








-
Salinity has a substantial effect on various biological processes (e.g., growth and development) of the plants. Ionic toxicity, osmotic stress, and oxidative damage are only a few detrimental effects on plants caused by the saline stress[1]. Economic growth is determined by plants or crops in most countries; however, salinity has become a challenge for these crops. Various plant species have been studied to enhance their SST traits, focusing on improving their ability to withstand periods of saline stress[2]. However, under saline stress conditions, the data on tolerance mechanisms (especially biochemical, along with transcriptional regulation) of the many plant species are still scarce, and studies are needed to determine mechanisms related to SST[1].
Osmolytes, such as sugars, betaines, and amino acids, are essential to osmotic control because they function as compatible solutes to lessen the detrimental effects of osmotic stress[2]. These osmolytes protect cellular structures and functions from the detrimental effects of saline stress by acting as osmoprotectants in addition to aiding in the regulation of water intake and retention[3]. Additionally, key enzymes like CAT, SOD, as well as ascorbate peroxidase (APX) play a vital role in the plant's defense mechanism. This defense system is bolstered to effectively counteract ROS that are produced when the plant experiences salt-induced stress. Therefore, these adaptations are key for the plants to keep cellular components, including nucleic acid, proteins, and lipids, safe from the damage caused by ROS[4]. Furthermore, in situations of salt stress, the complex web of enzymatic and non-enzymatic antioxidant defenses is essential for reducing the oxidative damage brought on by the buildup of ROS. Plant responses to environmental stresses are complex, and one key mechanism that helps plants survive and thrive in harsh environments is the regulation of gene expression[5]. TFs hold key significance in this regulatory network, coordinating the expression of genes that respond to stress and regulating various plant processes. TF families, including AP2/EREBP, GRAS, MYB, WRKY, bZIP, GATA, bHLH, and NAC, have been discovered as important regulators of SST pathways in a variety of plant species. These TFs regulate a wide range of molecular functions, such as antioxidant defense and osmotic control, which help plants withstand abiotic challenges, mainly salinity stress[6].
There is a lack of comprehensive studies related to SST mechanisms in most plant species, despite their great importance in terms of species development and survival. Therefore, this review aims to uncover the SST mechanisms, viz., biochemical mechanisms linked with transcription regulation developed in plants to enhance tolerance against saline stress. The documentation of existing data on this particular topic through this review will offer a holistic perspective on recent advances in the better understanding of regulatory networks that help plant species to withstand saline stress conditions. Overall, this study will provide a potential pathway for researchers to develop salt-resistant varieties of a number of plants.
-
It is one of the most critical biochemical mechanisms by which plants accumulate compatible solutes or toxic ions that help increase osmotic tolerance. The osmolytes affect how water moves across cellular boundaries, helping the process of osmotic regulation in stomatal opening[7]. The process of water movement within plants is closely associated with a number of purposes, like cellular hydration. In plants, this water movement serves a number of functions related to cell hydration, including increased absorption of water through the root system and stomatal opening. Most of the compounds that take part in osmotic regulation are well known, e.g., glycerol, sorbitol, pinitol, sucrose, mannitol, and arabinitol, organic compounds like oxalate and malate, and nitrogen-containing compounds like glycine, choline, betaine, putrescine, Pro, proteins, and 4-gemma aminobutiric acid[8]. Figure 1 illustrates how various biochemicals and associated molecules mitigate salt stress.
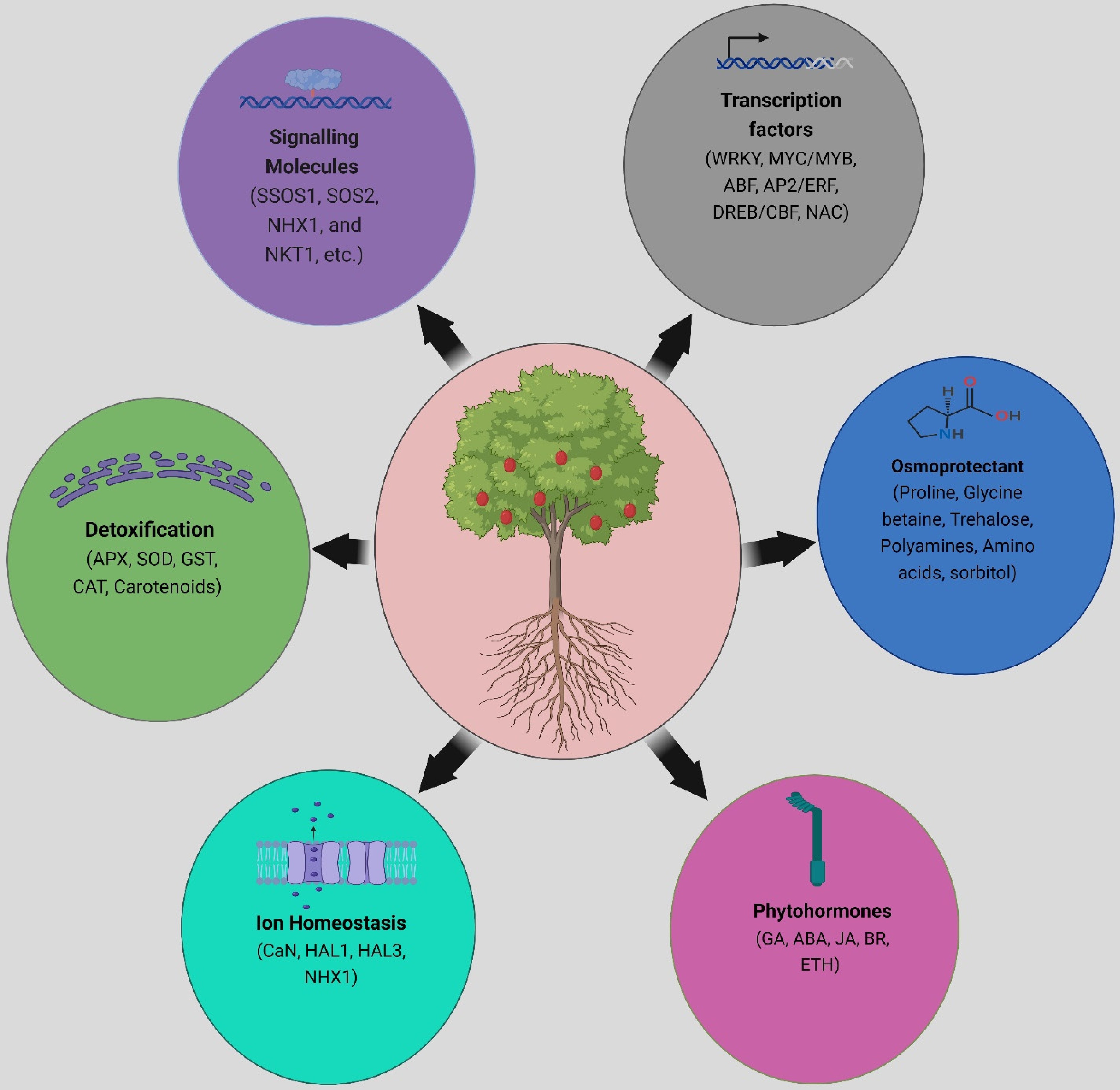
Figure 1.
Various biochemicals and their associated molecules help relieve salt stress. Plants respond to salt stress via a complex network of metabolic pathways regulated by a wide range of biomolecules. Signaling molecules such as SOS1, SOS2, NHX1, and NKT1 play critical roles in ion transport and osmotic control. Enzymes such as APX, SOD, GST, and CAT scavenge ROS, reducing oxidative damage caused by salt stress. Furthermore, the accumulation of carotenoids and osmoprotectants, including Pro, GB, GABA, and sugars, contributes to maintaining cellular homeostasis and osmotic equilibrium in saline environments. These biomolecules help plants withstand salt stress. Furthermore, the activation of TFs such as MYB, WRKY, NAC, ABF, ERF, and DREB is intimately related to the salt stress response, influencing the expression of stress-responsive genes and regulatory pathways. These TFs influence multiple cellular processes involved in salt stress adaptation, demonstrating the interconnectivity of molecular pathways that regulate plant resistance under harsh environmental situations. Created with BioRender.com.
https://BioRender.com/ujpt3z5. Navarro et al. reported that Mediterranean ornamental plants show limited osmotic adjustment when grown in soil with elevated saline levels[9]. According to Cirillo et al., Callistemon citrinus compartmentalizes chloride ions (Cl−) within the leaf vacuole and behaves as a typical Cl− accumulator. The utilization of chloride ions (Cl−) as an osmolyte by this particular plant species serves the purpose of reducing the osmotic potential required for the sustenance of leaf turgor. Even though the osmotic adjustment was moderate (0.2 MPa), however, this favored C. citrinus in maintaining turgor potential under saline conditions[10]. In terms of important organic osmolytes and effective compatible solutes, GB and Pro hold significant importance. Antioxidant properties are reported for Pro, whereas during dehydration, Pro also operates as a molecular chaperone to protect biological macromolecules. Therefore, this helps the plants cope with environmental stresses or maintain growth under stressful conditions. Another vital compound that synthesizes and accumulates under different stress conditions is GB, which protects PSII by acting as an osmolyte. Furthermore, it also protects the activity of enzymes in addition to the integrity of membranes against osmotic stress[11].
