










-
Regeneration refers to the capacity of plants to resume growth and reproduction through their own physiological mechanisms[1]. There are diverse forms of plant regeneration, such as de novo organogenesis, somatic embryogenesis, and tissue repair[2]. Plant regeneration not only serves as the foundation for genetic transformation but also represents an important field in modern plant biology and agricultural biotechnology[3]. Numerous key factors affect the transformation of cell fate during plant regeneration, including plant hormones, explants selection, environmental clues, and epigenetic modifications. Among these, cytokinin stands out as one of the most significant factors affecting plant regeneration.
Cytokinin, a well-known plant hormone, plays crucial roles in callus formation and tissue regeneration[4]. Treatment with cytokinin can induce cell proliferation and promote shoot formation from callus. The dynamic balance of endogenous cytokinin is essential for promoting plant regeneration[5]. Several cytokinin synthases genes have been identified for their important roles during shoot regeneration. Isopentenyl Transferases (IPTs) are the rate-limiting enzymes in cytokinin synthesis[6]. Overexpression of AtIPT4 can promote Arabidopsis callus shoot formation even in the absence of exogenous cytokinin[6]. Cytokinin oxidase/dehydrogenase (CKX) is the sole key enzyme for the irreversible degradation of cytokinin, and can regulate the homeostasis of cytokinin in plants[7]. CKXs play crucial roles in plant growth and development among different species. TaCKX2.1 and TaCKX2.2 are highly expressed in the spikes and stems of wheat, and their expression levels are positively correlated with the number of grains per spike[8]. In rice, OsCKX11 was important for delaying leaf senescence, increasing the grain number, and coordinately regulating source and sink relationship[9]. Compared with WT, the osckx11 mutant presented with significantly increased branch, tiller, and grain number[9]. Previous research indicated that modulation of the level of endogenous cytokinin via CKXs inhibition can positively affect plant growth, development, and yield[10]. Inhibiting the activity of CKXs using phenyl-adenine could enhance the accumulation of endogenous cytokinin and promote shoot regeneration[11]. However, the function of CKXs in regulating plant regeneration has been rarely studied, and the molecular mechanism by which plants regulate the expression of CKXs to affect cytokinin homeostasis, and subsequently influence plant regeneration, remains unclear.
Wheat is the most widely distributed and cultivated food crop in the world, rich in various trace elements, dietary fiber, vitamins, and fat. In China, wheat is the main food crop, with both its production and consumption ranking first in the world[12]. To obtain high yield wheat varieties, wheat breeding methods have gradually shifted from conventional cross breeding to molecular design breeding using modern biotechnology[12]. However, the weak regeneration ability of wheat has led to difficulties in genetic transformation, which significantly limits the progress of wheat molecular design breeding. Therefore, screening for key factors that can significantly enhance the regeneration ability of wheat is crucial for the development of the wheat industry. In recent years, significant progress has been made in wheat transformation technology. Overexpression of the GRF4 (GROWTH-REGULATING FACTOR4) and GIF1 (GRF-INTERACTING FACTOR1) has notably improved the transformation efficiency and regeneration ability of wheat[13]. Overexpression of the growth-regulating gene TaWOX5 has also promoted the efficient transformation of wheat, achieving genetic transformation and improvement of multiple wheat varieties[14]. Liu et al. demonstrated that the DNA binding with One Finger (DOF) transcription factors TaDOF3.4 and TaDOF5.6 can significantly improve the transformation efficiency and regeneration frequency of wheat[15]. Yu et al. showed that TaLAX1 can promote the shoot regeneration of wheat by enhancing the expression of TaGIF1, TaIPT1, and auxin-responsive genes, thereby improving the genetic transformation and genome editing efficiency of wheat[16]. Thus, the application of regeneration promoting factors can effectively improve the efficiency of wheat regeneration and genetic transformation, and the identification of new, highly efficient wheat regeneration promoting factors is of great importance for the development of the wheat industry.
DOF transcription factors belong to a plant-specific transcription factor family, typically consisting of 200 to 400 amino acids[17]. The DOF transcription factors contain a highly conserved DOF domain at the N-terminus, which is composed of 52 amino acids with a highly conserved C2C2-type single zinc finger DNA binding motif[17]. The DOF transcription factors specifically bind to the 5'-(T/A)/AAAG-3' element in the promoter of target genes through the DOF domain[17]. Since the first DOF transcription factor was identified in maize, genes encoding proteins containing the DOF domain have been identified in many plant species, including Arabidopsis, rice, and wheat[18−20]. The DOF transcription factors not only participate in regulating plant responses to various biotic or abiotic stresses but also play important roles in plant regeneration. In Arabidopsis, AtOBP4 (a DOF transcription factor) negatively regulated cell proliferation and expansion via inhibiting the expression of CyclinB1;1, CDKB1;1, and XTH genes[21]. Four DOF transcription factors (HCA2, TMO6, DOF2.1, and DOF6) were highly expressed at the injured site of the hypocotyl of Arabidopsis, promoting wound healing and tissue regeneration[22]. In wheat, although it has been reported that overexpression of TaDOF5.6 and TaDOF3.4 can significantly improve the genetic transformation efficiency and shoot regeneration ability of wheat varieties such as Fielder, JM22, and KN199[15], the underlying mechanism of how the DOF transcription factors regulate regeneration remains unclear.
In this study, through the comprehensive analysis of the multi-omics data during the early stage of wheat callus induction in previous studies, TaDOF4.7-B, a gene with a high expression level at the early stage of wheat callus induction, was identified. This suggested that TaDOF4.7-B might serve as a crucial transcription factor in regulating wheat regeneration. Experimental results from genetic, biochemical, and transgenic assays revealed that TaDOF4.7-B modulated the cytokinin homeostasis in wheat by specifically binding to the promoters of multiple CKX genes and inhibiting their expression, thereby leading to significant enhancement of the wheat regeneration efficiency. These findings not only pinpoint a novel regeneration associated factor but also clarify a mechanism by which DOF transcription factors regulate cytokinin homeostasis.
-
The experimental material was hexaploid wheat (Triticum aestivum L.) cultivars: Fielder. Fielder seeds were generously provided by Japan Tobacco Inc. and was employed for genetic transformation experiments. Wheat was grown under controlled environmental conditions in growth chambers with a photoperiod of 16/8 h (light/dark) at 22 °C following seed germination. Phenotypic evaluation of agronomic traits was conducted during the 2022-2023 growing season at the experimental field of Shandong Agricultural University (Shankou County, Taian City, Shandong Province, China).
Plasmid construction
-
The Ubi::TaDOF4.7-B-6MYC construct was generated using the PC186 vector by the Gateway system (Invitrogen, Carlsbad, CA, USA). The sequence of 6MYC and several restriction enzyme cutting sites (GCGGCCGCGGATCCGAATTCGGTACCCATATGCTGCAGGTCGACACTAGTGAACAGAAACTGATCTCTGAAGAAGATCTGGAACAGAAACTGATCTCTGAAGAAGATCTGGAACAGAAACTGATCTCTGAAGAAGATCTGGAACAGAAACTGATCTCTGAAGAAGATCTGGAACAGAAACTGATCTCTGAAGAAGATCTGGAACAGAAACTGATCTCTGAAGAAGATCTGGGCGCGCC) was synthesized and inserted into the pENTR vector to generate the mcs-6MYC-ENTR vector. The TaDOF4.7-B CDS sequences were amplified using Phanta Max Master Mix (Vazyme, Nanjing, China; P515-01) and inserted into the mcs-6MYC-ENTR vector between NotI and BamHI. TaDOF4.7-6MYC-ENTR was then recombined into the PC186 vector through Gateway LR recombination.
