









-
Rice (Oryza sativa L.) is extensively cultivated across Asia, Africa, Central and South America, serving as a staple crop for half of the global population. Currently, the world is grappling with significant food security challenges, making the enhancement of rice yield a critical objective in rice breeding. The architecture of the rice plant plays a pivotal role in determining yield, thereby making its improvement a key strategy in breeding efforts. Plant architecture is dynamically regulated by developmental and environmental factors, primarily encompassing plant height, tiller number, tiller angle, and panicle morphology. Optimal plant architecture can enhance the light interception, source-sink-flow structure, and ventilation, thereby reducing the incidence of pests and diseases and increasing yield. To achieve higher yield, researchers have proposed an ideal plant architecture model, which includes typical characteristics such as compact plant architecture, a high number of effective tillers, moderate panicle size, moderate plant height, green leaves, yellow seeds, and thick stems[1−3].
Strigolactones (SLs), a new class of plant hormones, have been shown in recent studies to play a crucial role in regulating plant architecture, particularly in shoot or tiller growth, primary growth, and lateral root development[4,5]. SLs are also involved in regulating root morphology, photomorphogenesis, leaf morphology, and senescence, as well as responses to biotic and abiotic stresses[6,7]. SL molecules released from plant roots into the rhizosphere trigger the branching of arbuscular mycorrhizal fungi and their symbiosis with the plant, thereby improving nutrient absorption by aboveground plants[8]. Through the study of SL-insensitive mutants in rice, three genes were identified to be involved in SL perception and signal transduction. D14, a signal-sensing molecule, encodes a protein from the α/β-hydrolase fold family, which binds to the hydrolyzed SL molecule to form a covalently linked intermediate molecule (CLIM)[9]. This reaction triggers a conformational change and forms a complex with D3, which encodes an F-box-like protein. The d3 mutant in rice exhibits a dwarf and high-tillering phenotype[10], and the homologous max2 mutant in Arabidopsis also exhibits multiple branches[11]. In the presence of SL molecules, D14 interacts with D3 and D53, resulting in D53 ubiquitination and the degradation of the transcriptional repressor D53 through the 26S proteasome, thereby activating the expression of downstream genes and inhibiting axillary bud growth[12,13]. The D53 protein contains three EAR motifs, which are essential for recruiting the transcriptional repressor TPL protein and inducing its oligomerization to form a receptor-coreceptor-nucleosome complex[14]. D53 can interact with the TPL protein TPR2 in rice, suggesting that the complex synergistically inhibits the expression of SL downstream target genes[12]. Studies have shown that the homologous genes of D14, D3, and D53 have similar functions in angiosperms Arabidopsis and peas[15,16]. A partial loss-of-function allele of the SL biosynthesis gene HTD1/D17 (HIGH TILLERING AND DWARF 1/DWARF17), which encodes CAROTENOID CLEAVAGE DIOXYGENASE 7 (CCD7), resulted in increased tiller number and improved grain yield in the mutants. Moreover, the HTD1 gene has been widely used and co-selected with SEMI-DWARF 1 (SD1)[4].
Although many functional genes regulating plant architecture have been identified, few of them have been applied to breeding practices. Gene-editing techniques have been developed from ZFN and TALEN to CRISPR/Cas9. CRISPR/Cas9 is a rapid, efficient, inexpensive, and easy technique for achieving knockout gene in the cell[17−20]. With the advancement of molecular breeding technology, CRISPR/Cas9-mediated gene editing has been employed in crop engineering. For instance, OsPAGN1 and OsPDCD5, which negatively regulate plant height, were knocked out using CRISPR/Cas9, resulting in increased plant height, tiller number, and grain number per panicle[21,22]. Editing a key negative regulator TAD1, a subunit of E3 ligase, led to mutant plants with increased tiller number and reduced plant height, thereby improving yield[23]. Mutants of another gene, OsFWL4 (fwl4), exhibited increased tiller number but reduced 1,000-grain weight and grain thickness[24,25].
A rice line, DS (japonica rice), exhibiting cold tolerance and superior grain quality but with excessive plant height and limited tillering, was previously selected. Since mutant plants of the key gene D3 in the SL signaling pathway showed increased tillering and dwarfism[10], the D3 gene in DS was targeted for editing to improve plant architecture. To this end, CRISPR/Cas9 technology was applied to edit the D3 gene in DS, resulting in an improved rice line without transgenic components. In addition, the role of D3 in resistance to rice bacterial blight was investigated, revealing that the d3 mutant showed enhanced resistance.
-
DS (japonica rice), originating from Yibin Academy of Agricultural Sciences, China, is renowned for its cold tolerance and superior grain quality. This variety was used as the genetic background for gene editing to improve plant architecture. DS and d3 rice mutants were grown under natural conditions in an experimental field at Sichuan Agricultural University, China.