The effective compartmentalization of undesirable ions within the vacuoles of plants serves as a pivotal mechanism to mitigate their direct harmful effects. By confining these ions within the vacuolar compartments, plants can minimize the detrimental impact of this stress. Consequently, it is primarily the ions that remain outside the vacuoles that contribute to saline damage, emphasizing the significance of vacuolar compartmentalization in protecting plant health[12]. While vacuolar compartmentalization is considered an effective mechanism for sequestering toxic ions like Na+ and Cl−, conflicting studies highlight its energy-intensive nature[12]. The root is the major organ for the exclusion of salts, which prevent the movement of ions to the plant's aerial parts. Most of the halophytes and some glycophytes utilize this mechanism to survive in case of saline stress[13]. The presence of saline vesicle glands in plants' epidermis can facilitate salt excretion, which is an adaptive strategy to prevent the accumulation of salts in various organs of the plant, like leaves[14]. Old leaves accumulate salts, which cause their death, and the falling of leaves from plants is also another exclusion mechanism of salinized stress. However, the energy and resources required for leaf production, maintenance, and eventual detachment may place a significant metabolic burden on the plant. This trade-off may hinder the plant's ability to sustain growth, especially under prolonged or extreme saline conditions[15]. The role of vacuolar compartmentalization in managing toxic ions such as Na+ and Cl− has been well documented, but there remains a lack of consensus on the energy costs associated with this mechanism. While vacuolar sequestration helps in protecting cellular integrity, the energy-intensive nature of this process, especially under prolonged saline stress, has been noted as a potential metabolic burden[16]. However, some studies have downplayed these costs, focusing instead on the efficiency of the ion transport systems like Na+/H+ antiporters and NHX exchangers. The conflicting views on the metabolic cost of vacuolar compartmentalization highlight a critical gap in understanding the trade-offs between energy allocation for ion sequestration vs other stress mitigation strategies[17]. Further research is needed to model the energetic costs of these mechanisms and evaluate their long-term viability under salt stress.
Osmolytes that maintain turgor
Polyamines
-
Polyamines encompass a category of organic compounds distinguished by their structural inclusion of two or more amino groups. Spermine, putrescine, alongside spermidine stand out as the most prevalent polyamines synthesized within higher plants[18]. These polyamines play a pivotal role in overseeing a variety of cellular functions crucial for normal growth, encompassing processes like cell proliferation, senescence, and signal transduction. Moreover, their influence extends to the regulation of gene expression aimed at enhancing PST in the face of abiotic stressors. Notably, polyamines operate as osmolytes by effectively neutralizing ROS and modulating ion channels[19]. According to Fariduddin et al., high endogenous concentrations of polyamines help plants tolerate extreme saline stress. There is some evidence that using exogenous polyamines on plants can partially reverse the decrease in photosynthetic efficiency caused by salinity. However, this effect is very dependent on both the amount and type of polyamines and the level of stress[20]. Furthermore, high concentrations of polyamines may result in the production of ROS during their catabolism, particularly through the action of polyamine oxidases, which can exacerbate oxidative damage instead of alleviating it[19].
Amino acids
-
Amino acids are organic compounds that serve an important part in many biological processes while also providing structure and support to cells. Some recent investigations have corroborated the evidence that the buildup of amino acids is related to SST response. Arginine, leucine, alanine, valine, serine, and Pro are some amino acids, whereas ornithine, citrulline, and gamma-amino butyric acid are critical non-protein amino acids that usually accumulate when there are stress conditions[21]. Pro is considered an important signaling molecule and osmolyte, and normally accumulates in the cytosol of the plant. It not only stabilizes the protein structure but also protects plant cells from oxidative or osmotic stress; thus, it functions as a molecular chaperone or ROS scavenger[22]. According to Shahbaz & Ashraf, Pro exogenous application has a substantial impact on increasing the endogenous level of Pro in plants, which improves growth, decreases uptake of Cl− and Na+, and improves the antioxidant defense system[23].
Arginine is also reported to ameliorate growth, particularly in Salicornia europaea L. plant seedlings, and help seedlings withstand or tolerate salt (NaCl) stress. Foliar application of arginine (5 mM) was observed to foster plant growth and SST by improving the accumulation of Pro and antioxidant defense[24]. Furthermore, the combination of arginine with manganese is more potent in mitigating the effects of excessive salts in peas. GABA is also crucial in improving SST in various plants, transported to mitochondria after being synthesized in the cytosol of the plant cell by glutamate decarboxylase[25]. In tomato plants, GABA functions to mitigate the detrimental effects of saline stress by regulating amino acid synthesis, sodium ion (Na+) absorption, alongside ROS metabolism. The treatment of 5 mmol/L GABA on tomato seedlings growing under 175 mmol/L NaCl was affected in improving SST by improving endogenous GABA[26]. Moreover, it has been noted that various amino acids can contribute to reinforcing plant resilience in the presence of salinization stress. Despite the significant role of amino acids in enhancing SST, there is limited research on the synergistic effects of multiple amino acids when applied together, as opposed to individually. Additionally, the long-term physiological and metabolic trade-offs of amino acid accumulation under saline stress remain poorly understood.
Betaines
-
Glycine betaine (GB) is a non-protein amino acid that is distributed among certain Arabidopsis spp. Most of the crops do not accumulate betaines. GBs are found in the chloroplasts of plants, where they perform vital roles like protection of thylakoids and chloroplast adjustment; therefore, they contribute to maintaining photosynthetic efficiency[27]. GB is an essential osmolyte in the regulation of cellular osmotic balance and the stabilization of proteins and membranes in plant saline stress management. Moreover, the activation of stress-responsive genes is orchestrated by their control of TFs (WRKY, MYC/MYB, ABF, AP2/ERF, DREB/CBF, NAC), thereby helping plants withstand salt stress better. The dual functions of GB as a transcriptional regulator and osmoprotectant highlight how important it is for enhancing plant resistance to salinity[28].
The foliar application of GB in cotton plant seedlings effectively reduces the damaging effect of induced saline stress (NaCl). The optimal concentration of GB was 5 mM, which was effective in improving growth characteristics, photosynthetic activities, and stomatal functions. Some plant species may better withstand saline stress after receiving exogenous GB. For instance, foliar GB treatment substantially enhances rice SST[29]. Sofy et al. examined the effectiveness of the application of GB (exogenous) in different concentrations (25 and 50 mM) in improving salt stress (NaCl) tolerance on Phaseolus vulgaris L. Even with the application of 25 mM GB, the pod yields increased by 59.4% and 29.8% in P. vulgaris plants irrigated with 100 and 50 mM NaCl, respectively. Additionally, foliar application of GB also improved the uptake of K+, whereas it reduced the accumulation of Na+, thus helping in maintaining K+/Na+. The principal mechanism by which GB may improve SST is by adjusting ion intake, restricting Na+ uptake, and, conversely, boosting K+ accumulation in the tissues of plants[30]. Dustgeer et al. applied GB on maize's foliar application with concentrations of 50 and 100 mM greatly increased the leaves per plant, root and shoot growth, shoot length, stomatal conductance, membrane stability, chloroplast content, and photosynthetic rate by reducing H2O2 and malondialdehyde accumulation[31]. Despite the extensive research demonstrating the beneficial effects of exogenous GB application in enhancing SST across various plant species, several gaps remain unexplored. Firstly, the long-term impact of repeated GB applications under field conditions, particularly in crops cultivated in naturally saline soils, requires further investigation to determine its agronomic feasibility. Secondly, the physiological and molecular mechanisms underlying GB-induced enhancement of ion homeostasis and photosynthetic efficiency in certain species remain poorly understood, particularly regarding crop-specific responses.
Sugars
-
The production of stable sugars is associated with the stabilization of protein structures, ROS scavenging, carbon storage, and osmotic adjustment[32]. Sugars also participate in osmotic adjustment as main osmolytes, and about 50% of the plants' total osmotic potential is contributed by sugars. Foliar treatment of trehalose (10, 50 mM) was observed to substantially boost diverse growth parameters and organic solutes like sucrose, glucose, and trehalose in leaves in salinity-stressed wheat plants. Moreover, after trehalose treatment, a decrease in H2O2 concentration, lipid peroxidation, and lipoxygenase enzyme activity was observed. A high level of trehalose was observed to enhance the SST in the quinoa plant by increasing the photosynthetic pigments, growth, and yield parameters, and more importantly, the antioxidant enzyme activities[33].
Yuan et al. examined the effectiveness of the application of trehalose (exogenous) on seedlings of Citrullus lanatus. The amount used for the treatment was 20 mM, and an improvement in SST via enhanced expression of SST-related genes was observed. Furthermore, carbohydrate biosynthesis pathways, plant hormone signal transduction, and the accumulation of phenylpropanoids were also reported to enhance SST[34]. According to Nawaz et al.[35], the use of osmoprotectants is regarded as a crucial tactic for developing SST in plants. Trehalose has gained a lot of interest recently since it has been shown to be an effective osmolyte for promoting salinity tolerance. Trehalose enhances plant water relations and nutrient absorption, improves membrane integrity, and decreases leakage of electrolytes and lipid peroxidation under saline stress. Additionally, trehalose enhances gas exchange characteristics and shields plants against oxidative damage caused by saline stress to the photosynthetic system. In a study on Lactuca sativa L. by Zhang et al.[36], it was noticed that the application of exogenous chitosan resulted in significant improvements in various growth parameters of lettuce, including shoot fresh weight, total leaf area, as well as shoot and root dry weight. Additionally, exogenous chitosan treatment led to increased levels of leaf Pro content, chlorophyll a, and soluble sugar contents. The myo-inositol exogenous application was effective in reducing salinity-induced stress in Malus hupehensis Rehd. The exogenous application not only changed the stomal behavior but also inhibited Na+ uptake and ROS accumulation with increased antioxidant system functionality and expression of genes (SOS1, SOS2, NHX1, and NKT1) involved in Na+ uptake[37]. Despite substantial evidence highlighting the role of osmolytes such as polyamines, amino acids, betaines, and sugars in maintaining turgor and enhancing SST, several research gaps persist. The interaction and synergistic effects of multiple osmolytes when applied simultaneously remain poorly understood. Additionally, the long-term implications and sustainability of exogenous osmolyte applications under natural field conditions need further investigation to optimize their practical use in agriculture. Exploring species-specific responses and molecular mechanisms could further enhance the understanding of osmolyte-mediated stress tolerance.