Wheat transformation
-
Immature wheat (cv. Fielder) embryos were harvested at 14 d post-anthesis and subjected to Agrobacterium-mediated transformation using the EHA105 strain harboring the designated transformation vectors, following established protocols[23]. Post-inoculation, embryos were initially cultured on WLS-AS medium for 48 h. Following embryonic axis excision, the explants underwent sequential culture on selective media: WLS-Res medium for 5 d, followed by WLS-P5 medium (contain 0.5 mg/l 2,4-D and 5mg/L phosphinothricin but excluding cytokinin) for 14 d. Subsequently, each explant was bisected and transferred to WLS-P10 medium supplemented with phosphinothricin (10 mg/L) for 21 d to facilitate callus induction. Proliferated callus tissue was then transferred to LSZ-P5 medium for shoot regeneration. Upon shoot development, plantlets were transferred to LSF-P5 medium for root development and elongation. The abbreviations CIM used in this study indicates the WLS-AS, WLS-Res, WLS-P5, and WLS-P10 medium, and SIM indicates the LSZ-P5 medium.
Regenerative phenotype calculations
-
The regeneration frequency and regenerating shoot frequency were calculated after the immature embryos had been cultured on CIM for 42 d, and then on SIM for 20 d. For the calculation of regenerated shoots, only those > 2 cm in length were considered, as they exhibited a higher potential for development into fully grown seedlings and plants compared with smaller shoots. Regeneration frequency was the number of calli that showed at least one regenerating shoot relative to the total number of inoculated embryos. Regenerating shoot frequency was calculated as the number of regenerating shoots per total number of inoculated embryos.
Total RNA isolation and real-time qPCR analysis
-
Total RNA was extracted from the indicated tissues using the CW Bio Ultra pure RNA Kit. The FastKing RT Kit (Tiangen Biotech, Beijing, China) was used to synthesize first-strand cDNA. SYBR Green Real-Time PCR Master Mix (Tiangen Biotech) was used to perform RT-qPCR analysis. The SYBR Green Master Mix was used to dilute each cDNA. Relative transcript levels were normalized to those of TaTublin. All measurements were carried out with three biological replicates. The comparative CT method was used to determine expression values. The primers used for RT-qPCR analysis are listed in Supplementary Table S1.
Transcriptome analysis
-
For transcriptome sequencing, calli induced from de novo transformed immature embryos transformed with the empty vector or Ubi::TaDOF4.7-B-6MYC vector were collected and cultured on CIM for 3, 5, 8, 11, and 14 d. Transcriptomics analysis was performed using previously described protocols[24]. Three independent biological replicates were performed for each specimen; sequencing was performed by OE Biotech (Shanghai, China). Raw reads were processed using FASTP (v0.23.1) to remove adapters, trim low-quality bases, and filter bad reads[25]. Clean reads were aligned to the wheat reference genome (IWGSC RefSeq v1.0)[26] using HISAT2 (v2.0.5)[27], and the aligned reads were sorted and converted into bam files using samtools (v1.5)[28]. FeatureCounts (v2.0.1) was used to count the number of paired mapping reads that overlapped each annotated gene (IWGSC Annotation v1.1)[29]. The count matrices were used to identify DEGs using the R package DESeq2 (v1.28.1), with thresholds of Padj < 0.05 and abs (log2 FoldChange) > 1[30]. K-means cluster analysis of DEGs was performed in R (v4.3.2) using Z-scaled TPM, and the gene expression heat map was displayed using the R package ComplexHeatmap (v2.4.3), and TBtools-II[31,32].
RNA in situ hybridization
-
TaDOF4.7-B CDS was amplified and ligated into the pEasy-Blunt3 vector, and the insertion direction of the fragments was verified by sequencing. The clones with forward or reverse sequences were screened by DNA sequencing; then the sequences were labeled as sense or antisense RNA probes in vitro using T7 RNA polymerases. RNA in situ hybridization was performed with digoxigenin-labeled sense and antisense probes on 8-mm sections of the indicated tissues as described previously[33].
Chromatin immunoprecipitation (ChIP) analysis
-
T2-generation transgenic lines overexpressing TaDOF4.7-B (15 d old) were harvested for ChIP analysis according to previously described protocols with minor modifications[34]. The DNA fragments combined with total protein were co-immunoprecipitated using anti-MYC antibody (1:500, Sigma-Aldrich, C3956-100UG). The IgG antibody (1:500, MIllipore, 12-371B) were used as negative controls. Plantpan4.0 (
https://plantpan.itps.ncku.edu.tw/plantpan4/promoter_analysis.php ) was used to analyze the potential binding sites of TaDOF4.7-B in the promoter of target genes. Primers were designed based on the predicted binding site positions. For each ChIP assay, we performed three biological repeats. All primers used for the ChIP assays are listed in Supplementary Table S1.Luciferase activation assays
-
The promoters with 5'UTR of the detected genes were individually fused with the luciferase reporter gene via the XhoI and BamHI sites of the pGreenII 0800-LUC vector[35]. The coding region of the TaDOF4.7-B gene was fused to the pGreenII 62-SK vector downstream of the 35S promoter between the BamHI and XhoI sites. The plasmids were transformed into A. tumefaciens strain EHA105. Nicotiana benthamiana leaves were infiltrated with different mixed combinations of Agrobacterium, and collected for transient transcription assays as described previously[33].
Cytokinin content measurement
-
Calli induced from immature embryos that had been transformed with the empty vector or Ubi::TaDOF4.7-B-6MYC vector were collected, and cultured on CIM for 42 d. Endogenous cytokinin levels were measured by Wuhan Greensword Creation Technology using LC-MS/MS analysis as reported previously[36].
Field trait evaluation
-
Grain morphometric and yield component analysis were conducted using wheat seeds randomly sampled from field trials (12 seeds per 1.2-m row). Hundred-grain weight, grain length, and grain width were quantified. Additional agronomic parameters, including spike length, grain number per spike, spikelet number per spike, and effective tiller number, were assessed post-harvest under field conditions.
Statistical analysis
-
At least 10 spikes or plants were measured for each trait. Statistical analyses were performed with GraphPad Prism 8. All data were analyzed using Student's t-test (two-tailed), or one-way ANOVA with Tukey's multiple comparisons test, and p < 0.05 was considered significant.
-
By analyzing the chromatin accessibility of the promoter of DOF transcription factors in wheat during the early stage of callus induction using the published data, the chromatin accessibility of the promoters of TaDOF4.7-B/D was found to be higher than TaDOF4.7-A (Supplementary Fig. S1)[15]. RT-qPCR was used to detect the expression levels of TaDOF4.7-A/B/D throughout the wheat regeneration process. The results showed that the expression level of TaDOF4.7-B was higher than that of TaDOF4.7-A/D during the regeneration process (Fig. 1a). Thus, TaDOF4.7-B was selected for further study. To clarify the subcellular localization of TaDOF4.7-B, TaDOF4.7-B was fused to green fluorescent protein (GFP) in the overexpression vector 35S::TaDOF4.7-B-GFP. The fusion protein was transiently expressed in N. benthamiana leaves using Agrobacterium infiltration. And the green fluorescent was observed in the nucleus, indicating that TaDOF4.7-B may function in the nucleus (Fig. 1b). RNA in-situ hybridization experiments were used to observe the expression pattern of TaDOF4.7-B, using immature embryos of Fielder wheat and calli at different stages of the regeneration process as materials. TaDOF4.7-B transcripts accumulated in the compact cell clusters of wheat calli, which were the regions where shoot primordia might emerge (Fig. 1c). These results implied that TaDOF4.7-B might play important roles in the wheat regeneration process.
