









-
Flower bud development is an essential phase in the life-cycle of plants, marking an important transition from vegetative to reproductive growth[1]. This complex morphogenetic process is jointly regulated by internal factors and external influences such as light, temperature, and phytohormones[2,3]. The timing of flowering, the quantity and quality of flowers, and the duration of the flowering period, which directly influence the ornamental and economic value of flowers, are closely associated with flower bud development[2,4]. In ornamental plants, elucidating the molecular mechanisms underlying floral bud development can facilitate the identification of key genes involved in flowering, which aids in regulating flowering time and extending the flowering period, while also providing theoretical support for flower breeding and genetic engineering[5].
Extensive research on the annual model species Arabidopsis thaliana has established a genetic and molecular framework that integrates various signaling pathways to determine the transition from vegetative to reproductive growth. Flower development in A. thaliana involves six pathways: photoperiod, gibberellin, vernalization, autonomous, senescence, and ambient temperature, which co-regulate flower-specific genes, causing the vegetative meristem to physiologically transition into a floral meristem[6,7]. Like other angiosperms, floral development in A. thaliana begins after the floral transition, when the shoot apical meristem (SAM) shifts from producing leaves to forming flowers. In A. thaliana, each stage of floral development, namely, floral transition, floral bud initiation, and floral organ development is controlled by specific gene networks[8]. Recent research on the molecular regulatory mechanisms governing floral bud development has led to significant advancements across various non-model plant species. For example, comparative transcriptomic analyses have uncovered the novel roles for transcription factors (TFs) and hormones in the induction of flowering and the differentiation of floral buds in Prunus avium[9]. Circadian rhythms are also widely implicated in the regulation of floral bud development[10]. Transcriptome analysis of differentially expressed genes (DEGs) in Anthurium andraeanum revealed that genes associated with sucrose metabolism and auxin response play crucial roles during the process of floral bud development[11]. Furthermore, transcriptome and metabolome analyses of the calyx, pistil, and stamen differentiation stages in Cornus wilsoniana indicated that 33 MADS-box TFs, including homologs of AP1 and SEP3, may synergistically regulate the morphogenesis of floral organs[5]. Transcriptome sequencing of various stages of floral bud differentiation in Carya illinoensis identified key genes potentially involved in regulating floral bud differentiation and flowering time, as well as in determining the identity of floral meristems and organs[12]. Research on flowering regulation in lilies has primarily focused on low-temperature vernalization or photoperiod pathways, with further studies exploring the functions and mechanisms of key genes involved in these processes[13,14]. However, the molecular mechanism underlying flower development in perennial monocotyledonous bulbous flowering plants remains relatively unexplored. In particular, despite a solid literature on genetics and breeding[15], there are no reported studies focusing on floral development within Fritillaria species.
Fritillaria spp. is a genus of perennial bulbous plants belonging to the Liliaceae family, known for their significant medicinal and ornamental value. The genus Fritillaria L. used to comprise 139 species, 17 subspecies, and nine varieties[16], although the taxonomy has changed considerably since then[17]. These plants are widely distributed across temperate regions of the Northern Hemisphere, with a particularly rich abundance in countries surrounding the Mediterranean[18,19]. F. pallidiflora, a wild species, is primarily found in Xinjiang, but is extensively cultivated as a medicinal plant throughout China. Research on the annual growth and development cycle of F. pallidiflora has revealed that bulbs need to reach a critical size of 1.8–2.7 g in fresh weight and a diameter of 1.2–2.2 cm for the initiation and formation of flowers[20]. That study also indicates that it takes 4–5 years for bulbs to attain this size. Based on our experience, the optimal time for planting bulbs in the Beijing area is autumn (from September to early October, when temperatures range from 15–25 °C), as this period is ideal for rooting and initiating a stage of low-temperature dormancy. In the subsequent spring (late March, when the ground temperature is 3–5 °C), buds begin to germinate and grow rapidly, leading to flowering in April. Following the blooming period, fruits and new bulbs develop, and by mid to late June, when temperatures surpass 28 °C, the aboveground parts of the plant wither. At this point, it is necessary to remove the bulbs from the soil and store them in a dry, well-ventilated environment at 18–22 °C to complete the differentiation of regenerated flower buds (June to September). This marks the beginning of the next life cycle.
Currently, research on Fritillaria species has primarily concentrated on medicinal aspects, including chemical composition, pharmacological effects, and clinical applications[21,22], their use as a resource[23], species identification, and classification[24,25], tissue culture and in vitro morphogenesis[26,27], and a few reports on genetic diversity[28,29]. However, the mechanism governing floral bud development in fritillaries, particularly within the genus Fritillaria, remains largely unknown. In this study, a high-throughput next-generation sequencing platform was employed to sequence cDNA libraries at three different developmental stages of floral bud differentiation in F. pallidiflora. We identified and analyzed the DEGs involved in the process of floral bud differentiation. A total of 45 candidate genes potentially associated with floral bud development were discovered. These findings may offer initial insight into the molecular mechanisms underlying floral development in Fritillaria species.
-
F. pallidiflora bulbs of nearly identical size, with a diameter of approximately 1.2 cm, were freshly harvested from the ground after flowering, once the stems and leaves had withered. These were collected on June 20, 2019 from the Yumin experimental station in Xinjiang, China. From June 29 to September 21, the bulbs were stored in dry, high-temperature sterilized sand (121 °C, 0.12 MPa, for 30 min), with a relative water content of less than 1%. They were then placed in an artificial climate incubator (CLC-BIV-M/CLC404-TV, MMM, Munich, Germany), and grown at a temperature of 25 °C during the day and 18 °C at night, under a 12-h photoperiod, with a light intensity of 1,000 lux, and a relative humidity of 60%–80%.
Morphological observations
-
Floral bud development was observed through paraffin sections. A total of 15 floral buds were collected every seven days from June 29 to September 21 and stored in FAA solution (formalin: acetic acid: 50% ethanol = 5:5:90, v/v/v). Paraffin sections were obtained as previously described, and sections were stained with safranin O and fast green[30]. The images of these sections were then observed using an optical microscope (Leica DFC550; Leica Microsystems GmbH, Wetzlar, Germany).
RNA extraction, cDNA library construction, and transcriptome sequencing
-
Based on morphological analysis of floral buds, total RNA was extracted from samples of F1, F2, and F3, corresponding to the three representative developmental stages of floral buds (S1, S3, and S5). F1 represents the sample from the S1 stage, sampled after 7 d of treatment (on July 6). F2 represents the sample from the S3 stage, sampled after 49 d of treatment (on August 17). F3 represents the sample from the S5 stage, sampled after 84 d of treatment (on September 21). This was done for two independent biological replicates (each sample is labeled as F1-1, F1-2, F2-1, F2-2, F3-1, and F3-2, respectively), each consisting of three floral buds. Total RNA extraction was performed using the EASYspin Plus plant total RNA rapid extraction kit (OMEGA, Norcross, GA, USA), following the manufacturer's instructions. The concentration and purity of total RNA were assessed using a Nanodrop ND-100 spectrophotometer (NanoDrop Technologies LLC, Wilmington, DE, USA) and an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The integrity of the RNA was confirmed through agarose gel electrophoresis. The RNA-seq libraries were prepared and sequenced by Gene Denovo Co. (Guangzhou, China). Transcriptome sequencing was carried out on the Illumina HiSeqTM 4000 high-throughput sequencing platform.