While there is general consensus on the importance of osmolytes like Pro, GB, and polyamines in maintaining turgor under saline stress, some studies report varying degrees of effectiveness depending on the plant species and stress conditions[19]. Figure 2 depicts key biomolecules involved in osmoprotection and salt stress tolerance in plants, shown with their chemical structures. For instance, while Pro is widely recognized for its role in stabilizing proteins and scavenging ROS, there are contradictory reports on its overall impact on salt tolerance across species. Some research suggests that Pro accumulation may not always correlate with improved stress tolerance in certain plants, such as those adapted to environments with low salinity. This discrepancy can be attributed to species-specific differences in metabolic pathways and osmolyte synthesis[38]. Future studies should investigate the species-specific roles of Pro and other osmolytes, particularly through targeted proteomics and metabolomics approaches.
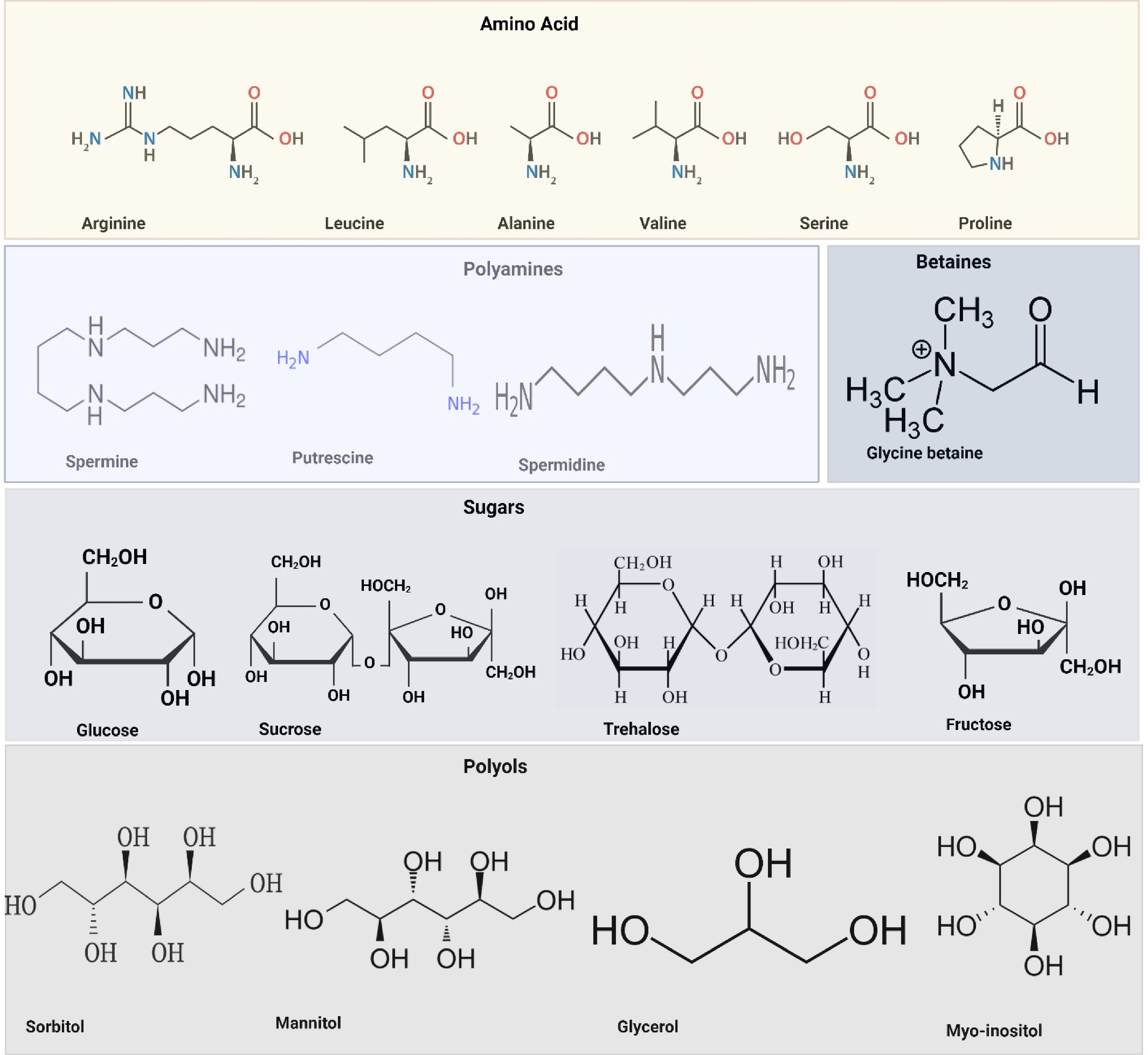
Figure 2.
Key biomolecules involved in osmoprotection and SST in plants are presented with their structural representations. Amino acids such as arginine, leucine, alanine, valine, serine, and proline play essential roles in osmotic adjustment and metabolic regulation under stress. Polyamines, including spermine, putrescine, and spermidine, contribute to ion homeostasis and oxidative stress mitigation. Betaines, such as glycine betaine, function as osmoprotectants that stabilize cellular structures under saline conditions. Sugars, including glucose, sucrose, trehalose, and fructose, serve as osmolytes and energy reserves, aiding in ROS scavenging and cellular protection. Polyols, such as sorbitol, mannitol, glycerol, and myo-inositol, help regulate osmotic balance and protect cellular integrity during abiotic stress. These biomolecules collectively enhance plant resilience by contributing to SST mechanisms. Created with BioRender.com.
https://BioRender.com/tgrndjg .Antioxidant defense
-
Plants employ an antioxidant defense mechanism as a safeguard against the oxidative stress triggered by salinity. Elevated concentrations of salt may cause oxidative damage, leading to the accumulation of ROS that damages the cellular components of plants[4]. Fortunately, plants develop antioxidant defense mechanisms to deal with these kinds of problems which, exert a substantial effect on metabolic and physiological processes. The increased production of antioxidant enzymes, e.g., (1) CAT, (2) SOD, and (3) POD as a part of antioxidant defense, is the response of the plants to ROS accumulation. These enzymes are well-known for their ROS-scavenging[29]. These enzymes with antioxidant potential are harbored in different sites of the plant cells and perform their role in detoxifying ROS. Moreover, some other antioxidant compounds (carotenoids, flavonoids, ascorbate, tocopherol, and glutathione) with low molecular mass are also critical for their role in ROS homeostasis in plant species[29].
Enzymatic antioxidants
-
Plants employ enzymatic antioxidants to neutralize reactive oxygen species through coordinated high-turnover reactions, beginning with catalase (CAT) in peroxisomes and glyoxysomes, which decomposes hydrogen peroxide into water and oxygen at a rate of roughly six million molecules per minute to prevent harmful accumulation[39]. Superoxide dismutase (SOD) isoforms located in mitochondria, peroxisomes, plastids, the cytosol, and chloroplasts convert superoxide radicals into hydrogen peroxide and oxygen throughout cellular compartments, and the hydrogen peroxide thus formed is scavenged by the ascorbate-glutathione cycle. In this cycle, ascorbate peroxidase (APX) reduces hydrogen peroxide to water using ascorbate as an electron donor, monodehydroascorbate reductase and dehydroascorbate reductase regenerate ascorbate from its oxidized forms using NADPH and reduced glutathione, respectively, and glutathione reductase restores reduced glutathione from its disulfide form, thereby sustaining continuous detoxification of ROS. Additionally, class III peroxidases (POD) in the apoplast use H2O2 to oxidize phenolic compounds, reinforcing cell walls and further mitigating oxidative stress[40].
Non-enzymatic antioxidants
-
Flavonoids are the secondary metabolites chiefly involved in the pigmentation of fruits, flowers, stems, and roots. Derived from malonyl-coenzyme A and phenylalanine, chalcone is the precursor for flavonoids, which are aromatic compounds. Flavonoids can be categorized based on chemical structure into anthocyanins, flavans, flavanones, flavanediols, flavones, isoflavones, and condensed tannins[41]. Flavonoids play a key role in antioxidant defense by inhibiting ROS-generation enzymes, thus preventing ROS increase. ROS-generating enzymes are lipoxygenase, monooxygenase, cyclooxygenase, and xanthine oxidase, which are inhibited by the flavonoids and thus involved in antioxidant defense. Furthermore, flavonoids also protect plants' cellular compartments against some dangerous oxidizing pollutants like sulfur dioxide (SO2) and ozone (O3) by serving as antioxidant barriers. Compared to ascorbic acid or alpha-tocopherol, flavonoids have an antioxidant capacity many times higher because of their great ability to donate H-atoms or electrons[42].
Ascorbic acid is a well-known non-enzymatic antioxidant in plants that acts as a cofactor or redox buffer for different enzymes. Ascorbic acid has a structure that is similar to that of hexose sugars; specifically, it has a conjugated structure that is made up of a 5-carbon lactone ring and contains enediol groups on carbon atoms two and three. The ability of ascorbic acid to donate electrons to a variety of electron donors in non-enzymatic and enzymatic reactions makes it the most powerful scavenger of ROS[43]. In addition, it supports violaxanthin deepoxidase's ability to dissipate excessive excitation energy in chloroplasts by acting as a cofactor for that enzyme. Ascorbic acid interacts with tocopherol, superoxide, hydroxyl radicals, and oxidized glutathione due to its redox potential. Tocopherols fall into the class of lipophilic compounds and also exhibit important antioxidant properties. They are non-enzymatic antioxidants that possess the ability to inhibit lipid peroxidation, whereas tocopherols neutralize the damaging effect of free radicals by donating hydrogen atoms and thus act as antioxidants[44].