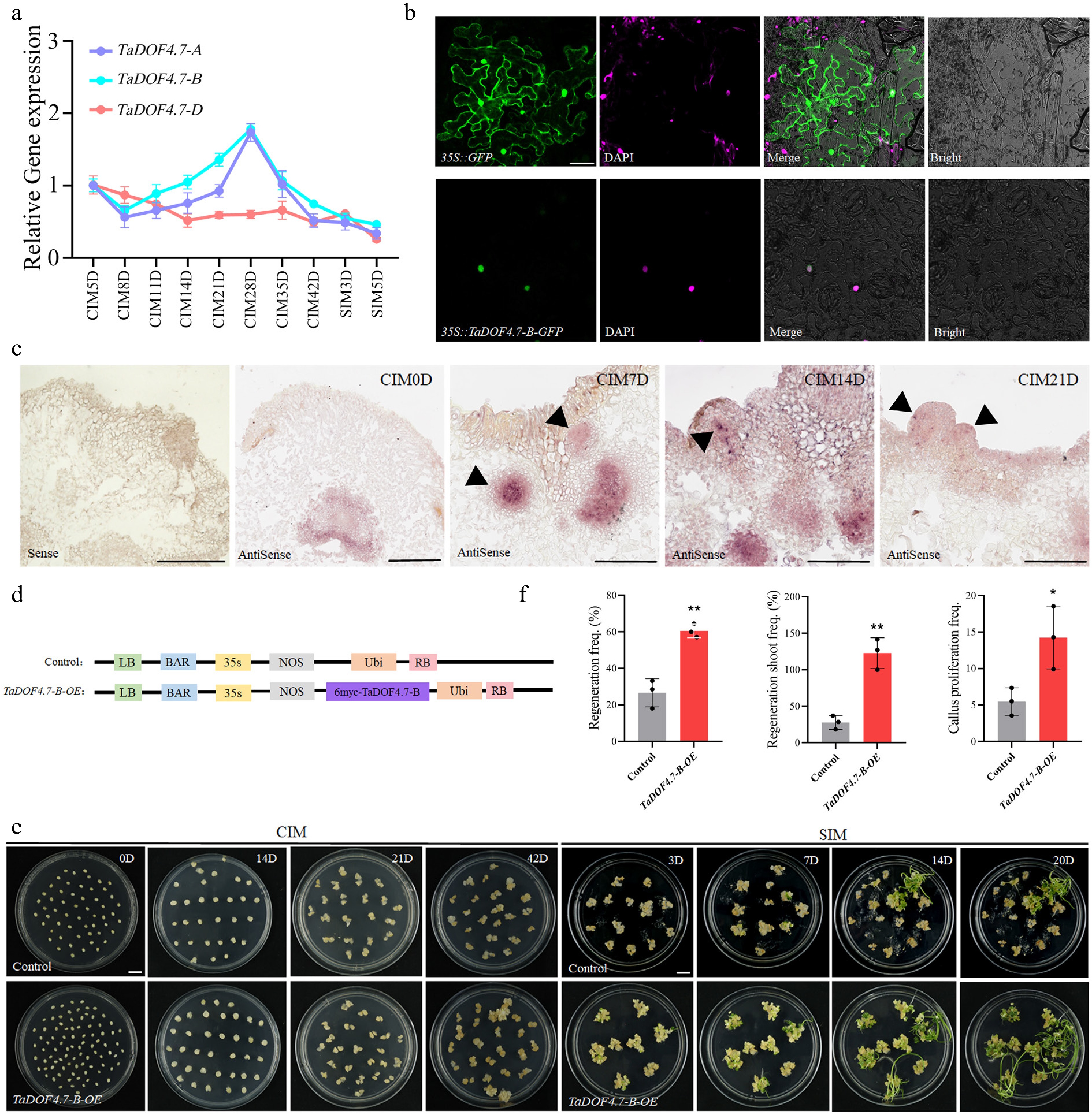
Figure 1.
TaDOF4.7-B enhances shoot regeneration in wheat. (a) The expression level of TaDOF4.7-A/B/D throughout the wheat regeneration process. The error bars indicate the means ± SEs of three independent biological replicates (n = 3). (b) Transient expression of the 35S::TaDOF4.7-B-GFP constructs in N. benthamiana leaves using Agrobacterium infiltration. Green fluorescence was observed using a confocal microscope, and the nucleus were visualized by DAPI staining. 35S::GFP was used as a control. (c) RNA in situ hybridization experiments. Immature embryos of Fielder wheat and calli at different stages of the regeneration process were as materials. (d) Schematic representation of the Ubi::TaDOF4.7-B-6MYC vector. The PC186 empty vector was used as a control. (e) Shoot regeneration phenotypes of immature embryos transformed with the empty vector (control) or Ubi::TaDOF4.7-B-6MYC vector (TaDOF4.7-B-OE). CIM, callus induction medium; SIM, shoot induction medium. Scale bar, 1 cm. (f) Regeneration frequencies, regenerating shoot frequency, and callus proliferation frequencies of immature embryos transformed with control or Ubi::TaDOF4.7-B-6MYC vector. Regeneration frequency = number of calli showing at least one regenerating shoot/number of inoculated embryos × 100%. Regenerating shoot frequency = number of regenerating shoots/numbers of inoculated embryos × 100%. Callus proliferation frequency = increased weight of callus after induction on CIM for 42 d/weight of immature embryos before induction. Values in (f) are means ± SE from three independent experiments (n = 3). Asterisks indicate significant differences (* p < 0.05, ** p < 0.01) based on 2-tailed Student's t-tests.
To explore the role of TaDOF4.7-B in the shoot regeneration process of wheat, an overexpression vector Ubi::TaDOF4.7-B-6MYC was constructed using the PC186 vector (Fig. 1d). The PC186 empty vector was used as a negative control. These vectors were transformed separately into immature embryos of Fielder through the PureWheat Agrobacterium-mediated wheat genetic transformation technology[23]. The results showed that the calli transformed with the Ubi::TaDOF4.7-B-6MYC vector (TaDOF4.7-B-OE) exhibited earlier appearance of shoot primordia, a better growth trend of calli, and a larger number of green shoot primordia (Fig. 1e). The regeneration frequency, regenerating shoot frequency, and callus proliferation frequency were statistically analyzed. The regeneration efficiency of the TaDOF4.7-B-OE transgenic calli was significantly higher than that of the control. The regeneration frequency increased from 22.65% ± 1.44% to 60.57% ± 3.43%, the regenerating shoot frequency increased from 32.11% ± 3.99% to 122.91% ± 18.75%, and the callus proliferation frequency increased from 6.28 ± 1.65 to 13.01 ± 1.28 (Fig. 1f). These results demonstrated that TaDOF4.7-B can improve the shoot regeneration ability of wheat.