Construct, transformation, and genotyping of gene-edited lines
-
The target gRNA sequence in the exon region of the D3 gene was designed using the website (
http://skl.scau.edu.cn/targetdesign/ )[26] (Fig. 1a) and constructing Cas9-D3 plasmid with the CRISPR-Cas9 vector pRGEB32 following a previous study[27]. Cas9-D3 was genetically transformed into DS mediated by Agrobacterium strain EHA105. The primer sequences are listed in Supplementary Table S1. The resultant transgenic plants were examined for mutations in D3 by polymerase chain reaction and DNA sequencing.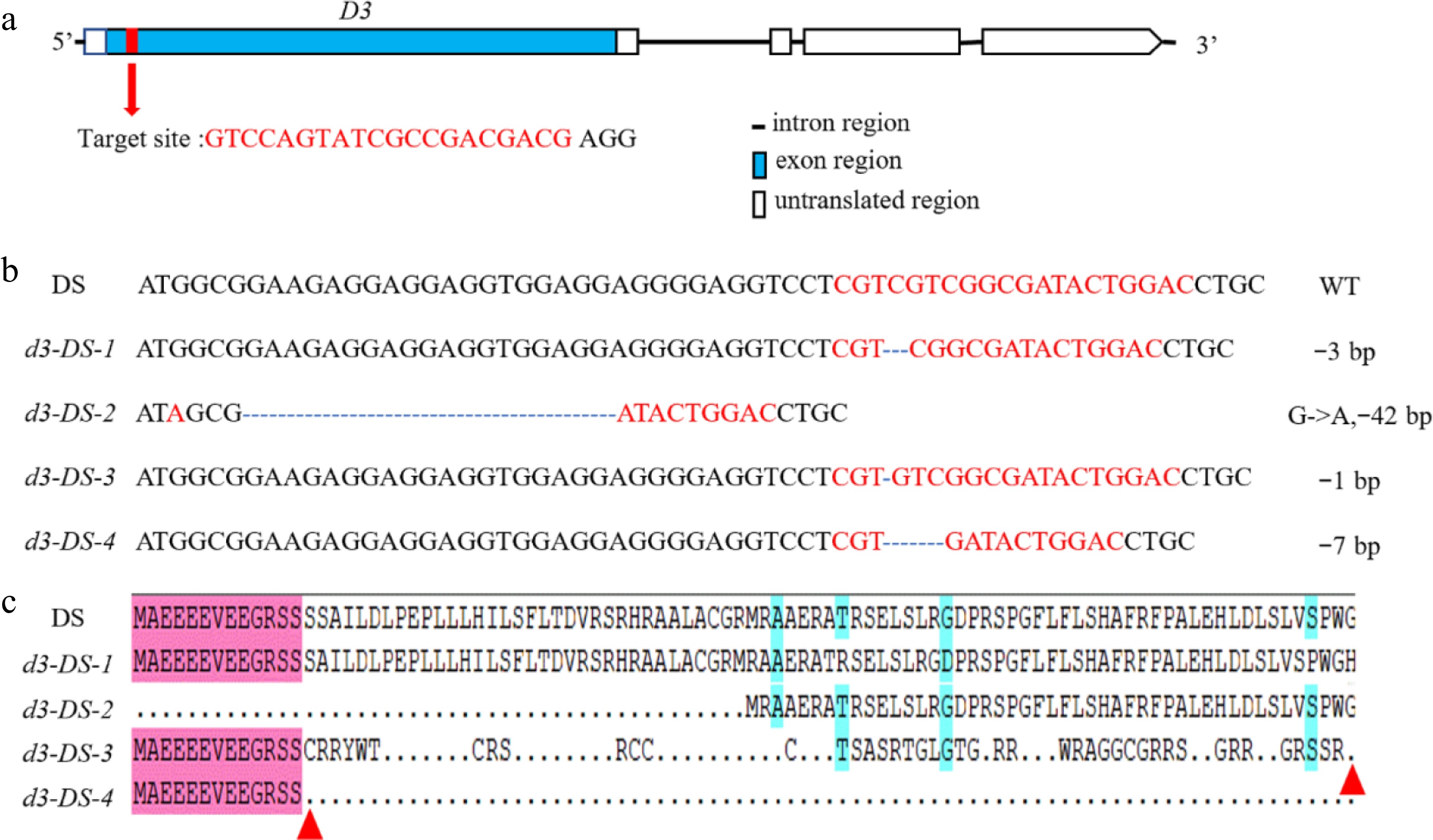
Figure 1.
Loss-of-function mutations of D3 by CRISPR/Cas9. (a) The gene structure of D3 and the location of the target site. Introns are represented by straight lines, exons are represented by blue boxes, untranslated regions are shown as white boxes, and the target site is marked with a red box. (b) Sequence alignment illustrating mutations of four types of d3 mutants. '-' indicates deletion, and numbers indicate nucleotides. (c) Amino acid sequence alignment using DNAMAN. The red triangle indicates the termination site.
To assess the genotypes of the CRISPR/Cas9 mutants, genomic DNA was extracted from rice leaves by the CTAB method[28]. Subsequently, a PCR-based approach was employed to amplify the target fragments using designed target-specific primers (Supplementary Table S1), followed by DNA sequencing to confirm the presence of mutations at the target sites.
Hygromycin resistance identification
-
The leaves of rice plants were first marked with a marker pen, excised, and then immersed in distilled water containing 30 μg/mL hygromycin (HYG) and 1 μg/mL 6-benzylaminopurine (6-BA), with positive and negative controls established. After 2–3 d, the leaves of HYG negative plants exhibited cell death, characterized by the appearance of yellow, ring-like spots. In contrast, the leaves of HYG-resistant plants showed no such symptoms.