In an experiment on onions, it was found that foliar spray of α-tocopherol enhances SST through an improved level of antioxidant defense. Exogenous application of α-tocopherol reduced lipid peroxidation and endogenous H2O2 while increasing enzymatic activity (APX, SOD, GR, and CAT) and non-enzymatic activity (glutathione and ascorbic acid)[45]. Sunflower seeds were immersed in α-tocopherol at four concentration levels (100, 200, 300 mg/L, and distilled water as control) for 14 h before sowing in the soil and irrigating with 0 and 120 mM NaCl concentrations until the final harvest. The treated plant with α-tocopherol showed an increased level of enzymatic antioxidant activity (APX, SOD, GR, and CAT) and increased root and shoot dry weight. The SST was reported at its maximum with two α-tocopherol concentrations, e.g., 200 and 300 mg/L[46].
Glutathione acts as a fundamental non-enzymatic antioxidant, pivotal for regulating redox balance through the scavenging of ROS, consequently serving as a cornerstone in antioxidant defense strategies[29]. The role of glutathione ranges from metabolite conjugation, stress-responsive gene expression, xenobiotic detoxification, and signal transduction. Glutathione was also reported to be important in controlling the level of various ROS, like ·OH and O2·− and H2O2 in plant cells by changing the balance between oxidized (GSSG) and reduced (GSH) forms. Therefore, this dynamic relationship provides valuable information on cellular redox potential and its regulation[47]. Figure 3 illustrates the cellular mechanisms of osmoprotection, ion balance, and ROS scavenging in plant cells.
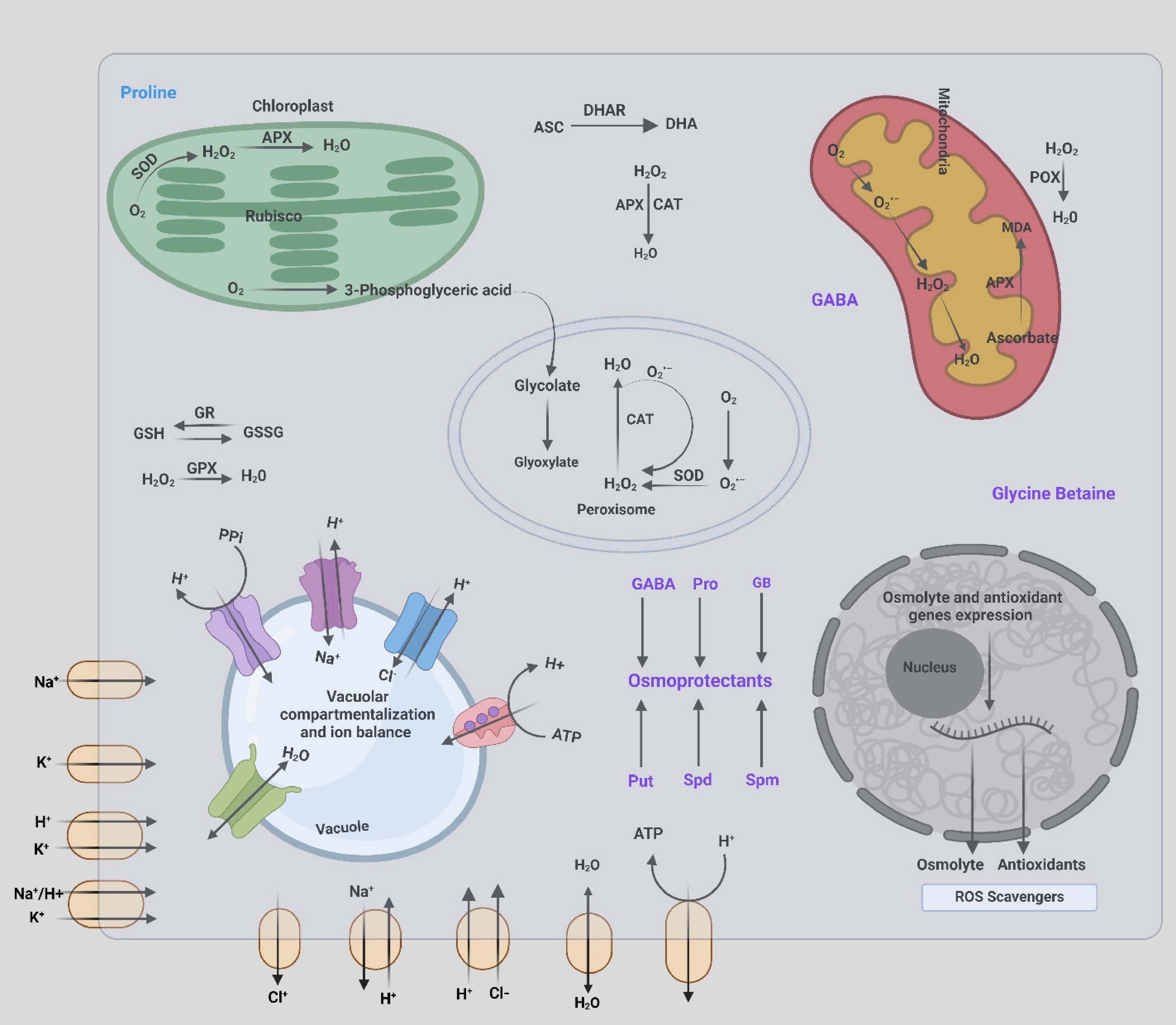
Figure 3.
Cellular mechanisms of osmoprotection, ion balance, and ROS scavenging in plant cells. The chloroplast involves SOD converting O2− to H2O2, which is further detoxified by APX to H2O. The Calvin cycle enzyme Rubisco is depicted with its substrates, O2 and 3-phosphoglyceric acid. The mitochondria show the role of ascorbate (Asc) and other antioxidants in detoxifying H2O2 through the action of POX and APX, with the production of malondialdehyde (MDA) as a marker of lipid peroxidation. In the peroxisome, CAT and SOD work together to convert glycolate to glyoxylate and detoxify H2O2 to H2O. The vacuole displays ion transport mechanisms, including Na+/H+ antiporters, K+ channels, and Cl− channels, contributing to vacuolar compartmentalization and ion balance. Vacuolar ATPase (V-ATPase) and pyrophosphatase (PPi) pump H+ into the vacuole to maintain pH and ion homeostasis. In the cytosol, the reduction-oxidation (redox) cycling of glutathione (GSH) to glutathione disulfide (GSSG) is mediated by glutathione reductase (GR) and glutathione peroxidase (GPX), with H2O2 being converted to H2O. In the nucleus, the expression of genes involved in the synthesis of osmolytes and antioxidants is regulated. Osmoprotectants such as GABA, Pro, and GB contribute to osmoprotection alongside antioxidant production. These interconnected processes integrate osmoprotection, antioxidant defense, and ion balance to maintain cellular homeostasis and protect plant cells from oxidative damage under salinity stress. Created with BioRender.com.
https://BioRender.com/8nvvmfx .Known for their pivotal role in photosynthesis, carotenoids also act as a shield, protecting plants from the detrimental effects of photo-oxidation. Carotenoids are derived from isoprene and consist of eight isoprene units and possess hydrophobic properties[48]. The plastid is the site where carotenoids are synthesized from dimethylallyl diphosphate and isopentenyl diphosphate[49]. Li et al. found that overexpression of three genes involved in the biosynthesis of carotenoids improved the level of SST in tobacco plants. Therefore, it was concluded that under salt-stress environments, plants can produce and store more carotenoids, resulting in greater oxidative stress resistance through increased antioxidant enzyme production and activity[50]. While extensive research has shed light on the role of enzymatic and non-enzymatic antioxidants in combating oxidative stress under saline conditions, several areas require deeper exploration. For example, there is limited understanding of how these antioxidant systems interact dynamically within different cellular compartments to achieve optimal ROS balance. Moreover, the potential trade-offs between antioxidant activity and energy allocation in plants under prolonged salinity stress remain largely unexamined. Future research should also emphasize uncovering the antioxidant responses of lesser-studied plant species and the molecular mechanisms driving their resilience, which could pave the way for more targeted strategies to enhance SST in crops.
The manuscript emphasizes the critical role of enzymatic antioxidants such as CAT, SOD, and APX in managing ROS under salt stress[4]. However, some studies argue that non-enzymatic antioxidants (e.g., ascorbate, tocopherol, and flavonoids) may play a more significant role in ROS detoxification than enzymatic antioxidants, particularly in species that lack efficient enzymatic defense systems[51]. For example, while CAT is known to be a powerful antioxidant, certain plants might exhibit enhanced tolerance due to the increased activity of non-enzymatic antioxidants rather than enzymatic pathways. These contrasting findings underscore the need for a comprehensive approach that considers the balance between enzymatic and non-enzymatic antioxidant systems. Research that integrates both systems in proteomic and metabolic analyses would be invaluable to better understand their synergistic interactions and relative contributions to SST[51].
Ion transporters and hormonal regulation in salt stress tolerance
-
To mitigate the adverse effects of salinity, plants have evolved highly specialized mechanisms involving ion transporters and hormones, which work together to ensure cellular stability and survival in saline environments[52].