Transcriptome analysis of the mechanism by which TaDOF4.7-B promotes shoot regeneration in wheat
-
To explore the mechanism by which TaDOF4.7-B promotes shoot regeneration in wheat, callus materials were collected at 3 (CIM 3 d) (D3), 5 (CIM 5 d) (D5), 8 (CIM 8 d) (D8), 11 (CIM 11 d) (D11), and 14 d (CIM 14 d) (D14) after transforming immature embryos of Fielder wheat with the Ubi::TaDOF4.7-B-6MYC vector (TaDOF4.7-B-OE) for RNA-sequencing (Supplementary Fig. S2). Transformations with the PC186 empty vector served as a control [CIM 3 d (P3), CIM 5 d (P5), CIM 8 d (P8), CIM 11 d (P11), CIM 14 d (P14)]. Compared with transformation using the PC186 empty vector, the RNA-seq data revealed 1,610 differentially expressed genes (DEGs) (1,401 genes were upregulated and 209 genes were downregulated) at CIM 3 d (Fig. 2a, Supplementary Table S2), 1,374 DEGs (1,194 genes were upregulated and 180 genes were downregulated) at CIM 5 d (Fig. 2b, Supplementary Table S3), 5,631 DEGs (4,336 genes were upregulated and 1,295 genes were downregulated) at CIM 8d (Fig. 2c, Supplementary Table S4), 7,295 DEGs (3,197 genes were upregulated and 4,098 genes were downregulated) at CIM 11d (Fig. 2d, Supplementary Table S5), 14,056 DEGs (8,356 genes were upregulated and 5,700 genes were downregulated) at CIM 14 d in the calli transformed with Ubi::TaDOF4.7-B-6MYC (Fig. 2e, Supplementary Table S6). The data revealed that the number of DEGs significantly increased after transforming immature embryos of Fielder wheat with the Ubi::TaDOF4.7-B-6MYC vector for 8 d.
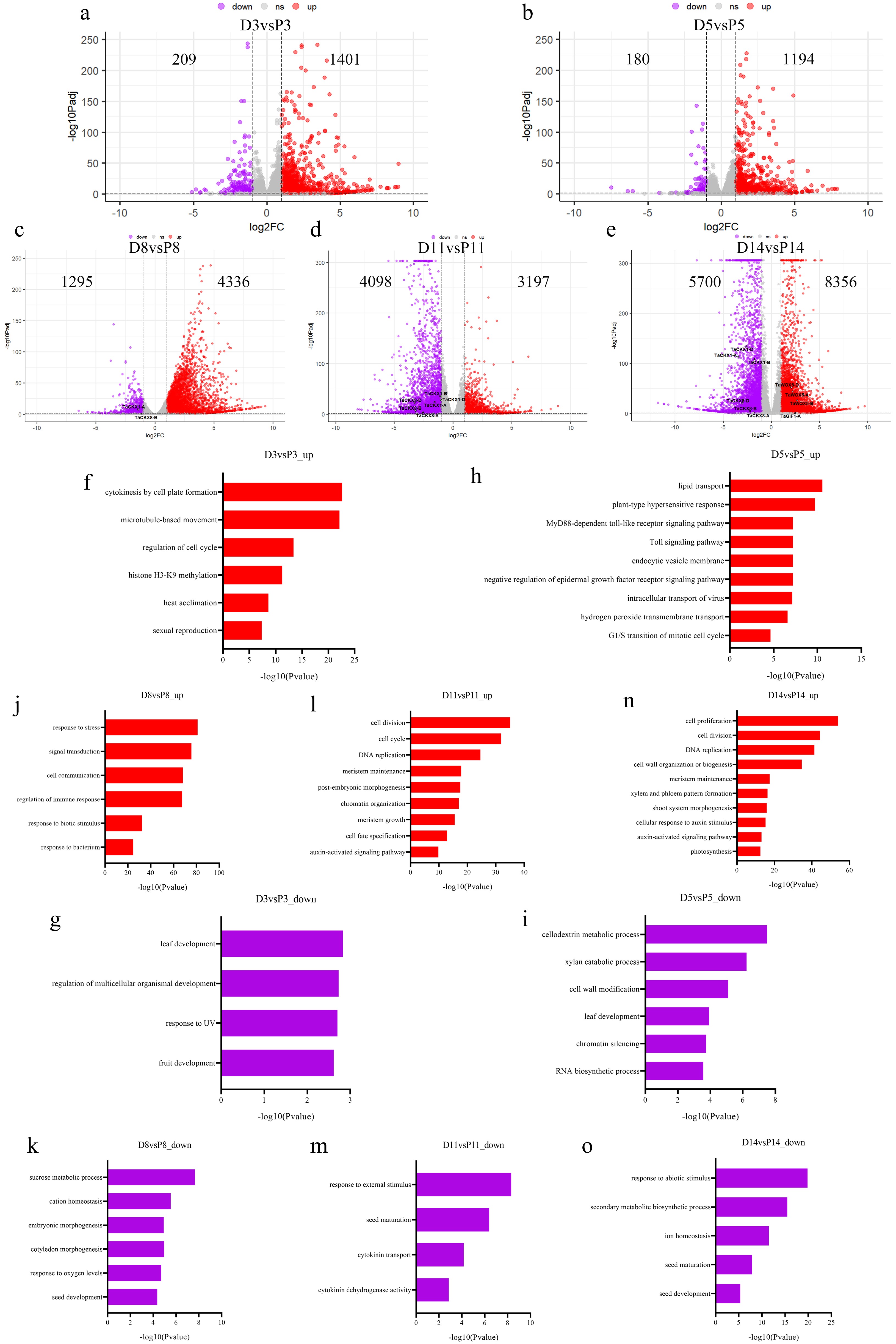
Figure 2.
RNA-seq analysis of immature embryos of Fielder wheat after transforming with the Ubi::TaDOF4.7-B-6MYC vector or PC186 empty vector. (a) Volcano plot of up- and downregulated genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 3 d (D3 vs P3). (b) Volcano plot of up- and down-regulated genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 5 d (D5 vs P5). (c) Volcano plot of up- and down-regulated genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 8 d (D8 vs P8). (d) Volcano plot of up- and down-regulated genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 11 d (D11 vs P11). (e) Volcano plot of up- and down-regulated genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 14 d (D14 vs P14). (f), (g) GO enrichment analysis of up- (F, D3 vs P3_up) and down-regulated (G, D3 vs P3_down) genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 3 d. (h), (i) GO enrichment analysis of up- (H, D5 vs P5_up) and down-regulated (I, D5 vs P5_down) genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 5 d. (j), (k) GO enrichment analysis of up- (J, D8 vs P8_up) and down-regulated (K, D8 vs P8_down) genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 8 d. (l), (m) GO enrichment analysis of up- (L, D11 vs P11_up) and down-regulated (M, D11 vs P11_down) genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 11 d. (n), (o) GO enrichment analysis of up- (N, D14 vs P14_up) and down-regulated (O, D14 vs P14_down) genes in TaDOF4.7-B-OE vs PC186 empty vector at CIM 14 d.