Investigation of agronomic traits
-
Each rice line was planted in three rows, with ten plants per row. Plant height, tiller number, and seed-setting rate were measured at rice maturing stages. Grain length, grain width, 1,000-grain weight, and grain number were quantified using the Wanshen SC-G seed tester. The experimental design followed standard practices in transgenic plant phenotyping, in which 30 plants were initially grown per line, with edge-row plants excluded to minimize environmental variability (e.g., light, nutrient gradients). From the remaining plants, five phenotypically consistent individuals were selected for detailed analysis. This approach aimed to reduce biological noise caused by developmental or growth-stage differences, thereby improving the reliability of trait comparisons, even with smaller sample sizes (as supported by the research of Hu et al. [29]).
Xoo inoculation
-
Xoo strains P6 and P3 were cultured in potato sucrose agar medium at 28 °C.
The leaf wound infection method was employed for bacterial blight pathogen inoculation[30]. The inoculation was conducted in the field, where temperatures fluctuated diurnally. It was performed at the late tillering stage of rice (40 d after transplanting), under weather conditions conducive to minimizing the impact of rainfall on the results. Considering the rice growth stage, disease development occurred at temperatures between 28–34 °C and relative humidity of ≥ 80%. Fully expanded rice leaves were excised 1 cm from the top after immersion of scissors in the bacterial blight culture solution. Four plants were selected for each strain, with eight leaves inoculated per strain. The suspension concentration of the P6 and P3 strains was OD600 = 0.6. Lesion length was measured at 2–3 weeks after inoculation.
-
To make a D3-targeted construct, the D3 gene sequence in DS was identified and confirmed to be identical to that of Nipponbare (NPB; Supplementary Fig. S1). A gene-specific guide RNA was then designed to edit D3 in DS using the CRISPR/Cas9 technology (Fig. 1a). Multiple transgenic T0 lines with four types of mutations were obtained. Among them, the d3-DS-1 line had a 3-bp deletion, resulting in the loss of one amino acid residue in the D3 protein. The d3-DS-2 mutant harboured a G-to-A substitution at the initiation codon and a 42-bp deletion afterwards, leading to truncation of 47 amino acids at the N-terminus of D3 protein. The d3-DS-3 and d3-DS-4 mutants contained 1-bp and 7-bp deletions, respectively, causing frameshifts and premature stop codons (Fig. 1b, c).
To obtain homozygous d3 mutants devoid of transgenic elements, the T1 generation was screened for hygromycin-negative plants. Specific primers on the Cas9 vector were designed to identify T1 generation plants by PCR. The results revealed 20 homozygous mutants among the T1 generation, including two plants devoid of transgenic elements, both identified as d3-DS-2 mutants. Subsequently, the T2 generation was analyzed. Thirty d3-DS-2 plants were identified, all lacking transgenic elements. Additionally, 10, 30, and 30 plants of the d3-DS-1, d3-DS-3, d3-DS-4 homozygous mutants were respectively detected, yielding four, eight, and nine homozygous mutants without transgenic elements for d3-DS-1, d3-DS-3, and d3-DS-4, respectively (Table 1).
Table 1. Identification of 'transgene-free' homozygous d3 mutant plants in T1 and T2 generations.
T1 homozygous mutant line T1 with transgenic elements Number of examined T2 plants Number of T2 'transgene-free' plants d3-DS-1 Positive 10 4 d3-DS-2 Negative 30 30 d3-DS-3 Positive 30 8 d3-DS-4 Positive 30 9 Reduction of plant height and increase of tiller numbers in the d3 mutants
-
To confirm the stable inheritance of these traits in the offspring, agronomic traits of the T2 generation were further analyzed, and five plants from each mutant type were selected for assessment. All T2 plants were derived from the T1 generation, in which four types of d3 homozygous mutants had been identified (Supplementary Fig. S2). The results showed that the agronomic traits of the four homozygous mutants were indeed stably inherited by the subsequent generation (Fig. 2a). Analysis of plant height revealed that the d3-DS-2, d3-DS-3, and d3-DS-4 mutant lines were significantly shorter than the wild-type DS, exhibiting a height difference of 19.4 ± 3 cm, while d3-DS-1 showed a slight reduction of 5.5 ± 5 cm compared to DS (Fig. 2b). Measurements of the main stem indicated that the internodes of d3-DS-2, d3-DS-3, and d3-DS-4 mutants were shorter than those of the wild type (Fig. 2b).