Ion transporters in salt stress adaptation
-
One of the key components in salt stress tolerance is the regulation of ion transport across cellular membranes. Plants have developed a variety of ion transporters that actively manage the influx and efflux of ions, thereby maintaining cellular homeostasis under high salinity conditions. Among the most critical ion transporters are Na+/H+ antiporters, such as SOS1, which play a vital role in sodium (Na+) exclusion from the cytoplasm. SOS1 pumps excess Na+ ions out of the cytoplasm into the apoplast, thereby protecting cellular machinery from Na+ toxicity[53]. Similarly, HKT1 (High-affinity K+ Transporter 1) is involved in the retrieval of K+ ions from the soil solution, ensuring the proper functioning of the plant's ion balance by maintaining potassium (K+) levels, which are essential for enzyme activity and membrane potential[54]. Another crucial transporter is the vacuolar Na+/H+ exchanger (e.g., NHX1), which is involved in compartmentalizing Na+ ions into vacuoles, where they do not disrupt critical cellular processes. This sequestration of Na+ into vacuoles reduces the cytoplasmic Na+ concentration, maintaining enzymatic functions and stabilizing metabolic processes. The coordinated action of these ion transporters not only prevents ionic toxicity but also aids in maintaining osmotic balance and turgor pressure, both of which are essential for plant survival under saline stress[55]. Furthermore, recent studies have highlighted the intricate regulation of these transporters under salt stress. They are tightly controlled by signaling pathways, including those mediated by ABA (abscisic acid), which modulates the expression of ion transporters in response to elevated salt levels. This regulatory network allows plants to fine-tune their ion homeostasis strategies according to the severity of salt stress, ensuring optimal energy use and preventing cellular damage[52].
Hormonal regulation and its role in salt stress tolerance
-
Hormones serve as essential regulators of plant stress responses, and their involvement in salt stress tolerance is of paramount importance. Abscisic acid (ABA), in particular, plays a central role in orchestrating plant responses to osmotic stress, including salt stress. Upon perceiving elevated salinity, ABA accumulates and activates several stress-related genes, including those involved in ion transport, osmotic adjustment, and the antioxidant defense system[56]. For example, ABA enhances the expression of SOS1, HKT1, and NHX1, all of which contribute to ion homeostasis. Additionally, ABA promotes stomatal closure, reducing water loss through transpiration, which is crucial for maintaining hydration during periods of high salinity[57].
In addition to ABA, other hormones such as cytokinins, auxins, and gibberellins also influence salt stress responses. Cytokinins, known for promoting cell division and growth, play a vital role in maintaining root growth under salt stress by regulating the expansion of root cells, which allows the plant to access water and nutrients[57]. Studies have shown that cytokinin-treated plants exhibit enhanced salt tolerance by improving root architecture and reducing salt-induced damage to root tissues. On the other hand, auxins help coordinate cell elongation and growth, particularly in the root system, enabling better adaptation to soil salinity[58]. Gibberellins influence seedling growth, and their regulation under saline conditions helps ensure proper seedling establishment. Crosstalk between ABA and other phytohormones is crucial for integrating stress responses and fine-tuning plant growth[59]. For instance, ABA can interact with gibberellins and auxins to regulate plant growth, ensuring that the plant does not expend energy on unnecessary growth processes when under stress. This hormonal interplay is essential for the plant to prioritize stress mitigation, such as ion homeostasis and osmotic regulation, over growth-related processes[56].
-
Signal transduction pathways activated by salt stress perceive and transmit stress signals to transcription factors, which in turn modulate the expression of target genes to enhance plant tolerance.
Early perception, signal transduction cascades
-
The perception of salt stress in plants begins at the plasma membrane, where the accumulation of salt ions, particularly sodium (Na+), triggers a cascade of signaling events. One of the primary mechanisms for early salt stress detection is the Salt Overly Sensitive (SOS) pathway. SOS3, also known as CBL4, is a calcium-binding protein that acts as a sensor for increased Na+ concentrations in the cytoplasm. When salt stress occurs, SOS3 interacts with SOS2, a protein kinase, to form a complex that activates SOS1, a Na+/H+ antiporter responsible for the efflux of Na+ from the cell. This early perception mechanism is essential for protecting cellular integrity by preventing the excessive buildup of Na+ in the cytoplasm[60].
Once salt stress is perceived, it triggers a series of intracellular signaling cascades that help the plant adapt to the harsh conditions. The initial recognition of salt stress involves changes in the ion concentrations within the plant, particularly the elevated levels of sodium ions (Na+) in the extracellular space. These changes are sensed by specific receptors in the plant cell membrane, which activate a cascade of signaling events[61]. A critical pathway involved in this process is the calcium (Ca2+) signaling pathway, which plays a pivotal role in transducing the salt stress signal from the plasma membrane to the nucleus. The accumulation of Na+ in the cytoplasm leads to an increase in intracellular Ca2+ levels. This increase in calcium concentration serves as a key signal to activate downstream responses[62]. Calcium-dependent protein kinases (CDPKs) are one of the primary sensors of this Ca2+ influx. Upon binding Ca2+, CDPKs undergo a conformational change that allows them to phosphorylate specific target proteins, thereby propagating the signal within the plant cell. This phosphorylation cascade amplifies the initial signal and initiates a broader cellular response to salt stress[63].
Another crucial component of salt stress signaling is the activation of mitogen-activated protein kinases (MAPKs). MAPKs act as secondary messengers in response to the rise in intracellular Ca2+. These kinases are activated through a series of phosphorylation events and trigger a cascade of intracellular signaling that leads to the activation of various stress-responsive pathways. MAPKs play a central role in regulating a wide variety of plant responses to environmental stresses, including salinity[64]. The activation of MAPKs results in the phosphorylation and activation of transcription factors (TFs), such as WRKY, NAC, bZIP, and GATA. These TFs are pivotal in regulating the expression of genes that help the plant mitigate the damaging effects of salt stress (Table 1). Moreover, WRKY TFs are involved in regulating genes associated with ROS (reactive oxygen species) detoxification, ion transport, and oxidative stress responses, while NAC and bZIP TFs help activate genes related to osmotic regulation and cellular integrity under stress conditions[65].
Table 1. Crucial transcription factors driving the plant salt tolerance mechanism.
Plant species Genes Gene function Ref. Ipomoea batatas IbGATA24 ROS scavenging [72] Ananas comosus AcWRKY28 Enhancing SST by upregulating the expression of AcCPK genes [73] Zea mays ZmSNAC1 Salt stress response [74] Zma001259 Involved ABA-dependent signaling pathways [75] Zma029150 Involvement in ABA-dependent signaling pathways [75] ZmMRPA6 ATP-binding cassette transporter resistance [76] Triticum aestivum TaNAC47 Salt stress response [77] TaNAC29 Reduces MDA and H2O2, increases CAT and SOD activity [78] TaNAC67 Improves chlorophyll content, sodium ion balance [79] TaNAC69 Regulation of salt stress-responsive genes [80] TaNAC13 Saline stress response [81] OsNAP Upregulation of salt stress related genes [82] OsGATA8 ROS scavenging activity and chlorophyll biosynthesis [71] Sorghum bicolor SbNAC56 Salt stress response [83] Solanum Lycopersicon SlTAF1 Accumulation of osmoprotectant Pro [84] SlNAC2 Regulation of glutathione metabolism as salt stress response [85] Glycine max GmGRAS66 Salt stress response [86] Elaeis guineensis EgAP2.25 Improve antioxidant enzyme activities [87] Mangifera indica MiCOMT genes Promote the accumulation of MeJA and ABA [88] Santalum album SaSAUR08, SaSAUR13 Respond to the auxin [89] A key feature of salt stress signaling is the dynamic interaction between multiple signaling pathways. Calcium signaling not only activates MAPK cascades but also interacts with reactive oxygen species (ROS) signaling. ROS molecules, generated during salt stress, act as secondary messengers that amplify the stress signal and initiate further responses. These ROS-mediated signaling processes also activate transcription factors such as WRKY and NAC, which are involved in regulating the expression of genes responsible for maintaining cellular integrity and protecting against oxidative stress[66]. Figure 4 depicts the signal transduction pathway mediating the saline stress response in plants.