De novo assembly and annotation
-
The raw reads were filtered by removing adapter sequences and low-quality sequences. The resulting clean reads were then de novo assembled using Trinity[31], and the longest transcripts were selected as the unigenes. Functional annotation was conducted by aligning these unigenes against four databases: Nr (NCBI non-redundant protein sequences,
https://blast.ncbi.nlm.nih.gov/Blast.cgi ), Swiss-Prot (a manually annotated and reviewed protein sequence database,https://www.ebi.ac.uk/uniprot/ ), KEGG (Kyoto Encyclopedia of Genes and Genomes,https://www.genome.jp/kegg ), and KOG (Clusters of Orthologous Groups of proteins,https://www.ncbi.nlm.nih.gov/KOG/ ).Identification of differentially expressed genes
-
The expression level of unigenes were estimated using the fragments per kilobase of transcriptome per million mapped reads (FPKM) method[32]. Principle component analysis (PCA) was conducted using the 'prcomp' function in R with default settings. DEGs with an expression fold change ≥ 2, and a false discovery rate (FDR) < 0.01 between sequential stages were identified using the DESeq R package[33]. K-means clustering was performed to generate the expression clusters of gene expression dynamics across three stages using the OmicShare tools (
https://www.omicshare.com/tools/ ). To identify potential genes related to floral bud differentiation, DEGs were subjected to GO classification enrichment and KEGG pathway enrichment analysis using TopGO (version 2.11)[34], and KOBAS (version 2.0)[35], respectively.qRT-PCR analysis
-
Total RNA extraction and quality control procedures were performed as described in the Materials and methods. Subsequently, cDNA synthesis was performed with the HiScript® II Q RT SuperMix for qPCR (+gDNA wiper) Kit (Novoprotein Scientific Inc., Suzhou, China) following the manufacturer's protocol. Gene-specific primers were designed using NCBI Primer-BLAST (
https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi?LINK_LOC=BlastHome ) (Table 1). Quantitative real-time PCR (qRT-PCR) was conducted using the 2×Taq Pro Universal SYBR qPCR Master Mix Kit (Vazyme Biotech Co., Ltd., Nanjing, China) on a CFX96 Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Reactions were performed in 20 μL volumes containing 10 μL SYBR mix, 0.5 μM of each primer, and 50 ng cDNA template. Thermal cycling conditions were established following the methodology described by Dong et al.[36], with minor adaptations to optimize reaction specificity. The Actin gene[36] served as the internal reference for normalization. Each sample included three biological replicates, and three technical replicates.Table 1. List of primers used for qRT-PCR expression validation analysis.
Number Unigene ID Gene code Forward primer (5'→3') Reverse primer (5'→3') 1 Unigene0039492 MYB21 CCTCATCAGCCCCACCAAC CGTCATCTACACGACCAGCC 2 Unigene0025400 MYB308 CGCAGCTGGTTGAACCAAC GTGGAGAAGGGAGCTGGAG 3 Unigene0032323 MYB44 CTAACGAAGCCGCCATCTTC GGTGTGGAACTACATGTCGG 4 Unigene0029274 BHLH93 CCACCGTAGCCTGAACTCC GCTGACATCGACCATGAGC 5 Unigene0009307 MYB3R1 GTGGAGTTCTGCTGCTGTG CGGCAGTTCATTGCTGCTC 6 Unigene0025948 RAP2.9 CCGGTGCTATGTTCAGGTCAG ACACCGCCGTCTACTACCTC 7 Unigene0033886 PRR95 GACCGGCACTTCACTAGCT TAGCAGAGCAACGTCCTCG 8 Actin CATAATCCAGAGCCACATA TGCCAATCTACGAGGGTT -
The continuous floral differentiation of F. pallidiflora, from vegetative to reproductive meristems, can be divided into six main stages: the undifferentiated phase (S0), the bract primordium differentiation phase (S1), the floral primordium formation phase (S2), the petal primordium differentiation phase (S3), the stamen primordium differentiation phase (S4), and the pistil primordium differentiation phase (S5) (Fig. 1a). At S0, flower buds are very slim with a SAM of only about 2 mm. As the growth point initially broadens, the bract leaf primordium (S1), and flower primordium (S2) appear successively, followed by an obvious increase in the circumference of the buds, making the bracts clear and visible. Petals then differentiate (S3), followed by the differentiation of the stamen primordium (S4) and pistil primordium (S5) (Fig. 1a and b). Later, the development of the whole flower bud is completed, and differentiated sepals, petals, stamen, and pistils can be observed (Fig. 1a).
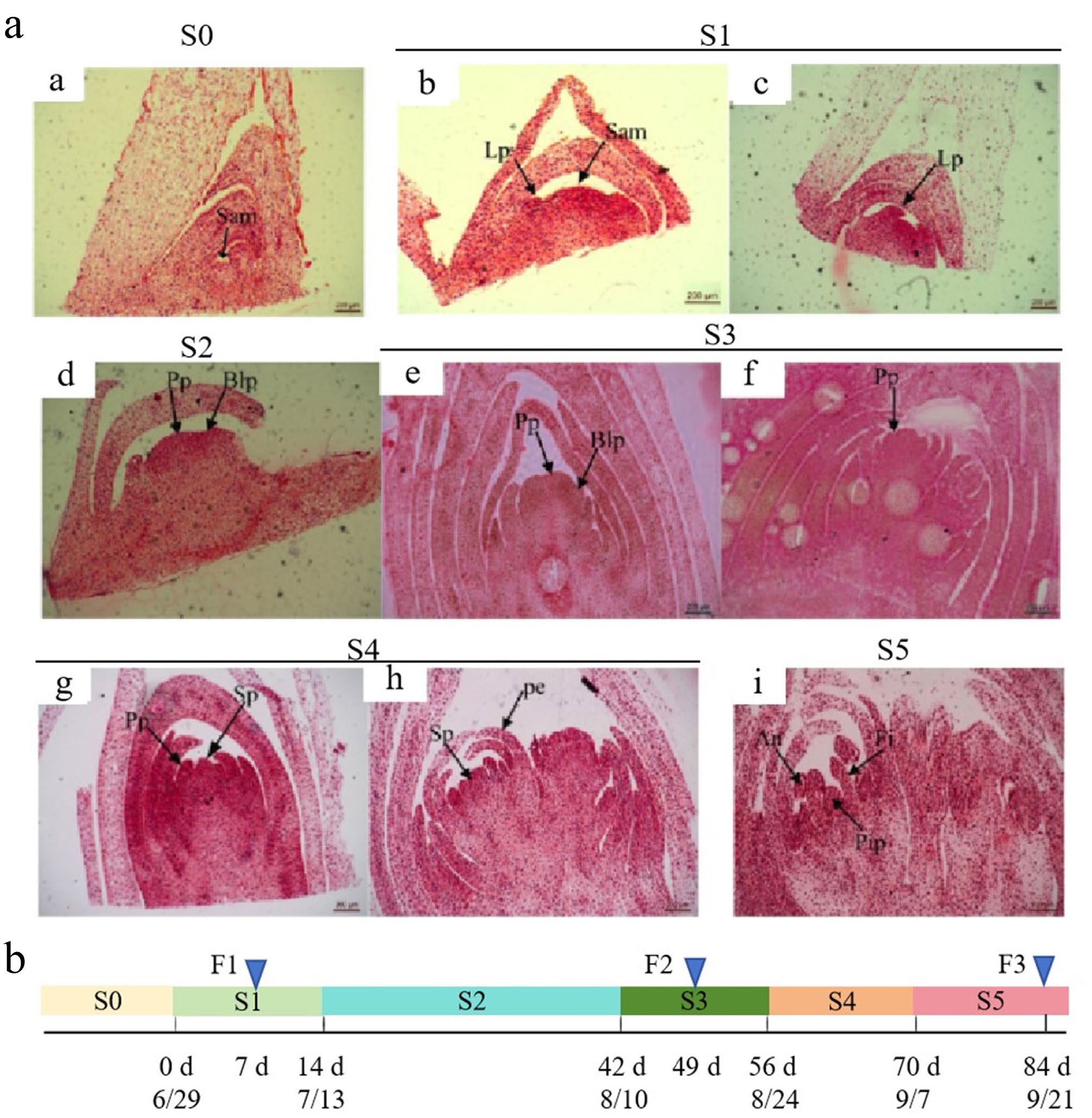
Figure 1.
Morphological characterization of floral bud differentiation in Fritillaria pallidiflora. a: Floral bud development was classified into six distinct stages (S0–S5) based on morphological characteristics. (a) Undifferentiated phase (S0). (b), (c) Bract primordium differentiation phase (S1). (d) Floral primordium formation phase (S2). (e), (f) Petal primordium differentiation phase (S3). (g), (h) Stamen primordium differentiation phase (S4). (i) Pistil primordium differentiation phase (S5). An: anther; Blp: bract leaf primordium; Fi: filament; Lp: leaf primordium; Pe: petal; Pip: pistil primordium; Pp: petal primordium; Sam: shoot apical meristem; Sp: stamen primordium. Scale bars: 200 μm. b: Sampling time line. F1, F2, and F3 represent three samples corresponding to the three representative developmental stages (S1, S2, and S3) of floral buds. F1 represents the sample from the S1 stage, sampled after 7 d of treatment (on July 6); F2 represents the sample from the S3 stage, sampled after 49 d of treatment (on August 17); F3 represents the sample from the S5 stage, sampled after 84 d of treatment (on September 21).