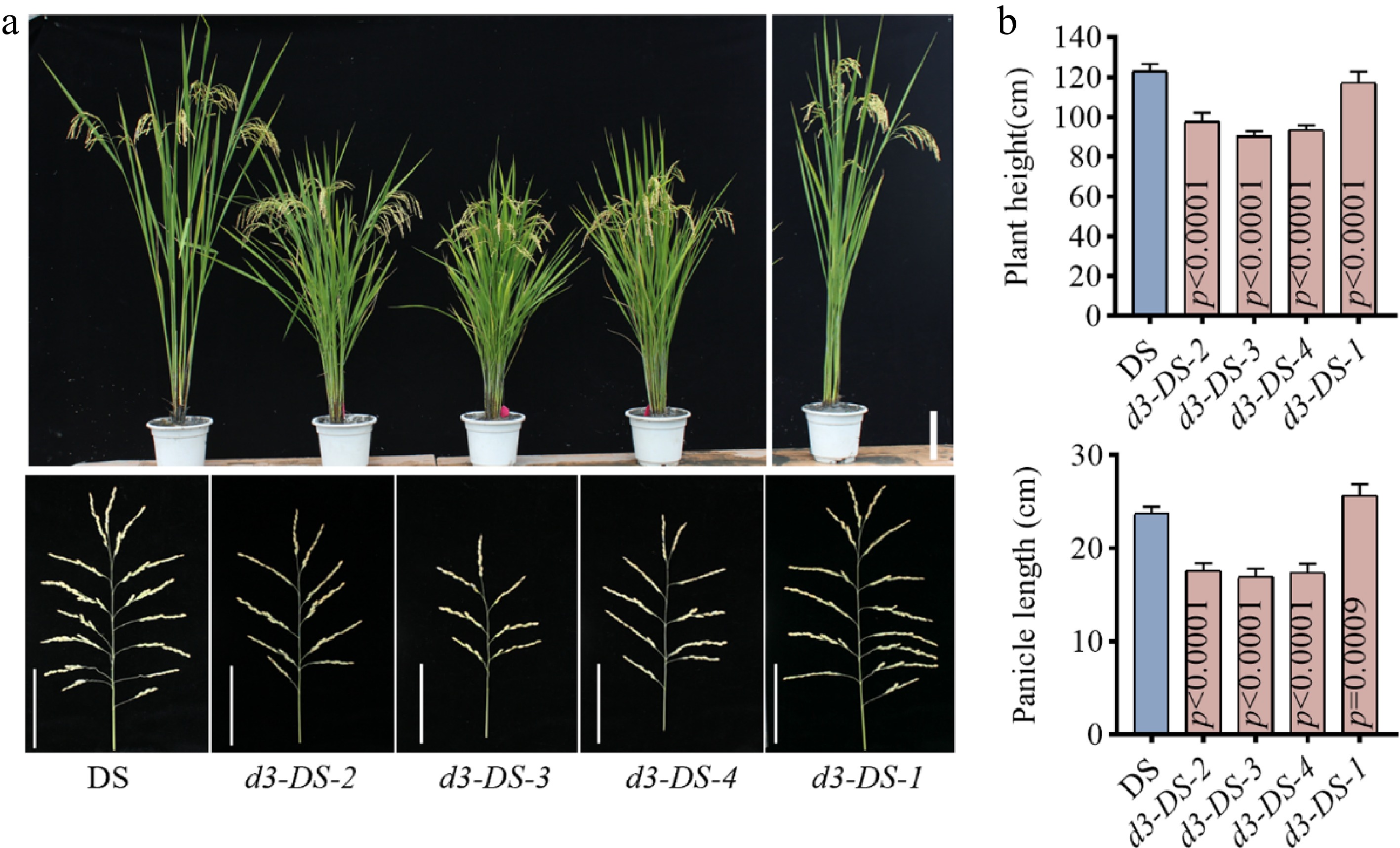
Figure 2.
Phenotypes of d3 mutants. (a) The upper and lower parts of the figure are the plant morphology and panicle morphology of the T2 generation d3 mutants and the wild type DS, respectively. The upper scale bar represents 15 cm, while the lower scale denotes 9 cm. (b) Plant height and panicle length. All P values were determined by unpaired Student's t-test, compared to DS.
Tiller number analysis showed that the d3 mutants of d3-DS-2, d3-DS-3, and d3-DS-4 had an average of 20 panicles per plant, significantly exceeding the wild-type DS and the d3-DS-1 lines, which averaged 7.9 and 7.5 panicles, respectively (Fig. 3a). Although increasing tiller number is generally beneficial for yield, d3-DS-2, d3-DS-3, and d3-DS-4 mutants displayed shorter panicle length (Fig. 2b), accompanied by reductions in both grains per panicle and filled grains per panicle (Fig. 3b, c). By contrast, the seed setting rate was not significantly changed (Fig. 3d). Measurements of grain width, grain length, 1,000-grain weight, and yield per plant showed no significant differences between the four mutant lines and the wild type DS (Fig. 3e–h). Together, these results indicate that the d3 mutants d3-DS-2, d3-DS-3, and d3-DS-4 exhibit improved plant architecture compared to DS, characterized by shorter plant height and significantly increased tillers. Conversely, the d3-DS-1 line with a 3-bp deletion did not show significant differences in agronomic traits compared to the wild type (Fig. 3a–h).