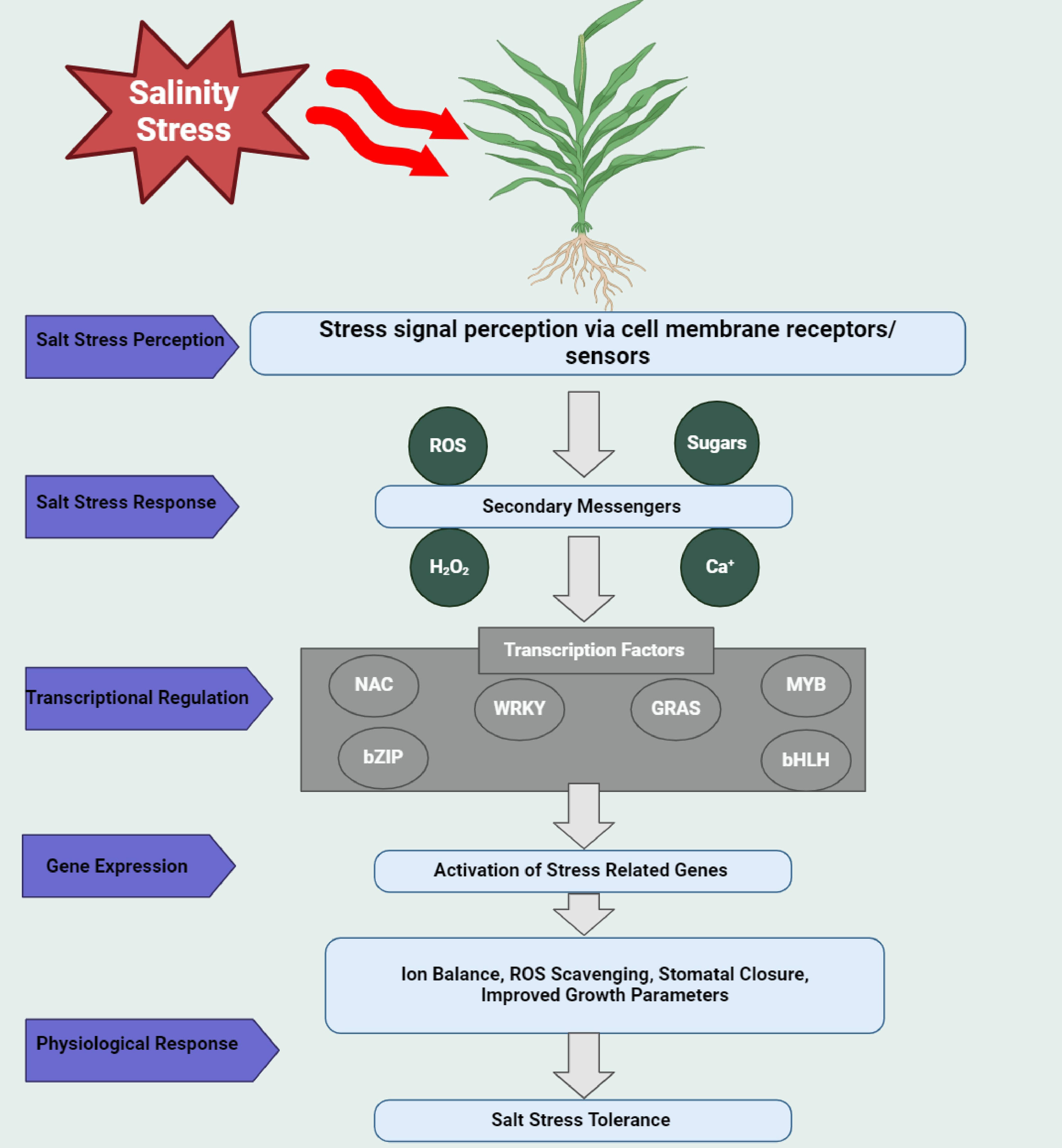
Figure 4.
Signal transduction pathway involved in saline stress response in plants. Salt stress triggers a complex signaling cascade that enables plants to perceive, respond, and adapt to high salinity conditions. The process begins with salt stress perception via membrane-bound receptors and ion transporters, including SOS3 (CBL4), NHXs (Na+/H+ exchangers), and HKT1, which detect excessive Na+ accumulation. These sensors activate secondary messengers such as Ca2+, ROS, sugars, and H2O2, which amplify the stress signal and initiate downstream responses. The SOS signaling pathway (SOS3-SOS2-SOS1) is critical for Na+ efflux, maintaining ion homeostasis, while Ca2+ signaling cascades activate MAPKs and calcium-dependent protein kinases (CDPKs) to further regulate stress responses. These signaling molecules activate TFs (NAC, WRKY, GRAS, MYB, bZIP, and bHLH), which regulate the expression of stress-responsive genes involved in ROS scavenging (e.g., SOD, CAT, POD), osmolyte biosynthesis (e.g., P5CS, RD29B), and ion transport (e.g., NHX1, HKT1;5). The coordinated activation of these genes leads to key physiological responses, including ion homeostasis, ROS detoxification, stomatal regulation, osmotic adjustment, and enhanced biomass accumulation, allowing plants to withstand salinity stress. Additionally, hormonal crosstalk, particularly with ABA, fine-tunes these responses by integrating environmental cues with developmental programs. This intricate and highly coordinated signaling network ensures plant survival in saline environments by activating molecular defense mechanisms that protect cellular integrity, maintain metabolic balance, and enhance SST. Created with BioRender.com.
https://BioRender.com/p9q2tl4 .Transcriptional regulation/dynamics
-
When extracellular Na+ rises, plant cells perceive the ionic and osmotic imbalance at the plasma membrane, triggering rapid cytosolic Ca2+ transients, bursts of reactive oxygen species, MAP-kinase cascades, and abscisic acid (ABA) accumulation. These signals converge on key transcription-factor (TF) families, most prominently WRKY, GATA, NAC, and GRAS, whose phosphorylation enhances their nuclear import or DNA-binding affinity[67]. Once in the nucleus, the activated TFs recognize specific cis-elements within target promoters and recruit chromatin remodelers and the basal transcriptional machinery, shifting the transcriptome toward defense mode. Upregulated genes encode plasma-membrane and vacuolar Na+/H+ antiporters (SOS1, NHX1/2) and K+ transporters (HKT1) that restore ion homeostasis, enzymes for osmolyte biosynthesis (proline, trehalose), and late-embryogenesis-abundant (LEA) proteins that balance cellular water potential, as well as antioxidant enzymes (SOD, CAT, APX) that scavenge stress-induced ROS; simultaneously, growth-promoting genes are often repressed to conserve energy. Feedback loops among TFs and crosstalk with other hormones such as ethylene and jasmonate refine the response until ionic and osmotic equilibrium is re-established, collectively conferring salt-stress tolerance[68]. Under salt exposure, a rapid Ca2+ spike activates the CBL4-CIPK24 (SOS3-SOS2) kinase complex, which phosphorylates multiple TFs and triggers their nuclear import[67]. WRKYs recognize the W-box (TTGAC[C/T]) in promoters of ion- and redox-related genes[68]. Under salt stress, WRKY TFs activate or repress the expression of genes involved in stress tolerance, such as ion transport and antioxidant defense. For instance, OsWRKY54 binds to the promoter of OsHKT1;5, a gene involved in K+ and Na+ transport, and activates its expression. This action helps maintain Na+ homeostasis by facilitating Na+ efflux out of the cytoplasm. Additionally, OsWRKY54 also regulates the expression of OsNHX4, a vacuolar Na+/H+ antiporter, which aids in the compartmentalization of Na+ into vacuoles. These regulatory mechanisms help improve salt tolerance by reducing Na+ toxicity and maintaining cellular ion balance[69].
GATA TFs have a crucial function in enhancing the response to saline stress in different plant species. GATAs are categorized into four subfamilies (A–D) according to their domain architectures, evolutionary relationships, exon-intron structure, as well as zinc loop motif[70]. However, the B-GATAs have been extensively researched in the field of plant research. When the OsISAP1 gene, a GATA gene from rice, was overexpressed in tobacco plants, it led to enhanced resistance not only to salt but also to cold and drought stress. GATA TFs are involved in regulating the expression of genes related to ROS scavenging, ion homeostasis, and chlorophyll biosynthesis under salt stress. OsGATA8, a key GATA TF in rice, activates the expression of genes involved in ROS elimination, such as SOD and CAT, helping to reduce oxidative damage. Furthermore, OsGATA8 also regulates chlorophyll biosynthesis genes, which enhances photosynthetic efficiency under stress. The activation of these genes by GATA TFs helps maintain cellular integrity and supports growth under saline conditions. An example of the target gene expression pattern is the upregulation of OsGATA8 under both salt and ABA stress, leading to the enhanced scavenging of ROS and improved plant growth[71]. In sweet potato, the IbGATA24 gene extracted from the Xushu55-2 line, known for drought tolerance, demonstrates significant induction under various abiotic stresses. Its overexpression in transgenic Arabidopsis plants triggers heightened drought and SST by stimulating the ROS elimination mechanism and hormonal signaling pathways. The presence of IbCOP9-5a was also detected throughout the salt treatment, indicating a robust reaction to challenging conditions. Transgenic plants that have been genetically modified to overexpress the IbCOP9-5a gene exhibit enhanced fresh weight, chlorophyll content, and survival rate when subjected to salt stress conditions. After subjecting the plants to stress treatment, the transgenic lines exhibited a considerable decrease in H2O2 content, MDA content, and electrolyte leakage compared to the WT[72].
Saidi et al. demonstrated that the BnGATA genes are expressed to develop tolerance to many abiotic stressors, including salt, cold, drought, and ABA. The genes like BnGATA3.1, BnGATA2.23, and BnGATA1.27 were shown to be elevated in response to saline stress. Nine GATA family genes (SlGATA28, SlGATA11, SlGATA2, SlGATA25, SlGATA13, SlGATA7, SlGATA17, SlGATA22, and SlGATA9) showed elevated expression when subjected to saline stress[90]. Similarly, treatments for ABA stress, salt, and drought all dramatically increase the expression of OsGATA26 in rice, a homolog of SiGATA18. These findings suggested that GATA domains have a significant part in responses to stress, specifically against salt[91].
NAC TFs are implicated in mediating plant responses to various abiotic stresses, often through intricate regulatory networks involving hormone signaling pathways. Notably, ABA-dependent and ABA-independent mechanisms have been implicated in the induction of NAC gene expression under stress conditions[92]. NAC TFs are involved in stress-induced gene expression through both ABA-dependent and ABA-independent pathways. Salt-induced NACs (e.g., OsNAC6) bind the NAC-recognition sequence (CACG) to drive osmolyte- and energy-metabolism genes[93]. The OsNAC6 regulates the expression of genes involved in nicotinamide biosynthesis, which is critical for cellular energy homeostasis under salt stress[94]. In Arabidopsis, ANAC019, ANAC055, and ANAC072 are involved in the regulation of ERD1 (early responsive to dehydration 1), a gene that encodes a chloroplast ATP-dependent protease, which helps in stress adaptation by managing protein turnover. The NAC TFs promote the expression of stress-responsive genes that assist in maintaining protein integrity and cellular function under high salinity conditions[95].