Gene Ontology (GO) enrichment analysis was performed on the differentially expressed genes at CIM 3 d, CIM 5 d, CIM 8 d, CIM 11 d, and CIM 14 d, respectively. At CIM 3 d, the up-regulated DEGs were mainly enriched in the cytokinesis by cell plate formation (GO:0000911) and microtubule-based movement (GO:0007018), while the down-regulated DEGs were enriched in leaf development (GO:0048366) and regulation of multicellular organismal development (GO:2000026) (Fig. 2f, g, Supplementary Table S7). At CIM 5 d, the up-regulated DEGs were mainly enriched in lipid transport (GO:0006869) and plant-type hypersensitive response (GO:0009626), while the down-regulated DEGs were enriched in cellodextrin metabolic process (GO:2000889), and xylan catabolic process (GO:0045493) (Fig. 2h, i, Supplementary Table S8). At CIM 8 d, the up-regulated DEGs were mainly enriched in the response to external stress, such as response to stress (GO:0006950) and regulation of immune response (GO:0050777), while the down-regulated genes were enriched in multiple secondary metabolic processes, such as sucrose metabolic process (GO:0005985), and cation homeostasis (GO:0055080) (Fig. 2j, k, Supplementary Table S9). CIM 11 d represents the callus proliferation process. At this stage, the up-regulated genes were mainly enriched in auxin-activated signaling pathway (GO:0009734), cell division (GO:0051301), DNA replication (GO:0006260), while the down-regulated genes were mostly enriched in response to external stimulus (GO:0009605), and seed maturation (GO:2000034) (Fig. 2l, m, Supplementary Table S10). Notably, at CIM 11 d, the down-regulated genes were enriched in cytokinin dehydrogenase activity (GO:0019139) (Fig. 2m). These results implied that TaDOF4.7-B promoted wheat shoot regeneration by regulating the concentration or signal transduction of auxin and cytokinin. CIM 14 d is the stage of shoot primordia formation. At this stage, the up-regulated genes were mainly enriched in shoot system morphogenesis (GO:0010016), meristem maintenance (GO:0010073), and cell proliferation (GO:0008283), while the expression levels of genes related to environmental stress response and seed maturation process were down-regulated (Fig. 2n, o, Supplementary Table S11). These results implied that TaDOF4.7-B might induce early embryonic cell fate transition by enhancing the response to external stimuli and repressing the expression of seed development genes. In the following period, it may promote cell division and differentiation by regulating the cytokinin and auxin pathways, thereby facilitating the regeneration of wheat shoots.
TaDOF4.7-B promoted shoot regeneration by upregulating the expression levels of multiple regeneration-related factors and enhancing auxin signal transduction
-
Due to the significant changes in gene expression after CIM 8 d, and taking the key stages in wheat genetic transformation (the rapid proliferation of callus tissue at CIM 8 d, the initiation of shoot primordia at CIM 11 d, and the formation of distinct shoot structures at CIM 14 d) into consideration, to further analyze the mechanism by which TaDOF4.7-B regulates wheat shoot regeneration, K-means analysis was performed on the expression levels of the genes identified by the transcriptome data at CIM 8 d, CIM 11 d, and CIM 14 d. Based on the gene expression patterns, the genes were classified into six clusters (Fig. 3a, Supplementary Table S12). In Cluster 3, the expression levels of genes were significantly elevated in the TaDOF4.7-B-OE callus at CIM 11 d and CIM 14 d compared with the control (Fig. 3b). GO analysis of the genes in Cluster 3 revealed that these genes were primarily enriched in biological processes such as regulation of cell cycle (GO:0007049), regulation of meristem growth (GO:0010075), meristem initiation (GO:0010014), and stem cell population maintenance (GO:0019827) (Fig. 3c, Supplementary Table S13). Notably, multiple genes that have been reported to substantially improve plant regeneration capability were also enriched in this cluster. RT-qPCR results demonstrated that the well-reported regeneration factors, such as TaWOX5 and TaGIF1, were induced in D14 (Supplementary Fig. S3). To determine whether TaDOF4.7-B acts as a direct regulator of these regeneration-related factors, chromatin immunoprecipitation followed by qPCR (ChIP-qPCR) experiments were conducted. The results indicated that TaDOF4.7-B could directly bind to the promoters of TaWOX5 and TaGIF1 (Fig. 3d, e). Furthermore, luciferase activation assays confirmed that TaDOF4.7-B directly bound to the promoters of TaWOX5, and enhanced its expression, supporting the role of TaDOF4.7-B in promoting the expression of multiple reported regeneration related factors during wheat regeneration (Fig. 3f).
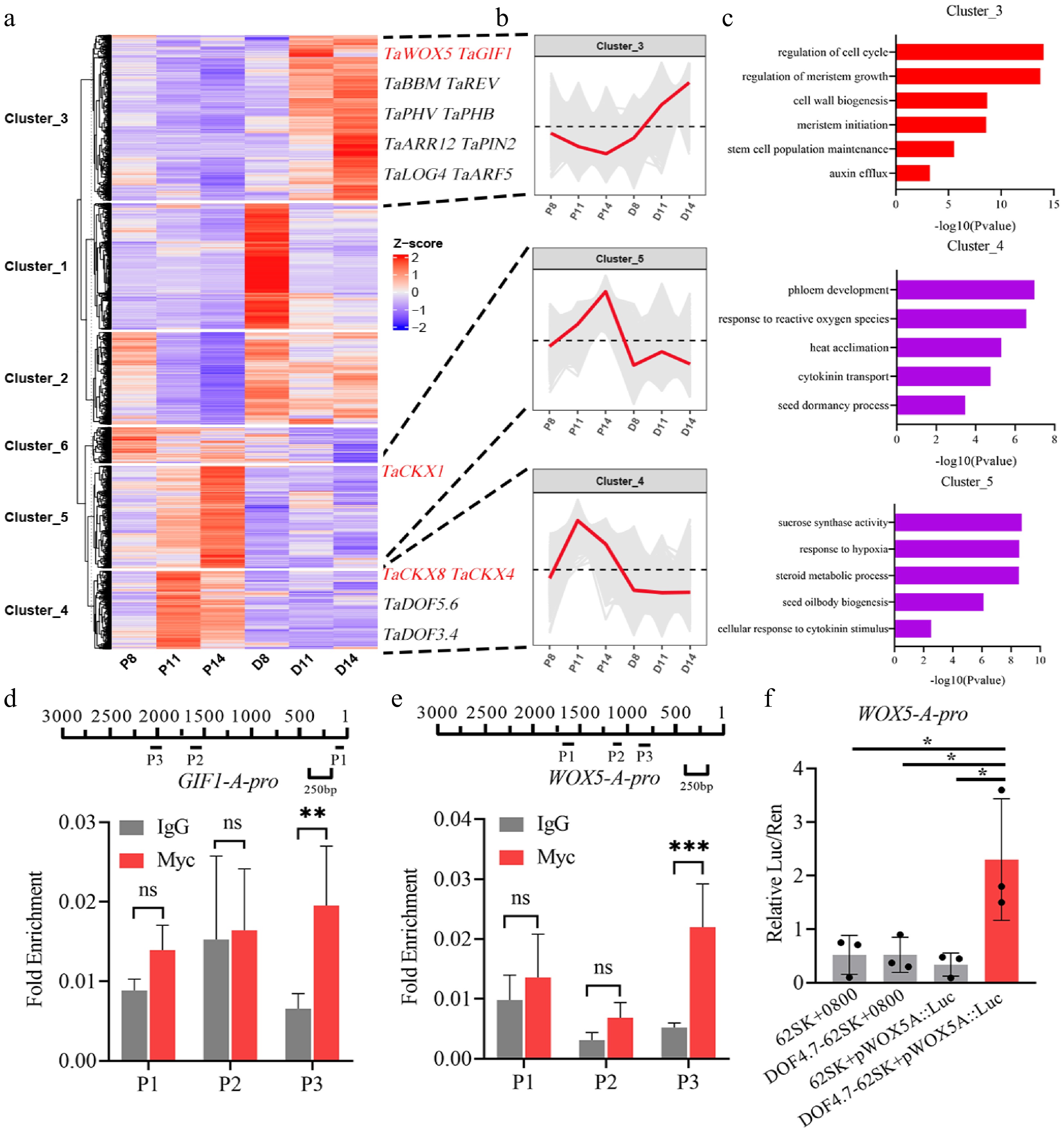
Figure 3.