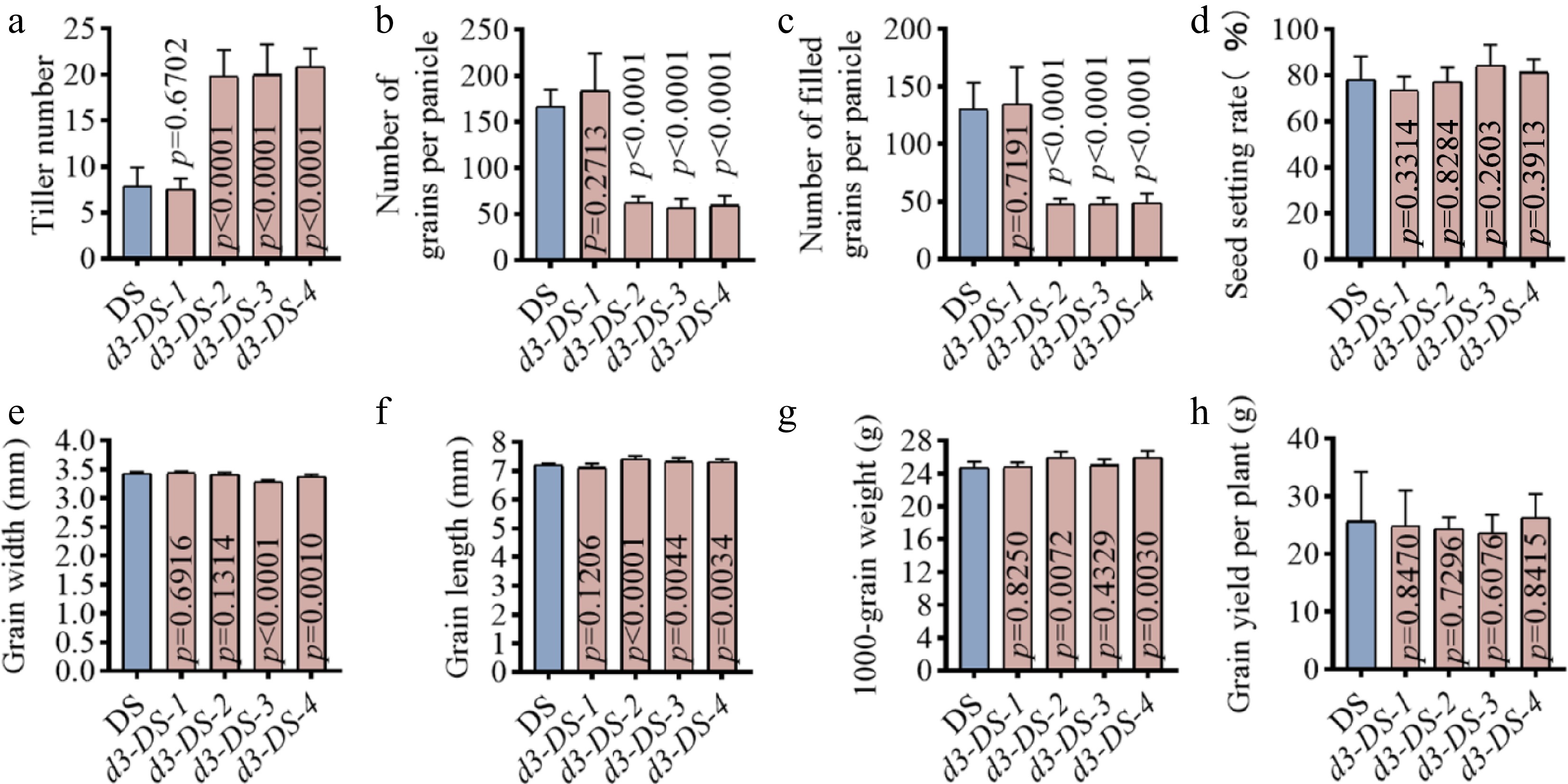
Figure 3.
Yield traits of d3 mutants. Major yield traits were measured at rice maturing stages. (a) Tiller number. (b) Number of grains per panicle. (c) Number of filled grains per panicle. (d) Seed setting rate. (e) Grain width. (f) Grain length. (g) 1,000-grain weight. (h) Grain yield per plant. All p values were determined by unpaired Student's t-test, compared to DS.
Early heading time in d3 mutants
-
To investigate the impact of D3 gene knockout on the growth period, a comparative analysis of leaf age was conducted between the d3 mutants and the wild-type DS. Eleven plants were selected from each of the mutant lines (d3-DS-3 and d3-DS-4) and the wild type. Leaf age was recorded starting from the 5-leaf stage after transplanting and then every 6 d until reaching the full heading stage. The results showed a significant difference in leaf age between the d3 mutants and DS, with an average deviation of 1.2 leaves. The inverted triangle marker in Fig. 4a illustrates that the leaf age of the wild type and the d3 mutants began to diverge from June 12. As a result, the heading time of the d3 mutants occurred earlier than that of the wild-type DS (Fig. 4b, c).