Sequencing, assembly, and annotation of the F. pallidiflora transcriptome
-
To identify key genes associated with floral bud development, RNA-seq sequencing was conducted on samples of F1, F2, and F3, corresponding to the three representative differentiation stages (S1, S3, and S5) of F. pallidiflora. Consequently, 405,088,264 clean reads were acquired, exhibiting a Q30 value greater than 90% and a GC content ranging from 50% to 52% (Table 2). These metrics suggest that the sequencing quality is high and suitable for assembly and further analysis. Ultimately, after splicing the clean reads with the Trinity program, a total of 74,353 high-quality unigenes were generated, with an average length of 782 bp and an N50 of 1305 bp. Out of these, 34,288 unigenes were functionally annotated, representing 46.12% of the total. Specifically, 33,839 (45.51%), 24,343 (32.74%), 21,176 (28.48%), and 13,957 (18.77%) unigenes were annotated in the Nr, Swiss-prot, KOG, and KEGG databases, respectively (Table 3). Meanwhile, reproducibility testing of samples was performed using PCA and clustering analysis based on the expression levels of all genes. As depicted in Fig. 2, the PCA plot (Fig. 2a), and expression clustering analysis plot (Fig. 2b) for each sample indicate that the transcriptome sequencing data was highly reproducible at each stage, allowing the different stages to be differentiated.
Table 2. Data quality statistics for RNA-seq of Fritillaria pallidiflora flower buds.
Samples Number of reads Q20 (%) Q30 (%) Clean reads ratio (%) GC content (%) F1-1 728,21,844 97.94% 94.83% 97.54% 51.30% F1-2 678,43,116 97.70% 94.27% 96.95% 50.86% F2-1 721,00,898 97.93% 94.77% 97.64% 51.42% F2-2 640,60,984 97.84% 94.55% 97.45% 51.89% F3-1 642,17,358 98.06% 95.04% 97.79% 50.63% F3-2 640,44,064 98.04% 95.02% 97.78% 50.42% Table 3. Statistics of gene annotation.
Database Number of annotated unigenes Annotation (%) Nr 33,839 45.51 Swissprot 24,343 32.74 KOG 21,176 28.48 KEGG 13,957 18.77 Total 34,288 46.12 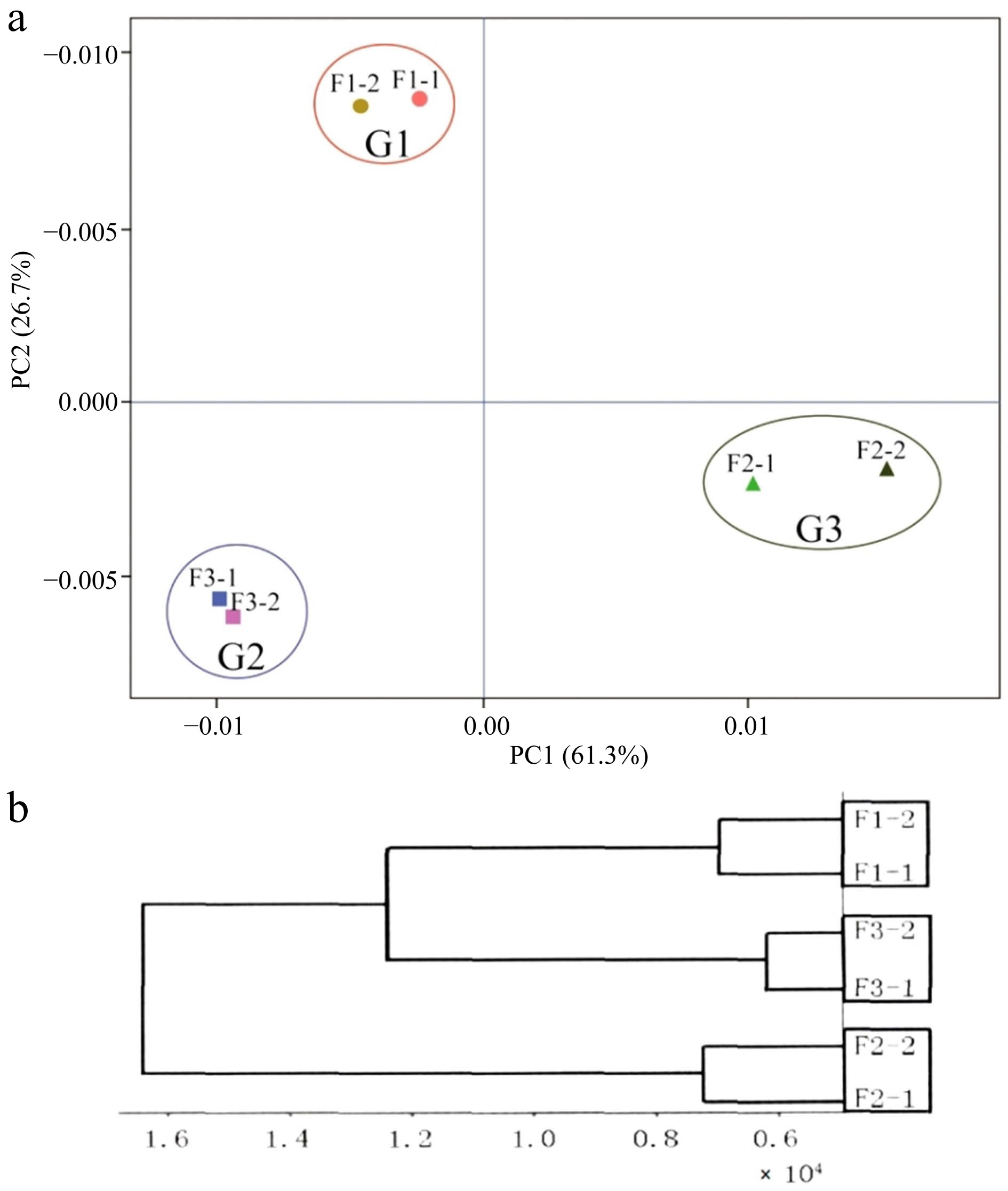
Figure 2.
(a) Principal component analysis (PCA) of each sample, and (b) cluster analysis of expression levels of samples. F1-1 and F1-2, F2-1 and F2-2, F3-1 and F3-2 represent two biological replicates for each sample respectively.
Analysis of differentially expressed genes
-
Clean reads were aligned to unigenes, and expression levels were estimated using the FPKM method. Three comparative groups, F1 vs F2, F2 vs F3, and F1 vs F3, were established to analyze the DEGs during floral bud development. In the F1 vs F2 group, 2,258 DEGs were identified, with 1,053 unigenes significantly up-regulated and 1,205 genes significantly down-regulated (Fig. 3a). In the F2 vs F3 comparison, there were 1,741 DEGs: 574 were up-regulated and 1,167 were down-regulated (Fig. 3a). In the F1 vs F3 comparison, there were 766 DEGs: 152 were up-regulated and 614 were down-regulated (Fig. 3a). As illustrated in a Venn diagram (Fig. 3b), a total of 3,402 DEGs were detected from three comparative groups, including nine common DEGs, and 1,079, 695, and 274 DEGs specifically expressed in F1 vs F2, F2 vs F3, and F1 vs F3, respectively.