TaDOF4.7-B promoted shoot regeneration by upregulating the expression levels of multiple regeneration related factors. (a), (b) K-means analysis based on the expression levels of the genes identified by the transcriptome data. (c) GO enrichment analysis of genes in cluster 3, cluster 4 and cluster 5. (d) The interaction between TaDOF4.7-B and the promoter of TaGIF1-A was confirmed by ChIP-qPCR. TaDOF4.7-B-OE samples with IgG antibody were used as negative controls. (e) The interaction between TaDOF4.7-B and the promoter of TaWOX5-A was confirmed by ChIP-qPCR. (f) The luciferase activity after transient expression TaDOF4.7-B and TaWOX5-Apro:LUC reporter in tobacco leaves. Values in (d)–(f) are means ± SE from three independent experiments (n = 3). Asterisks indicate significant differences (* p < 0.05, ** p < 0.01, *** p < 0.001) based on 2-tailed Student's t-tests.
Auxin is a crucial hormone that regulates plant shoot regeneration[37]. In Cluster 3, the key genes in auxin biosynthesis or auxin signaling transduction, such as PIN2 and multiple Auxin Response Factor (ARF) transcription factors, were also identified (Supplementary Fig. S4a, Supplementary Table S12). The results of RT-qPCR showed that the expression levels of several ARFs, such as TaARF5, TaARF11, and TaARF16, were significantly increased in D11 (Supplementary Fig. S4b). However, the ChIP-qPCR results indicated that TaDOF4.7-B could not directly bind to the promoters of the screened PIN2 and ARFs (Supplementary Fig. S4c−g). These results indicate that TaDOF4.7-B may indirectly regulate the auxin signaling pathway through unknown pathways.
Overexpression of TaDOF4.7-B promotes wheat shoot regeneration by regulating cytokinin homeostasis
-
Based on the results of K-means analysis, the genes in Cluster 4 and Cluster 5 showed gradually decreased expression patterns in D11 and D14 (Fig. 3a, b). The results of GO enrichment analysis revealed that the genes in Cluster 4 were mainly enriched in phloem development (GO:0010088), cytokinin transport (GO:0010184), and seed dormancy process (GO:0010162) (Fig. 3c, Supplementary Table S14). The genes in Cluster 5 were mainly enriched in sucrose synthase activity (GO:0016157), steroid metabolic process (GO:0008202), seed oilbody biogenesis (GO:0010344), and cellular response to cytokinin stimulus (GO:0071368) (Fig. 3c, Supplementary Table S15). In PureWheat-mediated transformation, the presence of cytokinin is essential for inducing callus formation and promoting shoot regeneration[23]. In Cluster 4 and Cluster 5, multiple CKX genes were identified (Supplementary Fig. S5a and Supplementary Table S12). CKXs can affect the cytokinin content by regulating its degradation, thereby playing roles in regulating shoot regeneration. The results of RT-qPCR showed that the expression levels of several CKX genes, such as TaCKX1-A/B/D and TaCKX4-A, significantly decreased in TaDOF4.7-B-OE calli (Fig. 4a). The promoter sequences of the screened TaCKXs were cloned. The motif analysis results showed that there were multiple binding sites for the DOF transcription factors in the promoters of these TaCKXs (Supplementary Fig. S5b). ChIP-qPCR was also employed to analyze the binding of TaDOF4.7-B to the TaCKXs promoters. The results indicated that TaDOF4.7-B could directly bind to the promoters of TaCKX1-A/B/D and TaCKX4-A (Fig. 4b−e). Luciferase activation assays further confirmed that TaDOF4.7-B can directly bind to the promoters of the screened TaCKX1 and inhibit its expression (Fig. 4f). These results indicated that TaDOF4.7-B may promote wheat shoot regeneration by inhibiting the expression of TaCKXs and regulating cytokinin homeostasis.
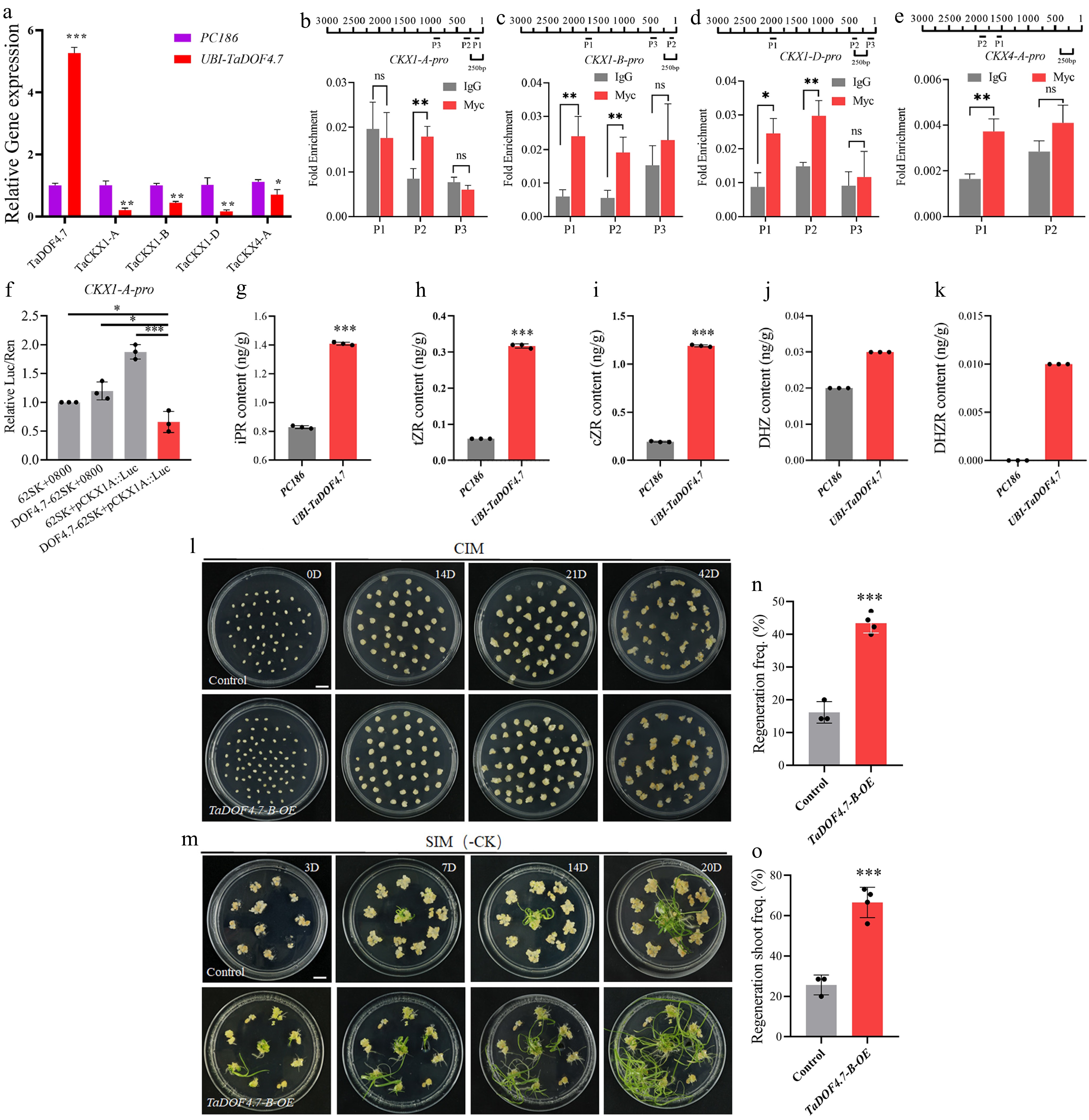
Figure 4.