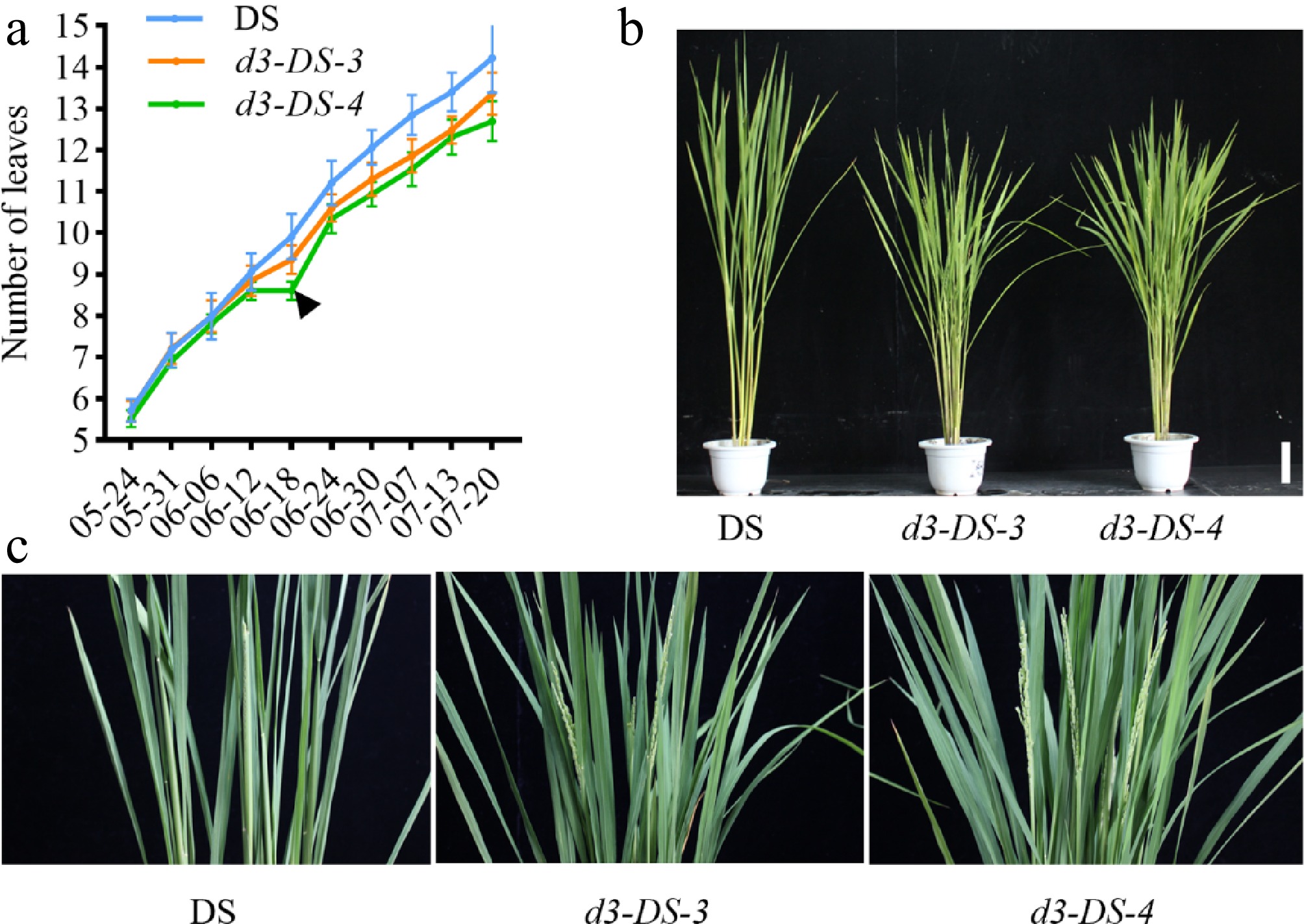
Figure 4.
Growth period analysis of d3 mutants. (a) Monitoring of leaf age of d3 mutants and the wild-type DS. The black inverted triangle marks the point when the leaf age difference emerged. (b) The heading stage phenotype of the d3 mutants and wild type in the field. (c) A close-up of the panicle part, with a scale bar of 10 cm.
Enhanced resistance against Xoo in d3 mutants
-
To determine the changes in disease resistance for the D3 gene knockout, Xanthomonas oryzae pv. oryzae (Xoo), the causative pathogen of a major rice disease, was inoculated onto the d3 mutants and the wild-type DS. During the rice tillering stage, two Xoo strains, P6 and P3, were selected for field inoculation. Eighteen days after inoculation, statistical analysis revealed that the lesion length in the mutants inoculated with the P6 and P3 strains was reduced by 22%–38% compared to the wild type (Fig. 5a, b). These data indicate that the d3 mutants exhibit enhanced resistance to bacterial blight compared to the wild-type DS.
