figure 7.
Correlation analysis of metabolites with total flavonoid content (TFC), total phenolic content (TPC), and antioxidant activity in leaves of Gesneriaceae.
-
Polyphenols are biologically significant in Gesneriaceae plants, with studies revealing their widespread presence and diverse functions in ornamental plants[38−40]. In this study, metabolomics analysis further verified these findings. The metabolite composition of leaves from ten Gesneriaceae taxa was systematically analyzed, tentatively identifying a total of 48 metabolites, including polyphenolic compounds such as flavonoids, phenolic acids, and phenylethanol glycosides, which exhibited significant antioxidant and anti-inflammatory properties. These phenylethanol glycosides are prevalent in several genera within Gesneriaceae, but the distribution of various species of phenylethanol glycosides varies significantly across different plant taxa[41], representing an important chemical characteristic that aligns with the findings of differential metabolites. Hierarchical cluster analysis further revealed significant differences in the metabolite distribution patterns across the leaves of the ten Gesneriaceae taxa, grouping them into two main clusters. However, this study faces certain limitations in metabolite annotation and pathway analysis. Since many of the initially identified metabolites are relatively novel, currently available public databases are insufficient to support comprehensive metabolic pathway analysis. To address this, a correlation analysis between differential metabolites and antioxidant activities was conducted to explore their potential biological significance. Notably, the synergistic expression pattern of phylloflavanine and isoacteoside may reflect an adaptive strategy of the plants to environmental challenges. This observation aligns with previous findings on the combinatorial effects of plant metabolites[42], suggesting that plants may improve their environmental adaptability by modulating synergistic interactions among multiple metabolites. From a metabolic regulation perspective, the significant positive correlation between phylloflavanine and isoacteoside implies a potential biosynthetic link. Plants may strategically produce multiple defense compounds by fine-tuning related metabolic pathways[43]. However, metabolite interactions are not always synergistic. The significant negative correlation observed between 7-hydroxy-6,8,4'-trimethoxyl-5-O-[β-D-glu-(1→6)]-β-D-glucopyranosyl flavone and both phylloflavanine and isoacteoside likely indicates complex interplay among these compounds. It has been demonstrated that metabolite interactions can influence biological activities through various mechanisms[44]. Additionally, these compounds may exhibit either synergistic or antagonistic effects within the cell by activating or inhibiting distinct signaling pathways[45]. Therefore, it is hypothesized that this negative correlation may result from their intricate interactions, although the precise mechanisms warrant further investigation. The multifaceted biological activities of phylloflavanine and isoacteoside, along with their predominant expression in Group 1 plants, may provide a competitive advantage within specific ecological niches. This result further emphasizes the key role of polyphenols in the chemical composition of the leaves of Gesneriaceae. Investigating the potential functions of polyphenols in Gesneriaceae contributes to a better understanding of the plant's biological characteristics and potential applications.
As key products of plant secondary metabolism, polyphenolic compounds not only impart vibrant colors and unique physiological properties to plants[46,47], but are also closely associated with the plant's ability to withstand both abiotic and biotic stresses. In this study, the bioactive substance content and antioxidant activity of Gesneriaceae leaf extracts were thoroughly investigated. The results indicated that all ten tested taxa exhibited significant antioxidant activity, with notable differences among them. These differences were strongly correlated with the total phenolic and flavonoid content, suggesting that polyphenols play a central role in the antioxidant process[48]. Studies have shown that plants like Trollius europaeus and Epipactis atrorubens enhance disease resistance and environmental adaptability by increasing polyphenol content[49,50]. Furthermore, studies on Paeonia delavayi[51] and Astragalus membranaceus[52] indicate that plants with higher polyphenol content tend to exhibit increased antioxidant activity, greater ecological adaptability, and enhanced ornamental value.
Additionally, active compounds in Gesneriaceae plants demonstrate significant medicinal and edible potential. Polyphenols extracted from Haberlea rhodopensis exhibit anti-aging properties[53]. Natural flavonoids extracted from Primulina juliae exhibit analgesic effects[54]. As a new functional vegetable, P. juliae has high biomass and rich calcium content[55]. These findings indicate not only the potential of Gesneriaceae plants in the medicinal field but also provide a robust theoretical basis for their application in the food industry.