Overexpression of TaDOF4.7-B promotes wheat shoot regeneration by regulating cytokinin homeostasis. (a) The expression levels of selected CKXs in TaDOF4.7-B-OE transgenic calli and empty vector transgenic calli. Values in (a) are means ± SE from three independent experiments (n = 3). (b)–(e) The interaction between TaDOF4.7-B and the promoters of the selected CKXs was confirmed by ChIP-qPCR. TaDOF4.7-B-OE samples with IgG antibody were used as negative controls. Values in (b)–(e) were means ± SE from three independent experiments (n = 3). (f) The luciferase activity after transient expression TaDOF4.7-B and TaCKX1-Apro:LUC reporter in tobacco leaves. Values in (f) were means ± SE from three independent experiments (n = 3). (g)–(k) The content of cytokinins measured in UBI-TaDOF4.7 calli and PC186 calli. (l), (m) Shoot regeneration phenotypes of immature embryos transformed with the empty vector (control) or Ubi::TaDOF4.7-B-6MYC vector (TaDOF4.7-B-OE) on a SIM without cytokinin. CIM, callus induction medium; SIM, shoot induction medium. Scale bar, 1 cm. (n), (o) Regeneration frequencies and regenerating shoot frequency of immature embryos transformed with control or Ubi::TaDOF4.7-B-6MYC vector. Values in (n), (o) are means ± SE from three independent experiments (n ≥ 3). Asterisks indicate significant differences (* p < 0.05, **p < 0.01, *** p < 0.001) based on 2-tailed Student's t-tests.
To verify whether TaDOF4.7-B promotes the regeneration ability of wheat by affecting cytokinin homeostasis, the content of cytokinin in calli induced from immature embryos that had been transformed with the empty vector (PC186) or Ubi::TaDOF4.7-B-6MYC vector (UBI-TaDOF4.7) was measured. The results showed that the content of various cytokinins in UBI-TaDOF4.7 calli was significantly higher than that in PC186 calli (Fig. 4g−k). Furthermore, the Ubi::TaDOF4.7-B-6MYC vector was used to transform immature embryos of Fielder wheat, and the transformed embryos were cultured on SIM without cytokinin. The results demonstrated that, compared with the PC186 empty vector control, the wheat immature embryos transformed with the Ubi::TaDOF4.7-B-6MYC vector still maintained higher regeneration frequency and regenerating shoot frequency (Fig. 4l−o). These results indicated that overexpression of TaDOF4.7-B promotes wheat shoot regeneration by regulating cytokinin homeostasis.
T2-generation of TaDOF4.7-B overexpression transgenic wheat still had a highly regenerative capacity
-
To further clarify the role of TaDOF4.7-B in the shoot regeneration process of wheat, overexpression TaDOF4.7-B in transgenic wheat lines were generated (Supplementary Fig. S6). And the shoot regeneration ability of the T2-generation TaDOF4.7-B overexpression transgenic wheat was measured. The results indicated that the shoot regeneration ability was significantly higher in TaDOF4.7-B overexpression transgenic wheat than that of Fielder (Fig. 5a). The regeneration frequency and regenerating shoot frequency were 3.83 and 3.91 times higher than those of the Fielder wheat plants, respectively (Fig. 5b, c).
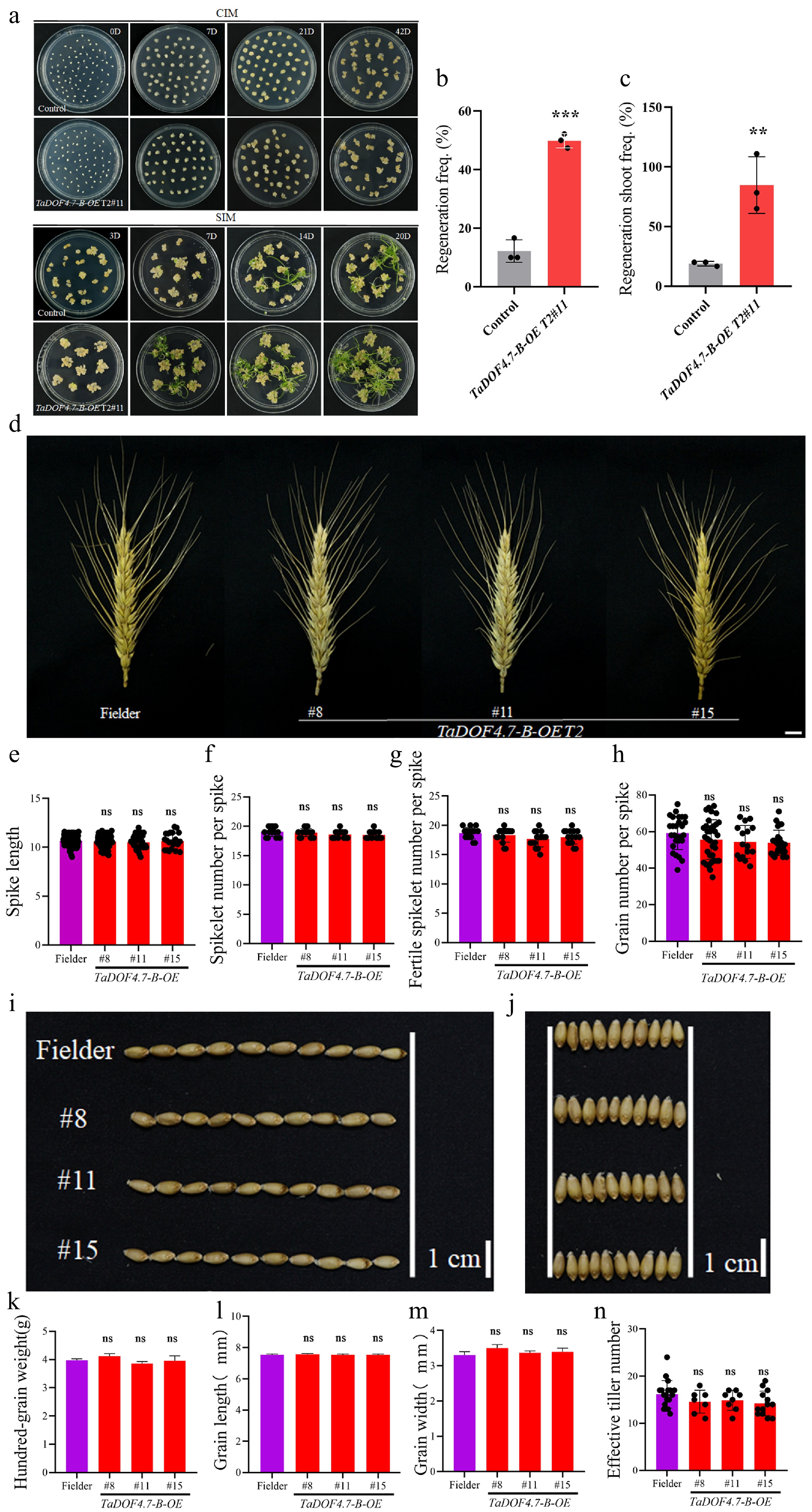
Figure 5.
T2-generation overexpression TaDOF4.7-B transgenic wheat still had a highly regenerative capacity. (a) Shoot regeneration phenotypes of overexpression TaDOF4.7-B transgenic wheat and control wheat. CIM, callus induction medium; SIM, shoot induction medium. Scale bar, 1 cm. TaDOF4.7-B T2#11, T2-generation overexpression TaDOF4.7-B transgenic wheat. (b), (c) Regeneration frequencies and regenerating shoot frequency of overexpression TaDOF4.7-B transgenic wheat and control wheat. Values in (b), (c) are means ± SE from three independent experiments (n = 3). Asterisks indicate significant differences (* p < 0.05, ** p < 0.01, *** p < 0.001) based on 2-tailed Student's t-tests. The yield traits, such as (d), (e) spike length, (f) spikelet number per spike, (g) fertile spikelet number per spike, (h) grain number per spike, (i), (l) grain length, (j), (m) grain width, (k) hundred-grain weight, and (n) effective tiller number, between overexpression TaDOF4.7-B transgenic wheat and control wheat. TaDOF4.7-B T2 #8, #11, and #15, T2-generation overexpression TaDOF4.7-B transgenic wheat. Values in (e)−(h), (n) are means ± SE from three independent experiments (n ≥ 10). Values in (k)–(m) are means ± SE from three independent experiments (n = 3). Asterisks indicate significant differences (* p < 0.05, ** p < 0.01, *** p < 0.001) based on 2-tailed Student's t-tests.