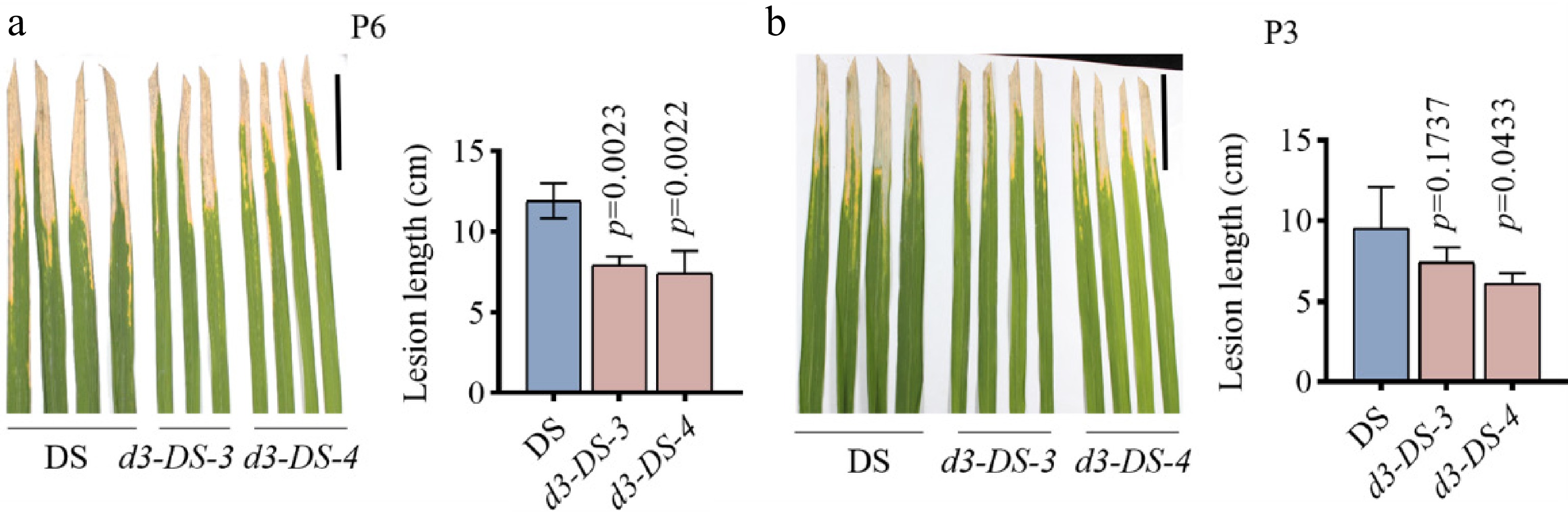
Figure 5.
Inoculation of Xoo on d3 mutants. (a), (b) Phenotype and lesion length statistics of the d3 mutant and wild-type DS inoculated with P6 and P3 strains after 18 d. The scale bar represents 5 cm, with P6 and P3 strain suspension concentration set at an OD600 of 0.6. All p values were determined by unpaired Student's t-test, compared to DS.
-
Genome-editing technology has been successfully used to achieve multiple desired agronomic traits in rice. For instance, employing CRISPR/Cas9 technology conducted multi-gene editing targeting TMS5, Pi21, and Xa13 in rice, yielding homozygous triple mutant lines without transgenic elements in the T1 generation[31]. These mutants exhibited thermo-photosensitive genic male sterility along with enhanced resistance to rice blast and bacterial blight. Editing the Ehd1 (Early heading date 1) promoter with multiple target sites reduced Ehd1 expression, delayed heading date, and improved yield-related traits in ZJ8[32]. Similarly, CRISPR/Cas9 lines targeting OsHXK1 led to the development of high-photosynthetic-efficiency and high-yielding rice varieties[33]. Additionally, researchers identified RBL1 (RESISTANCE TO BLAST1) in a lesion mimic mutant and obtained an allele, RBL1Δ12, through targeted genome editing. This allele conferred broad-spectrum disease resistance without compromising yield in rice[34].
In the present study, the D3 gene in the rice line DS was edited using CRISPR/Cas9 technology. The resulting d3 mutants showed significantly reduced plant height and earlier heading time, along with increased tiller numbers compared to wild-type DS (Figs 2 and 3a). Importantly, grain yield per plant of the d3 mutants was not significantly changed compared to DS (Fig. 3). Together with enhanced resistance to bacterial blight, the D3-edited transgene-free plants are promising candidates for rice breeding.
Disruption of D3 has previously been reported to result in tillering dwarf phenotypes in rice[10]. In this study, it was further demonstrated that panicles of d3 mutants became smaller and grain number per panicle also decreased (Figs 2 and 3). However, panicle number per plant increased in the d3 mutants, while maintaining normal grain size and grain weight, and thus resulting in no significant changes in yield per plant. These findings broaden the understanding of the roles of D3 in regulating crop yield traits. Earlier studies reported that d3 mutants in the Nipponbare[12] and Kitaake[35] (japonica subspecies) backgrounds exhibited extremely high tiller numbers (over 100), along with severely reduced seed-setting rates and grain yield. Notably, Nipponbare and Kitaake (japonica subspecies) inherently exhibit higher tillering capacity than DS (indica rice) even in the wild-type background[12,35]. Therefore, when the D3 mutation is introduced, their tillering number increases more dramatically, and seed-setting rates decline severely. These differences likely arise from distinct genetic backgrounds and variations in growing conditions (e.g., climate, field vs controlled environments).
In addition, D3 amino acid sequence indicated the presence of an N-terminal F-box-like and a leucine-rich repeat (LRR) in the middle, suggesting that D3 is a member of the F-box LRR family of proteins that function as a component of the ubiquitin E3 ligase complex[10]. Evidence indicates that the rice D3 protein interacts with the ASK1 protein through its F-box-like domain. Additionally, by examining the three-dimensional structures of AtASK1 and D3 (NCBI: 5HYW), it was observed that, apart from the F-box-like domain, the interaction with ASK1 may also involve the β1 and β2 sheets of the D3 protein, as well as the sequence between them[36]. The N-terminal F-box-like domain (residues 14–60 in D3) is essential for interacting with the ubiquitin ligase complex SCF (Skp1-Cullin-F-box), which mediates substrate recognition and degradation of key regulators in plant hormone signaling. In this study, the phenotypic similarity between the d3-DS-2 mutant (harboring an N-terminal deletion) and the frame-shifted d3-DS-3/d3-DS-4 mutants suggests that the deletion of the 47 amino acids − including the entire F-box-like motif − likely disrupts D3's ability to recruit substrates for ubiquitination, thus abolishing its regulatory function in SL signaling.