By studying the metabolites, bioactive substance content, and antioxidant activity of Gesneriaceae leaves, this study has provided a deeper understanding of the functions of these plants. This research demonstrates that Gesneriaceae leaves are rich in polyphenols and exhibit strong antioxidant activity, which enhances the plant's environmental adaptability and supports its potential for multifunctional development in medicine and food. Future research could further explore the synthesis pathways and regulatory mechanisms of polyphenols to enhance their content and antioxidant activity in plants through genetic engineering. Additionally, optimizing new ornamental varieties of Gesneriaceae with medicinal and edible potential could enhance their ecological benefits, ornamental appeal, and economic value.
-
This study revealed that the leaves of the ten Gesneriaceae taxa tested were rich in polyphenolic compounds and exhibited strong antioxidant activity. Wild species such as L. pauciflorus and P. fimbrisepala are indispensable in traditional ethnomedicine. However, their limited ornamental and commercial appeal restricts their use in modern horticulture and commercial applications. The new flower varieties that were selected and bred possess enhanced ornamental value and multifunctional development potential. Through comprehensive evaluation, two new varieties with notable advantages were identified: P. 'Purple Sundae' and P. 'Spring of Beilin', as well as two additional promising candidates, P. 'The Smurfs' and P. 'Nandia Dreams', which demonstrated comparable performance. These new varieties not only exhibit a pleasing aesthetic but also possess potential medicinal and nutritional values, meeting the modern market's demand for diversified ornamental plants. In the future, optimizing the metabolite compositions of these varieties could enable the development of new cultivars with enhanced environmental adaptability through targeted breeding. This approach would enable Gesneriaceae to thrive in diverse environmental conditions and highlights its potential for development in medicinal and food-related applications. Therefore, as a plant with multifunctional applications, it is worthwhile to pursue in-depth research and cultivation efforts to promote its broader application across various fields.
This research was funded by the Fundamental Research Funds for the Central Universities (Grant No. QNTD202306); The Special Research Funds for the Central Universities (Grant No. 2015ZCQ-YL-03); Horizontal project commissioned by enterprises and institutions for scientific and technological projects (Grant No. 2020HXFWYL20). We are grateful for the instrument support provided by the large-scale instrument-sharing platform of the Institute of Chinese Materia Medica, China Academy of Chinese Medical Sciences.
-
The authors confirm their contributions to the paper as follows: study conception and design: Shen Z, Luo L, Wan H; sample collection: Shen Z, Wu H, Liu Y; laboratory analysis: Shen Z; draft manuscript preparation: Shen Z; feedback on the analysis and manuscript: Luo L, Wan H. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Identification in leaves of 10 Gesneriaceae taxa.
- Supplementary Table S2 Identification's peak area in leaves of 10 Gesneriaceae taxa.
- Supplementary Table S3 Optimized MRM parameters for 48 metabolites in leaves of 10 Gesneriaceae taxa.
- Supplementary Table S4 DAMs in comparison groups.
- Supplementary Fig. S1 Total ion chromatograms (TIC) of QC samples.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shen Z, Wu H, Liu Y, Wan H, Luo L. 2025. Chemical composition and antioxidant potential of ten ornamental Gesneriaceae taxa. Ornamental Plant Research 5: e031 doi: 10.48130/opr-0025-0028 

Chemical composition and antioxidant potential of ten ornamental Gesneriaceae taxa
- Received: 31 December 2024
- Revised: 29 April 2025
- Accepted: 14 May 2025
- Published online: 11 August 2025
Abstract: Gesneriaceae is an important wildflower resource in China, with many new varieties of high ornamental value becoming increasingly popular in the market. Additionally, Gesneriaceae species have been used extensively in traditional ethnomedicine. Nevertheless, comprehensive studies on the chemical constituents and antioxidant activities of Gesneriaceae leaves remain limited. In this study, metabolomic analysis of leaf chemical components from ten Gesneriaceae taxa tentatively identified 48 metabolites across ten compound classes, including flavonoids, phenolic acids, and terpenoids. To further investigate the biological functions of these metabolites, the total flavonoid and phenolic contents were measured, and their in vitro antioxidant activities were evaluated. The results indicated significant variability in antioxidant activities across Gesneriaceae taxa, with phylloflavanine and isoacteoside as key components influencing this activity. Finally, through comprehensive evaluation, two new ornamental varieties—Primulina 'Purple Sundae' and Primulina 'Spring of Beilin'— were identified for their medicinal and edible development potential, providing a theoretical foundation for exploring multifunctional value in Gesneriaceae ornamental plants.
-
Key words:
- Gesneriaceae taxa /
- Metabolomics /
- Polyphenols /
- Antioxidant activity