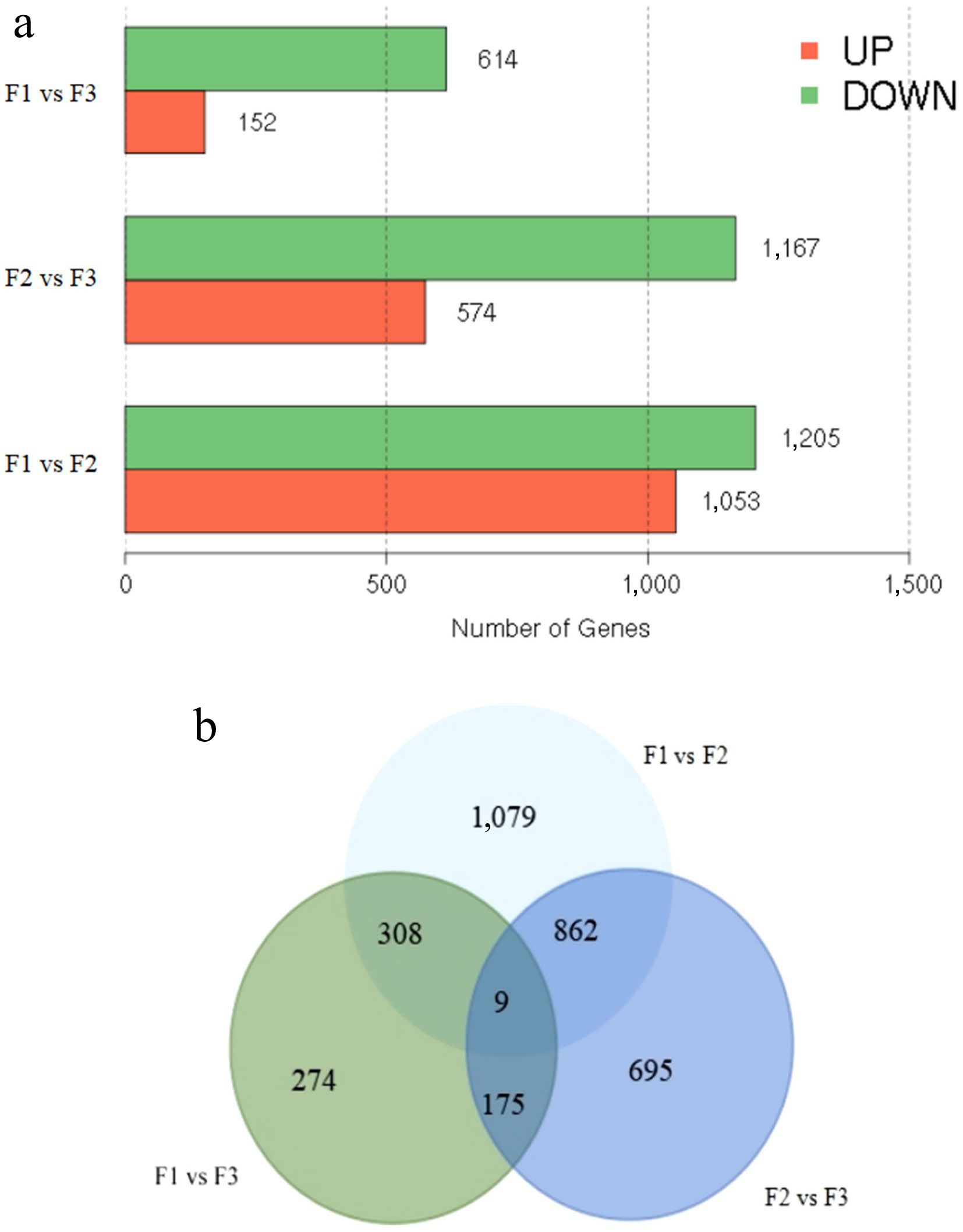
Figure 3.
Identification of differentially expressed genes (DEGs). (a) The number of up- and down-regulated DEGs between comparative groups in sequential stages. (b) Venn diagram of DEGs among groups.
To identify candidate DEGs involved in floral bud development in F. pallidiflora, we analyzed the K-Means clustering of gene expression patterns of these DEGs. As illustrated in Fig. 4, the DEGs were clustered into eight distinct expression patterns. DEGs in pattern 1 exhibited highly significant up-regulation at F2 compared to F1 and F3, whereas DEGs in pattern 2 showed highly significant down-regulation at F2 relative to F1 and F3 (p < 0.01). DEGs in pattern 3 showed significant down-regulation at F2 relative to F1 (p = 0.01 < 0.05). In other expression patterns, the differences between samples were not significant (p < 0.05). A total of 1,886 (1,041 + 845) DEGs demonstrated a clear differential expression pattern at F2. Consequently, our focus shifted to these 1,886 DEGs to uncover key genes (Supplementary Tables S1 and S2).
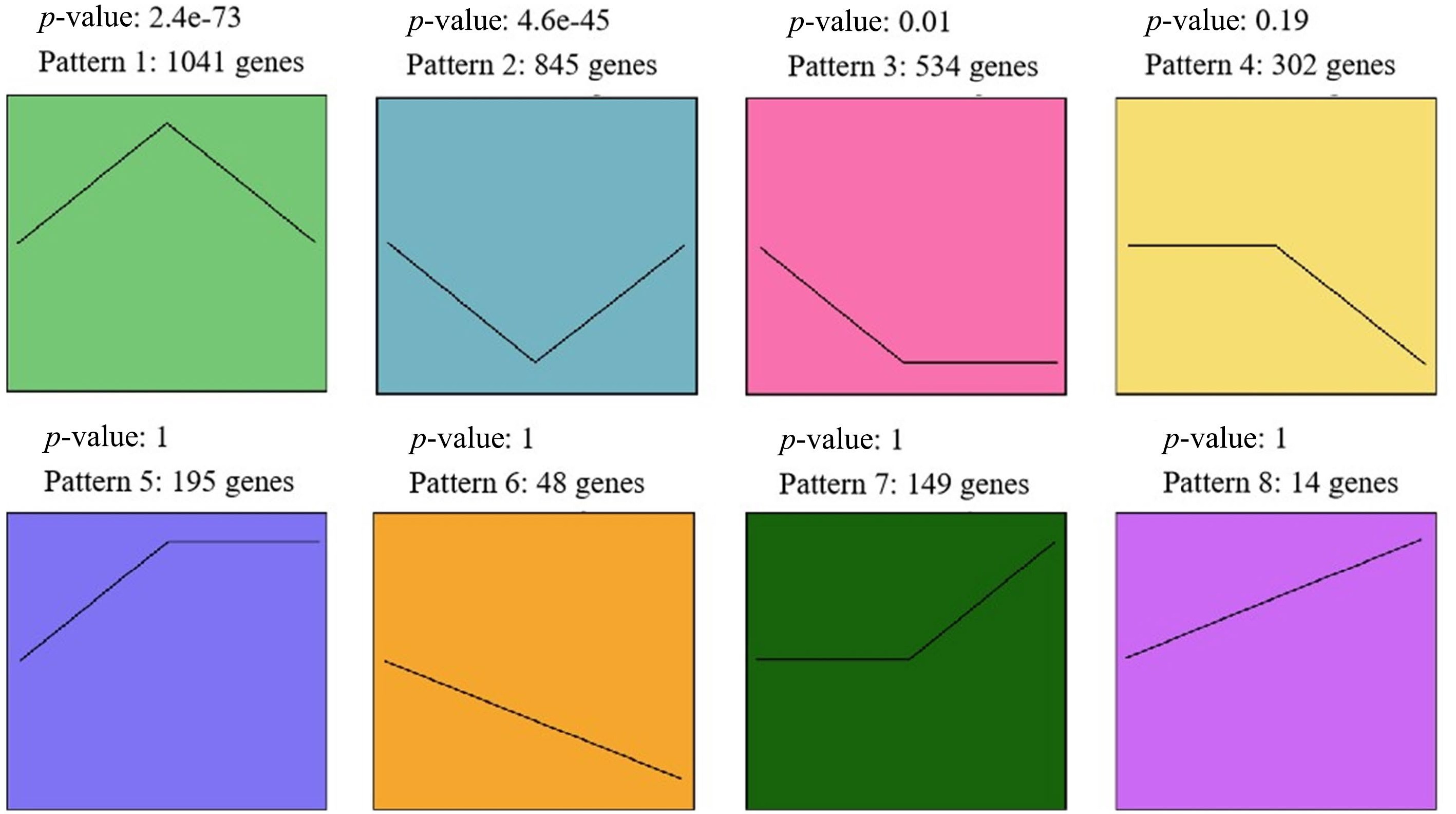
Figure 4.
Eight expression patterns across the three stages. Genes in pattern 1 were significantly up-regulated during F2 compared to F1 and F3; genes in pattern 2 were significantly down-regulated during F2 compared to F1 and F3.