It has been reported that SL is involved in abiotic stresses such as drought and salt stress in Arabidopsis thaliana[37]. In terms of responses to biotic stresses, SL-deficient and -signaling mutants such as d17 and d14 exhibited enhanced susceptibility to the blast fungus M. oryzae in rice[38]. This might be due to suppression of hydrogen peroxide- and ethylene-related genes. A gain-of-function allele of D53, which encodes a transcription repressor in the SL signaling, also showed increased susceptibility to M. oryzae infection[39]. Moreover, D53 can bind to the promoters of OsPALs, repressing their transcription and defense against M. oryzae. Additionally, SLs are involved in cotton resistance to the fungal pathogen Verticillium dahlia[40]. SL-biosynthesis genes CCD7 and CCD8 likely play a positive role in resistance to V. dahliae through crosstalk with JA and ABA pathways, as well as promoting ROS accumulation. As to bacterial pathogens, disruption of the D3 ortholog (MAX2) in Arabidopsis caused increased susceptibility to the necrotroph Pectobacterium carotovorum and the hemi-biotroph Pseudomonas syringae[41]. However, in this present study, d3 rice mutants displayed increased resistance to the hemibiotroph Xoo (Fig. 5). This difference might be due to different inoculation methods in these two studies. The previous study employed spray inoculation, and thus stomatal conductance regulated by MAX2 can influence the pathogen entering[41]. In contrast, the current study used a wound infection approach for Xoo. Altogether, the SL pathway plays a crucial role in plant resistance to multiple pathogens.
-
This study demonstrates a strategy for developing rice cultivars with improved plant architecture and enhanced disease resistance through CRISPR/Cas9-mediated editing of the D3 gene. Targeted modifications in D3 effectively reduced plant height by 21%–27% compared to the donor parent (DS), while simultaneously increasing tiller number, accelerating heading time, and maintaining grain yield at DS levels. Notably, the edited lines exhibited improved resistance to bacterial blight without compromising agronomic performance. These findings establish D3 as a dual-function target for optimizing rice architecture and disease resilience, offering a promising pathway for sustainable crop improvement.
We thank Professor Xue-Wei Chen from Sichuan Agricultural University (China) for providing the bacterial-blight strains P6 and P3. This work was supported by a grant from the National Natural Science Foundation of China to JF (Grant No. 32372490).
-
The authors confirm their contributions to the paper as follows: study conception and design: He J, He Z, Liu J, Hu X, Li G, Wang H, Huang F, Fan J, Wang W; data collection: He J, He Z, Zhao Y, Huang Y, Yi R, Chen P; software, visualization, and writing − original draft: He J, He Z; methodology, writing − review and editing: He J, He Z, Fan J, Wang W; conceptualization, resources, and funding acquisition: Fan J, Wang W. All authors reviewed the results and approved the final version of the manuscript.
-
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary materials.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Jia-Xue He, Zi-Wen He
- Supplementary Table S1 Primers for experiments.
- Supplementary Fig. S1 Sequence alignment of D3 coding region from DS with that from Nipponbare. The D3 coding sequence compared with D3 coding sequence from Nipponbare by NTI.
- Supplementary Fig. S2 Plant and panicle morphology of four homozygous d3 mutants at T1 generation. The upper and lower parts of the figure are the plant morphology and panicle type of the T1 generation of d3 mutant and DS wild type plant morphology and panicle type, respectively. The upper scale bar represents 15 cm, while the lower scale denotes 9 cm.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
He JX, He ZW, Zhao YD, Huang YQ, Yi R, et al. 2025. Genome editing of the D3 gene improves plant architecture and disease resistance in rice. Plant Hormones 1: e017 doi: 10.48130/ph-0025-0017 

Genome editing of the D3 gene improves plant architecture and disease resistance in rice
- Received: 29 April 2025
- Revised: 29 June 2025
- Accepted: 03 August 2025
- Published online: 25 August 2025
Abstract: CRISPR/Cas9-mediated genome editing has become increasingly prominent in crop breeding, facilitating the simultaneous improvement of multiple agronomic traits. Desired characteristics in rice breeding include plant architecture, yield, quality, and stress resistance. In this study, the engineering of a japonica rice cultivar, DS, renowned for its superior grain quality and cold tolerance, was undertaken. Since the D3 gene acts as a positive regulator of plant height but a negative regulator of tillering, CRISPR/Cas9-mediated gene editing was employed to modify D3 of order to reduce plant height in the DS cultivar. Multiple D3-edited lines were generated, with plant height reduced to 73%–79% of DS. Additionally, these edited lines showed increased tiller numbers, early heading time, and grain yields comparable to those of DS. Moreover, the D3-edited lines exhibited enhanced resistance to rice bacterial blight disease. These findings underscore the utility of gene-editing technology for rapidly enhancing key agronomic traits in rice, while uncovering novel functionalities of the D3 gene in coordinating agronomic traits and disease resistance.
-
Key words:
- CRISPR/Cas9 /
- DS /
- D3 gene /
- Plant architecture /
- Plant height /
- Tiller number /
- Xanthomonas oryzae pv. oryzae