GRAS TFs are also critical in making plants more resilient to the saline stress challenge. Several prior investigations have shed light on the functions of GRAS in response to saline stress. For example, the PeSCL7 (a GRAS gene) gene, in Populus euphratica, shows increased expression levels in response to salt and drought treatments. The expression of the PeSCL7 gene was observed in Arabidopsis thaliana, resulting in increased resistance to salt and drought conditions[101]. VaPAT1 overexpression in Arabidopsis led to higher expression of stress-related genes, including AtSIZ1, AtCBF1, AtATR1/MYB34, AtMYC2, AtCOR15A, AtRD29A, and AtRD29B, compared to wild-type plants under normal growth conditions, indicating its significant role in regulating stress-responsive pathways[102]. The response of cassava genes to a range of environmental stressors, including salt, was assessed through RT-qPCR analysis. The expression of MeGRAS2, MeGRAS32, MeGRAS12, MeGRAS22, and MeGRAS63 was upregulated in response to saline stress treatments, as reported by Shan et al.[103]. The expression patterns of these target genes often exhibit a clear upregulation under salt stress, reflecting the activation of TFs like WRKY, NAC, and GATA in response to salinity. These TFs typically function in conjunction with other signaling pathways, including those involving hormones like ABA, which further enhance the expression of stress-related genes. For instance, the expression of OsHKT1;5 and OsNHX4 is tightly regulated by ABA-mediated pathways, with WRKY and NAC TFs acting as critical regulators to fine-tune the plant's response to salinity. Thus, the coordinated action of these TFs ensures that plants efficiently cope with salt stress by modulating a network of genes involved in ion homeostasis, ROS detoxification, and osmotic regulation. The bZIP TFs (basic leucine zipper) regulate stress-responsive genes by binding to specific ABRE (ABA-responsive element) motifs in gene promoters. Under salt stress, bZIP TFs activate genes involved in osmotic adjustment and ion transport. For example, ABF1, a bZIP TF, regulates the expression of genes such as RD29B, a gene involved in osmotic stress response. The activation of these genes helps the plant to adjust its osmotic potential and maintain water balance under saline conditions.
This manuscript discusses the roles of key TF families (e.g., NAC, WRKY, bZIP, GATA) in the regulation of salt stress-responsive genes. However, the literature reveals some conflicting findings regarding the functional redundancy and specificity of these TFs. For instance, while NAC and WRKY TFs are widely reported to regulate stress-related gene expression, studies have shown that their roles may overlap significantly, complicating the identification of their unique contributions. Moreover, the regulatory interactions between these TFs and the various signaling pathways (ABA, ROS, calcium, etc.) are not fully understood, particularly regarding species-specific responses. A critical analysis of how TFs interact with other signaling molecules and their potential redundancy in stress responses is necessary to clarify these contradictions. Furthermore, more research is needed to explore the role of lesser-known TFs in salt stress adaptation, which could reveal novel targets for crop improvement.
-
In recent years, advancements in omics technologies like genomics, proteomics, transcriptomics, and their integration have opened new avenues to unravel the intricate mechanisms of SST in plants (Table 2). By adopting a multi-omics approach, researchers are now able to bridge the gap between molecular, cellular, and physiological responses, offering a systems-level understanding of how plants adapt to saline environments. These insights are instrumental in identifying potential genetic, biochemical, and regulatory targets for developing salt-tolerant crop varieties, which is critical in addressing agricultural productivity under increasing SST and environmental stress[104].
Table 2. Some successful breeding and genetic engineering efforts to enhance salt tolerance in crops.
Crop species Variety/line Gene(s) introduced/
modifiedBreeding/genetic
engineering approachImprovement in SST Ref. O. sativa IR64-Saltol OsHKT1;5 Marker-assisted breeding (MAB) Reduced Na+ accumulation, better root growth [96] S. lycopersicum Transgenic line SlTAF1 Agrobacterium-mediated Transformation (transgenic) Increased proline content, improved biomass [97] T. aestivum Transgenic wheat TaNAC29 CRISPR-Cas9 gene editing Increased antioxidant enzyme activity (SOD, CAT) [98] Q-18 (salt-tolerant line) Natural genotypic
variationsSelective breeding Efficient osmotic adjustment, increased ion compartmentalization [99] Gossypium hirsutum Transgenic cotton GhNAC79 Gene overexpression
(transgenic)Increased antioxidant defense, reduced oxidative stress [100] Genomics: deciphering the blueprint of salt tolerance
-
Genomics, the comprehensive study of an organism's entire DNA sequence, serves as the bedrock for understanding the genetic foundations of SST in plants[104]. As salinity becomes an increasing global challenge to agriculture, particularly in arid and semi-arid regions, identifying and manipulating genes that govern plant responses to salt stress is crucial for crop improvement[105]. By harnessing advanced genomic tools such as high-throughput sequencing technologies like next-generation sequencing (NGS), whole-genome sequencing (WGS), and transcriptomics (RNA sequencing), scientists have made significant strides in mapping the genetic networks involved in SST. These methods provide an in-depth view of the plant genome and transcriptome, enabling the identification of key genes, quantitative trait loci (QTLs), and epigenetic regulators involved in plant responses to environmental stresses like salinity[106].
The genetic basis of SST is complex and multi-layered, involving a variety of mechanisms that help plants cope with excess salts. In particular, ion homeostasis, osmotic regulation, and the scavenging of ROS are critical physiological processes that plants must manage under salt stress[104]. Through WGS and RNA-Seq technologies, several candidate genes associated with essential functions in salinity tolerance have been identified, offering a blueprint for understanding how plants manage salinity at the molecular level. The ultimate goal is to identify genetic targets that can be manipulated to improve SST in crops, ensuring food security in saline-affected regions[107]. One of the most transformative approaches in genomics is Genome-Wide Association Studies (GWAS), which allow researchers to link genetic markers to specific phenotypic traits. Through GWAS, genetic loci associated with desirable traits such as improved root structure, enhanced ion compartmentalization, and better photosynthetic efficiency under salt stress have been identified[108]. These traits are fundamental to improving crop performance in saline environments. By linking these traits to genetic markers, molecular markers can now be developed to assist in selecting salt-tolerant plants for breeding programs. GWAS has proven particularly useful in understanding the genetic architecture of complex traits like salt tolerance, where multiple genes with small effects collectively contribute to the overall phenotype[109].
CRISPR-Cas9, a revolutionary gene-editing tool, has further accelerated the pace of genomics in crop improvement. With CRISPR, precise modifications can be made to plant genomes, directly targeting salt-tolerance genes such as OsHKT1;5 or OsNHX1[110]. For example, through CRISPR-mediated editing, salt tolerance has been successfully enhanced in crops like rice, wheat, and maize by knocking out or modifying the expression of specific genes. The precision and efficiency of CRISPR allow for the development of salt-tolerant crop varieties that could help address food security challenges in saline-affected areas. The ability to fine-tune salt-tolerance genes offers a more sustainable approach compared to traditional breeding methods, which are often slow and labor-intensive[111]. Beyond identifying and editing individual genes, epigenomics has emerged as an essential area of study for understanding how environmental factors like salt stress can induce heritable changes in gene expression[112]. Recent high-throughput genomics and CRISPR screens are rapidly unearthing fresh gene targets that can be edited or selected to improve salinity tolerance in major crops. Key recent GWAS hits include OsWRKY53 and OsMKK10.2, which form a regulatory module that limits root Na+ influx; loss-of-function oswrky53 mutants show 35% greater survival at 120 mM NaCl, whereas osmkk10.2 knockouts are hypersensitive[113]. A combined pan-genome with eQTL pipeline identified STG5 (LOC_Os05g49700) as a master regulator that directly activates multiple OsHKT transporters; CRISPR null mutants accumulate 1.6-fold more Na+ than WT[114]. Knocking out the rice bHLH transcription factor OsbHLH024 (A91 CRISPR mutant) boosts salt tolerance by increasing chlorophyll and antioxidant activity, lowering shoot Na+ while raising K+, and up-regulating OsHKT1;3, OsHAK7, and OsSOS1, indicating OsbHLH024 is a negative regulator of salinity response[115]. CRISPR/Cas9 deletion of six ABA-induced transcriptional repressors (GmAITRs) in soybean produces Cas9-free mutants with greater ABA sensitivity but markedly improved salt tolerance from germination to field performance, establishing GmAITRs as negative regulators and practical genome-editing targets for salinity resilience[116]. The development of pan-genomics, a field that seeks to map the genetic diversity across entire species, has also provided valuable insights into the genetic mechanisms underlying salt tolerance. Pan-genomic studies have shown that different plant species harbor distinct sets of genes and genetic variants associated with salt tolerance[106].
Proteomics: elucidating protein dynamics in salt stress
-
Understanding how plants tolerate salt stress depends much on proteomics, the broad study of proteins. Since they perform most of the cellular tasks, from enzymatic activity to structural support, proteins are fundamental to almost all biological events. Since proteins are the effectors of cellular biochemical and physiological responses, a proteomic study offers direct understanding of the molecular processes underpinning SST in plants. Protein abundance, changes, and interactions provide a whole picture of how plants observe, react to, and navigate saline conditions[4].
With to developments in tandem mass spectrometry (MS/MS), isobaric tags for relative and absolute quantification (iTRAQ), and label-free quantification, proteomic technologies have made tremendous strides recently[117]. These techniques help to identify and analyze many proteins from challenging samples. Proteomics may provide priceless information on the expression dynamics of salt-responsive proteins by means of the ability to profile proteins across many experimental circumstances, therefore exposing which proteins are upregulated or downregulated under salt stress. This degree of detail provides understanding of how salinity affects the molecular-level adaptation of plants[118]. Plants reduce salt stress mainly by controlling ROS, harmful byproducts of cellular metabolism that build up under stress. Key antioxidant enzymes, include SOD, CAT, and APX, identified by proteomics, exhibit varying expression in response to salt stress[118].