Functional enrichment of differentially expressed genes
-
To investigate the functions of DEGs in expression patterns 1 and 2, GO and KEGG enrichment analyses were conducted. DEGs from expression patterns 1 and 2 were primarily clustered into GO terms associated with hormones, signals, secondary metabolism and regulation, including 'hormone-mediated signaling pathway' (GO:0009755), 'cellular response to hormone stimulus' (GO:0032870), 'signal transduction' (GO:0007165), 'signaling' (GO:0023052), 'metabolic process' (GO:0008152), 'cellular metabolic process' (GO:0044237), and 'regulation of transport' (GO:0051049) (Fig. 5a). KEGG analysis revealed that a significant number of DEGs were enriched in metabolism-related pathways, such as 'biosynthesis of secondary metabolites' (ko01110), 'carbon metabolism' (ko01200), and 'cysteine and methionine metabolism' (ko00270). Additionally, numerous DEGs were found to be enriched in signal transduction-related pathways, including 'plant hormone signal transduction' (ko04075), and 'MAPK signaling pathway-plant' (ko04016) (Fig. 5b).
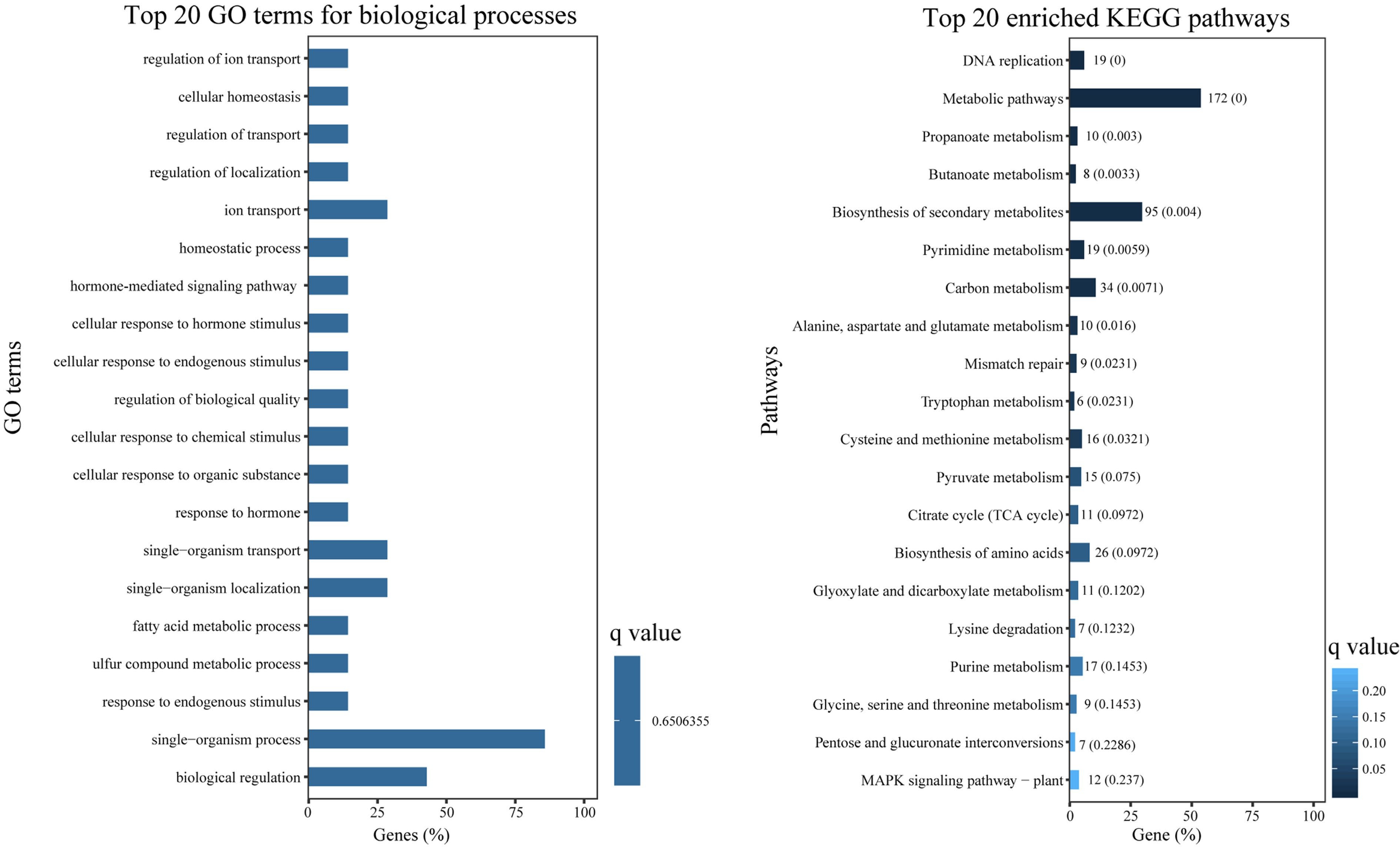
Figure 5.
GO enrichment and KEGG enrichment of DEGs. (a) The top 20 enriched GO terms in 'biological process' ontology. (b) The top 20 enriched KEGG pathways for DEGs.
Differentially expressed genes potentially related to different pathways involved in floral bud differentiation
-
In the present study, utilizing DEG enrichment with GO and KEGG analyses for patterns 1 and 2, a total of 44 unigenes potentially associated with floral bud differentiation were identified. These unigenes were categorized into five groups: circadian rhythm (seven genes), carbohydrate metabolism (eight genes), plant hormones (11 genes), temperature (seven genes), and other TFs (11 genes) (Fig. 6a).
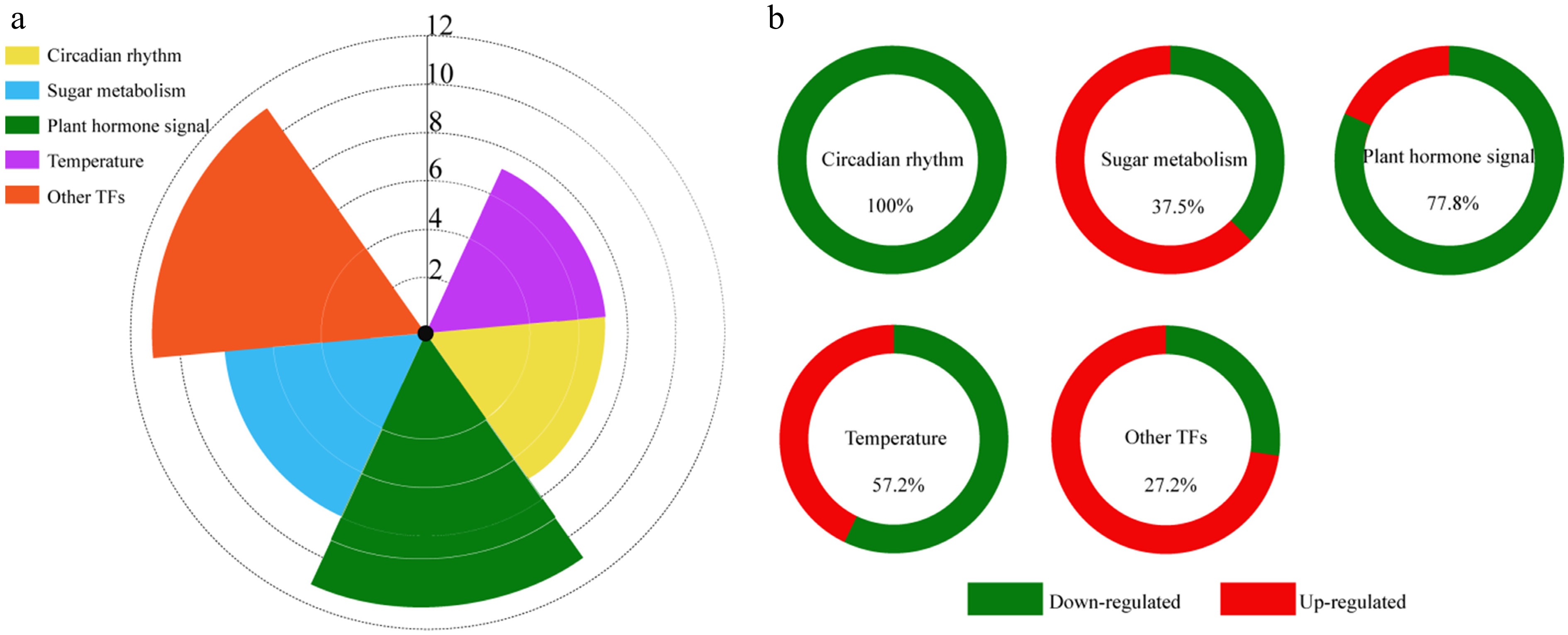
Figure 6.