Proteomic profiling has also shed light on how plants under salt stress control their photosynthetic apparatus. Salt tolerance has been linked in species like maize to the stability of photosynthetic proteins, including those involved in the electron transport system and the light-harvesting complex[119]. Under salt stress, proteins such as RuBisCO (ribulose bisphosphate carboxylase/oxygenase), the enzyme in charge of carbon fixation, also display changed expression, potentially to aid in maximizing carbon absorption under suboptimal circumstances[120]. Beyond pointing up salt-responsive proteins, proteomics has shown that post-translational modifications (PTMs) significantly control protein activity and coordinate stress responses. In reaction to salt stress, PTMs including phosphorylation, ubiquitination, and acetylation may change the function, stability, or localization of a protein, therefore adding even another level of control[121]. Phosphorylation of TFs, like NAC and WRKY TFs, for instance, improves their stability and DNA-binding affinity. This change ensures that stress-responsive genes are triggered in the correct tissues and at the proper moment, hence optimizing the transcriptional response to salt stress. PTMs may also control protein-protein interactions, hence allowing the creation of protein complexes cooperating to control cellular responses to stress[122]. Integrated omics of salt-stressed soybean uncovered over 5,800 DEGs that reinforce cutin-suberine barriers, while proteomics flagged 312 salt-responsive proteins, most notably 2.4-fold up-accumulation of DNA-methylation enzymes (GmMET1, GmDRM2) and a 2.6-fold rise of glycolytic enolase (GmENO2) acting as a metabolic hub[123]. Proteomic profiling of the Arabidopsis agb1-2 G-protein β-subunit knockout under NaCl revealed 27 salt-responsive spots (seven identified), showing that loss of AGB1 alters proteins tied to cellulose synthesis, chromosome/DNA maintenance, organellar RNA editing, and indole-glucosinolate pathways. These shifts outline candidate effectors through which AGB1 modulates root-to-shoot Na+ movement and provide targets for improving salinity tolerance in crops[124].
The integration of multi-omics approaches (genomics, transcriptomics, proteomics) has significantly advanced our understanding of SST, but there are still critical gaps in the comprehensive mapping of salt-tolerant genes across diverse plant species. While large-scale genomic studies such as Genome-Wide Association Studies (GWAS) have identified numerous loci linked to SST, the functional validation of these loci remains an ongoing challenge. Furthermore, although CRISPR-Cas9 and other gene-editing tools have made it possible to manipulate key salt tolerance genes in model crops, the effectiveness of these modifications under field conditions, especially in polygenic traits like salt tolerance, is still poorly understood. Additionally, while the application of multi-omics strategies has provided insights into SST at the molecular level, the integration of these datasets to form cohesive models of plant response to salinity remains a major challenge. Future research should focus on improving the integration of multi-omics data and developing predictive models for gene-environment interactions under salt stress conditions.
-
This comprehensive review sheds light on the intricate interplay of biochemical and transcriptional regulation that plants employ to counter the adverse effects of saline stress. The disruptive impact of salt stress on water balance and ion levels disrupts essential functions and hampers growth. However, plants respond with remarkable strategies like biochemical adaptations, including osmotic regulation and antioxidant defense mechanisms, which act as cornerstones in maintaining cellular integrity and combating oxidative stress induced by high salt levels. The heightened production of POD, SOD, and CAT across various cellular compartments serves as a critical defense mechanism against ROS in plants. This enhanced synthesis enables efficient ROS detoxification, thereby safeguarding cellular integrity and promoting SST. The intricate regulatory mechanisms governing transcriptional reactions to salt stress highlight the adaptive mechanisms that plants implement in order to cope with detrimental environmental circumstances. Biotechnology may use TFs like NAC, bZIP, and WRKY to modify crops for improved stress-responsive gene expression. Transgenic methods using stress-inducible promoters may precisely regulate the expression of antioxidant enzymes and osmolyte biosynthetic genes, enabling crops to adapt efficiently to varying stress conditions. The overexpression of GATA and NAC TFs has shown considerable enhancements in SST in rice, wheat, and several other crops, offering a means to extend these findings to other commercially relevant species. New studies can explore novel genetic engineering techniques to enhance the expression or activity of POD, SOD, and CAT genes in crops, thereby bolstering their resilience to oxidative stress under salt-affected conditions. Additionally, further investigation into the regulatory mechanisms governing the transcriptional activation of these antioxidant enzymes can unveil new targets for crop improvement strategies aimed at enhancing SST in agriculturally important plant species. Moreover, specialized ion transporters like SOS1, HKT1, and NHX1 actively regulate the flux of Na+ and K+ under saline conditions, protecting cellular homeostasis. Likewise, hormonal networks, particularly ABA, fine-tune the expression of these transporters and other stress-responsive genes, ensuring an adaptive balance between growth and stress mitigation.
A deeper understanding of the biochemical and transcriptional mechanisms underlying SST offers valuable opportunities for translating these insights into practical agricultural and biotechnological applications. One promising avenue is the development of salt-tolerant crop varieties through advanced breeding programs and genetic engineering. Techniques such as marker-assisted selection and CRISPR-Cas9 genome editing allow for precise modification of genes related to salt tolerance, ensuring crops can maintain productivity in saline environments. Additionally, the use of osmoprotectants like proline, glycine betaine, and trehalose offers a cost-effective strategy to improve short-term resistance to salt stress. These compounds can be applied as foliar sprays or soil amendments to enhance osmotic regulation and minimize oxidative damage, benefiting especially smallholder farmers in areas with high salinity who may not have access to genetically modified crops. Salinity stress continues to pose a significant challenge to global agriculture, threatening food security and economic stability, particularly in regions experiencing rising soil salinity. Addressing this issue requires a comprehensive approach that incorporates plant science, computational biology, and environmental science. Leveraging omics technologies, such as transcriptomics, proteomics, and metabolomics, will further elucidate the molecular pathways involved in salt stress responses. Moreover, integrating environmental modeling and soil management strategies can mitigate the impact of salinity at the ecosystem level. Such interdisciplinary research is crucial to enhancing the development of salt-tolerant crops and promoting sustainable agricultural practices in regions prone to salinity.
-
The authors confirm their contributions to the paper as follows: conceptualization, writing – original draft: Usman M; project administration: Usman M, Murtaza G, Manzoor MA; data curation: Usman M, Azam M; formal analysis: Azam M; visualization: Azam M, Yousaf N, Ahmad Z, Song C; review: Yousaf N, Ahmad Z; writing – review & editing: Song C, Murtaza G, Manzoor MA; resources and supervision: Murtaza G, Manzoor MA. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
We would like to acknowledge the efforts of Dr. Muhammad Abdullah and Dr. Iftikhar Hussain Shah for reviewing this manuscript and giving valuable suggestions.
-
The authors declare that they have no conflict of interest.
-
Received 28 October 2024; Accepted 7 July 2025; Published online 29 August 2025
-
# Authors contributed equally: Muhammad Usman, Muhammad Azam
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Usman M, Azam M, Song C, Yousaf N, Ahmad Z, et al. 2025. Integrating biochemical pathways, transcriptional regulation, proteomics, and genetic approaches for enhancing salt stress tolerance in crops. Tropical Plants 4: e030 doi: 10.48130/tp-0025-0026 

Integrating biochemical pathways, transcriptional regulation, proteomics, and genetic approaches for enhancing salt stress tolerance in crops
- Received: 28 October 2024
- Revised: 24 June 2025
- Accepted: 07 July 2025
- Published online: 29 August 2025
Abstract: Excessive salt concentrations in plants disrupt vital biochemical and physiological processes, damaging organelles like mitochondria and chloroplasts. To mitigate these effects, plants utilize osmolytes such as glycerol, sorbitol, proline (Pro), glycine betaine (GB), and gamma-aminobutyric acid (GABA), which help maintain cellular turgor and water balance under osmotic stress. Antioxidant defense mechanisms, including enzymatic (e.g., peroxidase, superoxide dismutase, catalase) and non-enzymatic compounds, are essential for salt stress tolerance (SST). These enzymes, upregulated under salt stress, detoxify reactive oxygen species (ROS) and protect cells from oxidative damage. Transcription factors (TFs) such as AP2/EREBP, WRKY, bZIP, GRAS, MYB, GATA, bHLH, and NAC regulate stress-responsive genes and are critical for plant adaptation to abiotic stresses. bZIPs enhance SST in rice, while GATAs are induced by salt stress across various species. NAC TFs interact with hormone signaling pathways like ABA to modulate gene expression in response to oxidative stress and ion imbalance. Additionally, specialized ion transporters (e.g., Na+/H+ antiporters and HKT1) and phytohormones such as ABA play pivotal roles in maintaining ion homeostasis and orchestrating stress responses under salinity. Advancements in metabolomics, proteomics, transcriptomics, and genomics have significantly advanced our understanding of the molecular mechanisms underlying salt stress tolerance. By studying biochemical pathways, protein dynamics, gene expression, and genomic variations, researchers are uncovering critical networks involved in ion homeostasis, ROS detoxification, and osmotic regulation. This review synthesizes current data, demonstrating how multi-omics approaches enhance our knowledge of salt stress adaptation and offering promising avenues for biotechnological interventions aimed at improving crop resilience.
-
Key words:
- Salt stress /
- Biochemical adaptations /
- Antioxidant defense /
- Transcription factors /
- Genetic approaches /
- Proteomics