DEGs related to floral bud differentiation. (a) Types and numbers of DEGs related to floral bud differentiation in Fritillaria pallidiflora. (b) Ratio of up- and down-regulated DEGs at F2 in each category.
There were seven DEGs in the circadian rhythm (photoperiod) pathway: CHS (Chalcone Synthase: unigene0003638), EFP (Early Flowering Protein: unigene0010964), APRR3 (Pseudo-response regulator 3: unigene0024917), PRR95 (Two-component response regulator-like PRR95: unigene0033886), LHY (Late Elongated Hypocotyl: unigene0040017), PHYC (Phytochrome C: unigene0051073), and FPA (Flowering Time Control Protein: unigene0046922). These seven genes were significantly down-regulated in F2 compared to F1 and F3 (p < 0.01) (Fig. 6b and Table 4).
Table 4. Candidate genes potentially related to floral bud differentiation of Fritillaria pallidiflora.
Pathways Unigene ID Gene name Annotation Expression pattern Expression trends Circadian rhythm Unigene0003638 CHS Chalcone synthase Pattern 2 F2 vs F1/F3 down Unigene0010964 EFP Early flowering protein Pattern 2 F2 vs F1/F3 down Unigene0024917 APRR3 Pseudo-response regulator Pattern 2 F2 vs F1/F3 down Unigene0033886 PRR95 Pseudo-response regulator Pattern 2 F2 vs F1/F3 down Unigene0040017 LHY Late elongated hypocotyl Pattern 2 F2 vs F1/F3 down Unigene0051073 PHYC Phytochrome C Pattern 2 F2 vs F1/F3 down Unigene0046922 FPA Flowering time control protein Pattern 2 F2 vs F1/F3 down Sugar Unigene0013557 SWEET14 Bidirectional sugar transporter Pattern 2 F2 vs F1/F3 down Unigene0045320 SP1 Starch phosphorylase Pattern 2 F2 vs F1/F3 down Unigene0048029 FBP Fructose-1,6-bisphosphatase Pattern 2 F2 vs F1/F3 down Unigene0014691 Susy1 SUS Pattern 1 F2 vs F1/F3 up Unigene0014693 Susy2 SUS Pattern 1 F2 vs F1/F3 up Unigene0014695 Susy3 SUS Pattern 1 F2 vs F1/F3 up Unigene0040753 Susy4 SUS Pattern 1 F2 vs F1/F3 up Unigene0033944 GBSSI Granule-bound starch synthase Pattern 1 F2 vs F1/F3 up Plant hormone signaling Unigene0021555 ABI5 ABA-insensitive Pattern 2 F2 vs F1/F3 down Unigene0022992 ERF1 Ethylene-responsive TF Pattern 2 F2 vs F1/F3 down Unigene0025948 RAP2.9 Ethylene-responsive TF Pattern 2 F2 vs F1/F3 down Unigene0041243 ABR1 Negative regulator of the ABA signaling pathway Pattern 2 F2 vs F1/F3 down Unigene0043547 GA3ox Gibberellin 2-β-dioxygenase Pattern 2 F2 vs F1/F3 down Unigene0015211 GASA1 Gibberellin-regulated protein Pattern 2 F2 vs F1/F3 down Unigene0025209 GASA2 Gibberellin-regulated protein Pattern 2 F2 vs F1/F3 down Unigene0025497 LOGL2 Cytokinin riboside 5'-monophosphate phosphoribohydrolase Pattern 1 F2 vs F1/F3 up Unigene0027601 CKX Cytokinin Dehydrogenase Pattern 1 F2 vs F1/F3 up Unigene0000376 ERF084 Ethylene-responsive TF Pattern 2 F2 vs F1/F3 down Unigene0073846 ARF Auxin response factor Pattern 2 F2 vs F1/F3 down Temperature Unigene0005515 HSF1 Heat stress TF Pattern 1 F2 vs F1/F3 up Unigene0036323 HSF2 Heat stress TF Pattern 2 F2 vs F1/F3 down Unigene0044090 HSP1 HSP Pattern 1 F2 vs F1/F3 up Unigene0050601 HSP2 HSP Pattern 1 F2 vs F1/F3 up Unigene0026481 HSP3 HSP Pattern 2 F2 vs F1/F3 down Unigene0033204 HSP4 HSP Pattern 2 F2 vs F1/F3 down Unigene0062240 HSP5 HSP Pattern 2 F2 vs F1/F3 down Other TFs Unigene0023294 WRKY71 WRKY 71 Pattern 2 F2 vs F1/F3 down Unigene0031385 MYB1R1 Myb 1R-1 Pattern 2 F2 vs F1/F3 down Unigene0039108 MYBX Myb-like protein X Pattern 2 F2 vs F1/F3 down Unigene0008589 BZIP bZIP-like protein Pattern 1 F2 vs F1/F3 up Unigene0009307 MYB3R1 Myb-related protein 3R-1 Pattern 1 F2 vs F1/F3 up Unigene0025400 MYB308 Myb-related protein 308 Pattern 1 F2 vs F1/F3 up Unigene0026858 NAC38 NAC domain-containing protein 38 Pattern 1 F2 vs F1/F3 up Unigene0029274 BHLH93 bHLH93 Pattern 1 F2 vs F1/F3 up Unigene0032323 MYB44 MYB44 Pattern 1 F2 vs F1/F3 up Unigene0039492 MYB21 Myb protein 21 Pattern 1 F2 vs F1/F3 up Unigene0068674 BHLH53 bHLH53 Pattern 1 F2 vs F1/F3 up ABA, abscisic acid; HSP, heat shock protein; SUS, Sucrose synthase; TF, transcription factor. The metabolism of soluble sugar might be closely related to floral bud development. Eight genes annotated in carbohydrate metabolism were identified from DEGs of patterns 1 and 2. Four SUSY genes (Sucrose Synthase: unigene0014691, unigene0014693, unigene0014695, and unigene0040753), and one GBSSI gene (Granule-Bound Starch Synthase: unigene0033944) were highly expressed in F2 compared to F1 and F3, while SP1 (Starch Phosphorylase: unigene0045320), SWEET14 (Bidirectional Sugar Transporter: unigene0013557), and FBP (Fructose-1,6-Bisphosphatase: unigene0048029) were significantly down-regulated at F2 (p < 0.01) (Fig. 6b and Table 4). The expression levels of starch and sugar synthase genes were higher in F2 than in F1 and F3, indicating that plant sugar metabolism was active, thus providing energy for floral bud development.
Plant endogenous hormones play important roles in the whole life cycle of plants, including the process of flowering[37]. In this study, a total of 11 genes associated with hormone signaling were identified among the DEGs of patterns 1 and 2. These genes are involved in the signal transduction pathways of abscisic acid (ABA), ethylene (ET), gibberellin (GA), cytokinin (CTK), and auxin (IAA) (Fig. 6a and Table 4). In the ABA signal transduction pathway, two DEGs were identified and annotated as ABI5 (Abscisic Acid-Insensitive 5: unigene0021555), and ABR1 (ABA Repressor 1: unigene0041243), both showing down-regulated expression in F2. Three DEGs involved in GA signaling, GA3ox (Gibberellin 2-β-Dioxygenase: unigene 0043547), and GASAs (gibberellin-regulated protein: unigene0015211, unigene0025209) were also down-regulated in F2 compared to F1 and F3. Additionally, three DEGs encoding ethylene-responsive TFs, ERF1 (unigene0022992), RAP2.9 (unigene0025948), and ERF084 (unigene0000376), and one DEG encoding an Auxin response factor (ARF: unigene0073846), were also identified. These genes were also down-regulated during F2. Furthermore, two DEGs in the cytokinin signal transduction pathway were identified, including LOGL2 (Cytokinin Riboside 5'-Monophosphate Phosphoribohydrolase: unigene0025497), and CKX (Cytokinin Dehydrogenase: unigene0027601). These two genes were up-regulated during F2.
Temperature can regulate the content of endogenous hormones and floral bud differentiation in plants[38]. In this study, seven DEGs related to temperature were identified, including two heat stress TFs (HSFs: unigene0005515, unigene0036323), and five heat shock proteins (HSPs: unigene0044090, unigene0050601, unigene0026481, unigene0033204, unigene0062240) (Fig. 6 and Table 4). As shown in Table 4, the two HSF genes exhibited opposite expression patterns. Among the five HSP genes, three were in pattern 2, while two were in pattern 1.
In this study, 11 TFs were identified, including MYBs, bHLHs, WRKYs, NAC, and others (Fig. 6 and Table 4). Six genes encoding MYB TFs were designated as MYB1R1 (unigene0031385), MYBX (unigene0039108), MYB21 (unigene0039492), MYB308 (unigene0025400), MYB3R1 (unigene0009307), and MYB44 (unigene0032323). Two genes, MYB1R1 and MYBX, were down-regulated, while the other genes were up-regulated during F2. One gene (unigene0023294) encoding a protein homologous to the WRKY71 TF of A. thaliana, was significantly down-regulated during F2. Additionally, four genes encoding TFs, NAC38 (unigene0026858), BHLH53 (unigene0068674), BHLH93 (unigene0029274) and bZIP (unigene0008589), were highly expressed in F2.
Validation of differentially expressed genes
-
To examine the reliability of transcriptomic data, seven DEGs were selected for qRT-PCR validation, each with three biological replicates (primer sequences are listed in Table 1). The expression patterns of these DEGs during three developmental stages of F. pallidiflora floral buds exhibited a high degree of concordance with the transcriptome profiling results (Fig. 7). This congruence confirms the accuracy and reproducibility of our transcriptomic sequencing data derived from distinct developmental stages of F. pallidiflora floral buds, thereby providing a reliable foundation for identifying key regulatory genes involved in floral bud development.
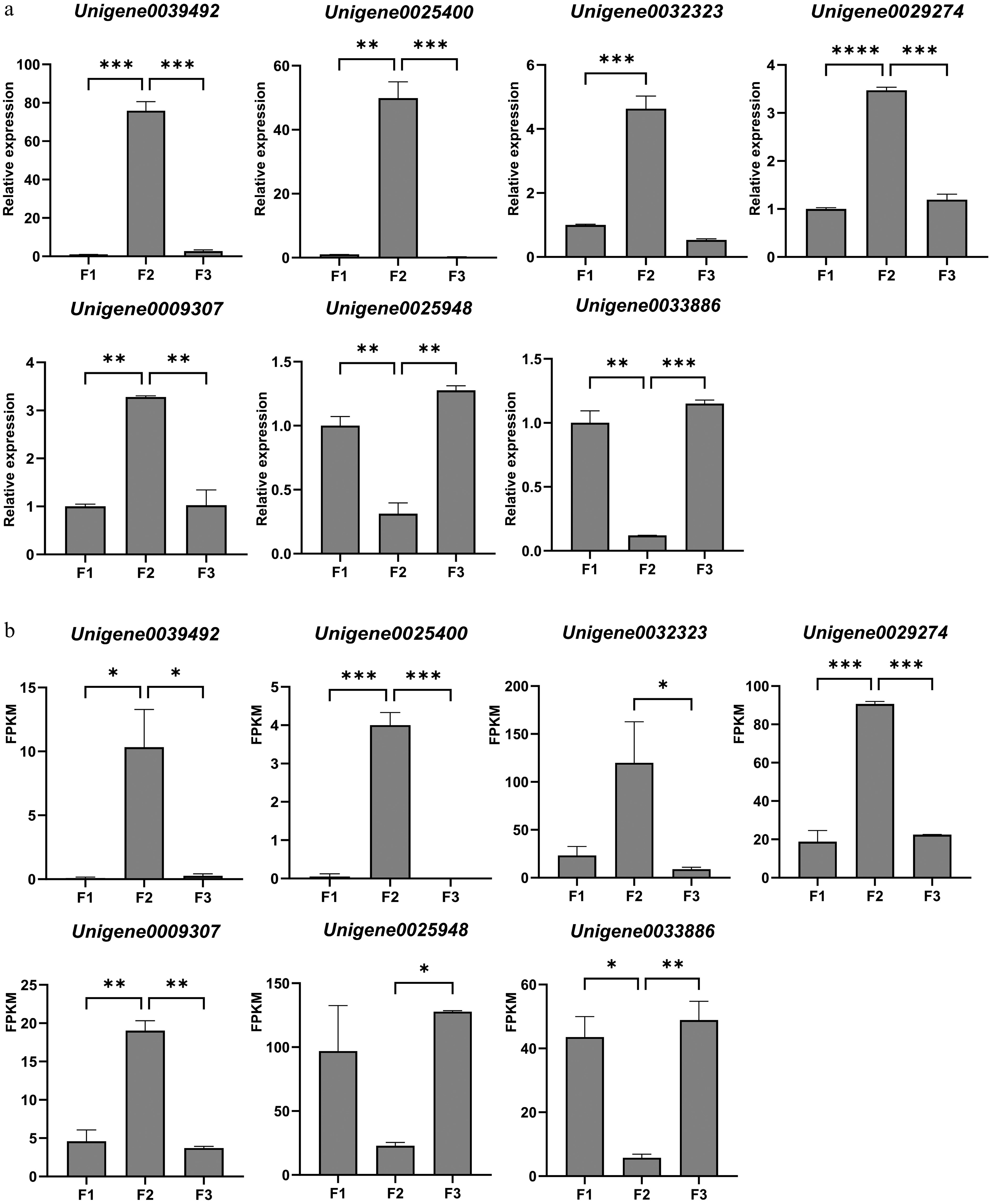
Figure 7.
Validation of the expression of selected DEGs from transcriptome data by qRT-PCR. (a) Relative expression results of qRT-PCR. (b) The FPKM value obtained from the transcriptome data. Values are means of three independent biological samples and each sample contains three plants. Error bars represent standard deviations and the statistical analysis was conducted using the Student's t-test. * denotes statistical significance, with the number of '*' corresponding to the level of significance (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001).
-
Flower bud development is crucial to the life cycle of ornamental plants[39]. It signifies a pivotal transition from vegetative to reproductive growth, and directly influences the quantity and quality of flowers produced by flowering plants[1,40,41]. In our study, the development of F. pallidiflora flower buds spanned nearly three months, and paraffin sections revealed that this process could be segmented into six distinct sub-phases (S0–S5): the undifferentiated phase (S0), bract leaf primordium differentiation (S1), flower primordium formation (S2), petal primordium differentiation (S3), stamen primordium differentiation (S4), and pistil primordium differentiation (S5). Prior to differentiation, flower buds gradually enlarged with only a small growth point, evolving into flower bud primordia. The bud primordia then began to elongate and widen, signaling the onset of floral bud differentiation. Subsequently, differentiated petals, stamens and pistils emerged in succession, culminating in the formation of a complete flower bud. The observed process was unique yet bore similarities to that of Magnolia sinostellata (Magnoliaceae)[42], Camellia azalea (Theaceae)[43], and Litsea cubeba (Lauraceae)[44].
Physiological and biochemical mechanisms underlying floral bud development
-
To investigate the complex molecular mechanisms underlying floral bud development in F. pallidiflora, RNA-seq analysis was conducted using samples from F1, F2, and F3, which correspond to flower buds at three differentiation stages: the bract primordium differentiation phase (S1), the petal primordium differentiation phase (S3), and the pistil primordium differentiation phase (S5). A total of 3402 DEGs were identified between sequential stages (F1 vs F2, F2 vs F3, and F1 vs F3), with 1041 DEGs significantly up-regulated at F2 compared to the other two stages (F1 and F3), and 845 DEGs significantly down-regulated at F2 (Figs 3 and 4). Based on the annotation of DEGs that were up- or down-regulated during the floral bud differentiation stage compared to the other two stages, 44 candidate unigenes were identified. These unigenes were found to be involved in circadian rhythm, carbohydrate metabolism, phytohormone signaling, temperature response, and various TFs. (Fig. 6 and Table 4), indicating their pivotal roles in initiating floral transition.
Flower bud development entails a series of physiological, biochemical, and morphological changes in the SAM, which occur following floral stimulation[45]. According to Liu et al., when a plant transitions from vegetative to reproductive growth, nutrients, regulatory substances, and metabolites in flower buds must reach a critical concentration to respond to the factors that induce flowering[46]. The controlled environment of in vitro conditions allows for such factors to be regulated[47]. Tu et al. observed that the metabolism of soluble sugar and soluble protein during lily (Lilium brownii var. viridulum) bud differentiation was positively correlated with the process of flower bud development[48]. Analysis of DEGs in the carbohydrate metabolism pathway during the flower bud development of F. pallidiflora, as indicated by sequencing results, revealed that most sucrose synthase genes were highly expressed during bud differentiation. The elevated expression of sucrose synthases at the bud differentiation stage suggests that sucrose content at this stage may be higher than that of the F1 and F3 stages, facilitating the complete development of Fritillaria flower buds.
Several studies have demonstrated that, in addition to nutrition, plant flower bud development is also regulated by various plant hormones, including GA, ABA, IAA, CTK, and others. Wang et al. observed that cytokinin, ABA and ethylene can promote flower bud development, while GA inhibits it[49]. GA3 accelerated flowering in the LA hybrid lily after the transcriptional regulatory network of the floral induction pathway was analyzed[50]. Wang et al. investigated the changes in endogenous hormone content during the development of Lycoris radiata flower buds and found that IAA and ABA played a significant role[51]. In their study, a total of nine genes associated with hormone signaling were identified from DEGs of patterns 1 and 2 involved in ABA, ET, GA, CTK, and IAA signal transduction. Specifically, two, three, three, and one genes were identified in the ABA, GA, ET, and IAA signal transduction pathways, respectively. All these genes were down-regulated during flower bud development. Furthermore, two DEGs in the CTK signal transduction pathway were also identified and were up-regulated during flower bud development. These findings suggest that the completion of flower bud development is a dynamic equilibrium process influenced by hormones. In addition, circadian rhythm and temperature are critical factors in floral bud development and the determination of flowering time. In many plants, a prolonged period of cold temperature is required for flowering, and varying photoperiods can also regulate flowering time. In the present study, DEGs of seven genes related to circadian rhythm (CHS, EFP, APRR3, PRR95, LHY, PHYC, FPA) and seven genes related to heat stress/shock were identified. This indicates that circadian rhythm and temperature may play significant roles in the development of flower buds in F. pallidiflora.
TFs regulate the expression of numerous downstream genes and are crucial for the growth, development, and metabolism of plants[52]. Hundreds of TFs have been isolated and identified from higher plants, including AP2/ERF, bHLH, bZIP, MADS-box, NAC, MYB, SBP-box, WRKY, etc.[22]. A large number of TFs were also identified in the development of F. pallidiflora flower buds. Based on annotation results, 11 TFs were identified, such as MYBs, bHLHs, WRKYs, and NACs. Other research has indicated that MYB TFs are particularly important in flower development. For instance, the MYB TF OsGAMYB in Oryza sativa is essential for floral organ development, and other GAMYB-like genes are involved in mediating GA signaling during growth and flowering responses[53]. TFs such as WRKY40/33/27/21/7, MYB, AP2/EREBP, bHLH, and NAC1/2/11 promote flowering of LA hybrid lily by responding to GA signals[50]. In our study, six MYB TFs were identified from DEGs, with four up-regulated and two down-regulated during F2. Previous studies have found that bHLH TFs are involved in various activities during flower development, including morphology[54]. bZIP proteins participate in numerous biological processes in plants, including flower development. Strathmann et al. observed a decrease in the expression of a bZIP gene, BZI-1, which affected petal development and resulted in smaller flowers[55]. In this study, only one bZIP gene was identified, exhibiting up-regulated expression during stage F2.
Our results provide a comprehensive analysis of gene expression patterns associated with three stages of floral bud development in F. pallidiflora. Numerous DEGs were identified. Gene annotation analysis enabled the classification of the main DEGs into five categories: circadian rhythm, carbohydrate metabolism, phytohormone signaling, temperature response, and other TFs. Genes implicated in various pathways suggest the coordinated regulation of F. pallidiflora floral development. This study constitutes an inaugural report on the dynamics of gene expression during floral bud differentiation in F. pallidiflora, offering a comprehensive gene repertoire for thorough investigation of its floral developmental mechanisms and the functional characterization of associated regulatory genes. However, this study is limited to a transcriptomic data analysis, and sole reliance on this approach has its limitations. While RNA-seq effectively captures dynamic gene expression patterns, it cannot confirm protein-level activity or post-translational modifications. Additionally, metabolic shifts in sugars, hormones, or secondary metabolites, which are key drivers of floral induction, need to be characterized. This study is limited to providing an analytical perspective at the transcriptional level, and future studies can integrate proteomics, targeted metabolomics, and verification of transgenic function. Subsequent metabolic analyses, including those of related hormones such as GA and ABA, would be necessary to validate the transcriptomic findings at specific stages. Comparative transcriptomic versus metabolic analyses have been employed to understand the regulatory mechanism of sipeimine biosynthesis in Fritillaria cirrhosa[56]. Nevertheless, these results provide an initial resource for further functional analyses of candidate genes that influence floral bud development in Fritillaria, and will be crucial for uncovering the molecular mechanisms mediating floral bud development in other perennial bulbous plants.
-
Not applicable.
This work was financially supported by the Introduced Talents Project of Hebei Agricultural University (ZD201729), and the National Natural Science Foundation of China (32071817).
-
The authors confirm their contributions to the paper as follows: experiment design and conceptualization: Hao L, Teixeira da Silva JA, Yu X; experiment execution: Hao L; transcriptomic data analysis and statistical analyses: Hao L; experimental supervision: Yu X; scientific coordination and advice, and general supervision: Teixeira da Silva JA. All authors reviewed the results and approved the final version of the manuscript.
-
The raw data of the analyses may be found at NCBI's BioProject ID PRJNA883043 (https://submit.ncbi.nlm.nih.gov/subs/bioproject/SUB12088758/overview). The Sequence Read Archive (SRA) submission will be released on 2025-10-24 or upon publication.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The list of 1,041 differentially expressed genes (DEGs) from pattern 1.
- Supplementary Table S2 T2 The list of 845 differentially expressed genes (DEGs) from pattern 2
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hao L, Teixeira da Silva JA, Yu X. 2025. Transcriptome analysis of floral bud development in Fritillaria pallidiflora. Ornamental Plant Research 5: e034 doi: 10.48130/opr-0025-0034 

Transcriptome analysis of floral bud development in Fritillaria pallidiflora
- Received: 26 January 2025
- Revised: 26 May 2025
- Accepted: 04 June 2025
- Published online: 29 August 2025
Abstract: Flowering represents a significant transition from vegetative to reproductive growth, with floral bud development being crucial for the life cycle of ornamental plants. Despite advances in model plants, the molecular regulatory mechanisms governing floral bud development in the perennial ornamental bulbous plant genus Fritillaria L remain unclear. In this study, we defined the exact developmental stages of the flower bud of Fritillaria pallidiflora and comprehensively analyzed gene expression patterns associated with floral bud development. Based on the morphological observations of paraffin sections, the process of floral bud differentiation in F. pallidiflora was classified into six distinct stages: the undifferentiated phase (S0), bract primordium differentiation phase (S1), floral primordium formation phase (S2), petal primordium differentiation phase (S3), stamen primordium differentiation phase (S4), and pistil primordium differentiation phase (S5). Transcriptomic analysis of samples (F1, F2, and F3) from three representative developmental stages (S1, S3, and S5), identified a total of 74,353 unigenes, of which 34,288 were functionally annotated as protein-coding genes. Across the three stages, a total of 3402 differentially expressed genes (DEGs) were identified, among which only nine genes exhibited differential expression consistently at all three stages. GO and KEGG pathway enrichment analyses of the 1886 DEGs revealed that 44 unigenes potentially related to floral bud differentiation were identified, including seven genes associated with circadian rhythm, eight genes involved in carbohydrate metabolism, 11 genes related to plant hormones, seven genes related to temperature, and 11 transcription factors, which can serve as candidate genes for studying floral bud development in Fritillaria. Collectively, these findings provide a valuable collection of gene expression patterns that are characteristic of floral bud differentiation in F. pallidiflora, which may lay a foundation for dissecting the molecular mechanisms underlying the regulation of flowering time and facilitating molecular breeding in Fritillaria species, contributing to elucidation of the molecular mechanisms that regulate floral bud development in other perennial bulbous plants.