To elucidate the effect of TaDOF4.7-B overexpression on wheat growth and development, T2-generation transgenic lines overexpressing TaDOF4.7-B and Fielder wheat were cultivated in the experimental field, and various agronomic traits were documented. The results indicated that there were no significant differences in traits closely associated with yield, including spike length (Fig. 5d, e), spikelet number per spike (Fig. 5f), fertile spikelet number per spike (Fig. 5g), grain number per spike (Fig. 5h), grain length (Fig. 5i, l), grain width (Fig. 5j, m), hundred-grain weight (Fig. 5k), and effective tiller number (Fig. 5n) between the TaDOF4.7-B overexpression transgenic wheat lines and Fielder plants.
-
Wheat, a globally important staple crop, has been the focus of extensive research aimed at enhancing its genetic transformation efficiency by improving its regeneration ability[3]. Screening key factors that can enhance wheat regeneration ability and deciphering their molecular mechanisms are crucial for wheat breeding. In this study, a transcription factor, TaDOF4.7-B was identified, which can significantly improve the regeneration efficiency of wheat, and the molecular mechanism by which this transcription factor regulates plant regeneration was investigated through modulating the expression of multiple regeneration related factors, the auxin signaling pathway, especially cytokinin homeostasis.
Wheat shoot regeneration is a complex biological process and a critical determinant of the genetic transformation efficiency of wheat. To improve the genetic transformation efficiency of wheat, researchers are committed to screening key genes that can notably enhance wheat regeneration ability. The DOF transcription factor family is unique to plants, and plays important roles in regulating various biological processes[18]. In recent years, the functions of DOF transcription factors in regulating the growth and development of gramineous plants have received widespread attention. In rice, a DOF transcription factor, GLW9/OsDOF25, reduced the tillering angle by promoting the expression of OsPIN1b, and regulated grain shape by promoting the expression of EXPA6[38]. Another DOF transcription factor, OsDLT3, regulates tiller number by maintaining the expression of OsMOC1, OsMOC3, and OsFON1[39]. Through analyzing the wheat phytohormone metabolic regulatory network, TaDOF3A and TaDOF5.6B were considered as the major transcription factors on the metabolic regulation of cytokinin[40]. Recent studies have found that DOF transcription factors also play important roles in improving plant regeneration efficiency and may be key regulators for enhancing the regeneration ability of crops. Multi-omics data analysis was employed to reveal the transcriptional regulatory network responsible for shoot regeneration in wheat, and it was found that the transcriptional regulatory network driving shoot regeneration in wheat was controlled by 446 key transcription factors[15]. Among these 446 transcription factors, two genes, TaDOF5.6 and TaDOF3.4, were identified to be significantly upregulated during the early stages of wheat regeneration and maintain a high level of chromatin accessibility. Overexpression of TaDOF5.6 and TaDOF3.4 significantly improved the genetic transformation efficiency and shoot regeneration ability of wheat varieties such as Fielder, JM22, and KN199[15]. In this study, a DOF transcription factor gene, TaDOF4.7-B, with high chromatin accessibility during the early stage of wheat callus induction was screened. The TaDOF4.7-B was specifically expressed in the compact cell clusters of wheat callus and the regions where shoot primordia might emerge. Overexpression of TaDOF4.7-B can significantly increase the regeneration frequency, regenerating shoot frequency, and callus proliferation frequency of Fielder.
To analyze the mechanism by which TaDOF4.7-B promotes wheat regeneration, callus materials were collected at CIM 8 d, CIM 11 d, and CIM 14 d after transforming Fielder wheat immature embryos with the Ubi::TaDOF4.7-B-6MYC vector for RNA-sequencing. Transcriptome sequencing and ChIP-qPCR results showed that TaDOF4.7-B can directly bind to the promoters of TaWOX5 and TaGIF1. The GRF4-GIF1 fusion protein can improve the regeneration efficiency and genetic transformation efficiency of wheat[13]. TaWOX5 can improve the regeneration ability of multiple wheat varieties such as Fielder, KN199, JM22, and Ningchun4, thereby expanding the wheat genotypes that can undergo genetic transformation[14]. These results indicated that TaDOF4.7-B may affect wheat regeneration by regulating the expression of regeneration related factors. Previous study indicated that TaDOF5.6 and TaDOF3.4, which belong to the same family as TaDOF4.7-B, could improve wheat regeneration[15]. Based on the present results, TaDOF4.7-B can also promote wheat shoot regeneration. Therefore, it was speculated that their functions in regeneration might be redundant. Interestingly, transcriptome sequencing results showed that compared with the control, the expression levels of TaDOF5.6 and TaDOF3.4 were significantly lower (in cluster 4 of K-means) in the immature embryos of Fielder wheat transformed with the Ubi::TaDOF4.7-B-6MYC vector. This result implies that TaDOF5.6, TaDOF3.4, and TaDOF4.7-B might have mutual inhibition due to functional redundancy.
Plant hormones play a crucial role in regulating the regeneration process of crops[41]. Targeted manipulation of hormone signaling pathways has been shown to enhance shoot regeneration. ARF transcription factors are one of the most important regulatory factors of the plant auxin signaling pathway[42]. The transcriptome data in the present study showed that after transforming Fielder wheat immature embryos with the Ubi::TaDOF4.7-B-6MYC vector, the expression levels of multiple ARF transcription factors were significantly increased. However, ChIP-qPCR results showed that TaDOF4.7-B cannot directly bind to the promoters of the screened TaARFs. These results implied that TaDOF4.7-B promotes shoot regeneration of wheat by indirectly regulating the response to auxin signaling.
Cytokinin also plays a vital role in the plant regeneration process[43]. By optimizing the concentration and combination of different cytokinins, the regeneration efficiency of plants can be significantly improved. Cytokinin oxidase/dehydrogenase (CKX) irreversibly degrades cytokinin in plants[44]. Overexpression of CKXs led to a decrease in endogenous cytokinin levels and various plant development defects. Analysis of the transcriptome data in this study found that the expression levels of multiple CKXs in TaDOF4.7-B-OE callus were significantly lower than those in the control. ChIP-qPCR and luciferase activation assays confirmed that TaDOF4.7-B specifically binds to the promoter of TaCKXs and inhibits their expression. The endogenous cytokinin content in TaDOF4.7-B overexpression transgenic wheat was significantly higher than that in the control. Even without exogenous cytokinin application, overexpression of TaDOF4.7-B still maintained a high shoot regeneration ability. Therefore, these results suggest that TaDOF4.7-B regulates the cytokinin content in wheat by suppressing the expression of TaCKXs, thereby promoting shoot regeneration in wheat.
In conclusion, regeneration represents the primary bottleneck for efficient transformation and genome editing of wheat. Screening for key factors that can significantly enhance the regeneration ability of wheat is of crucial importance for advancing the process of wheat biological breeding. The present findings provide a novel regeneration factor, TaDOF4.7-B, for improving the regeneration efficiency of wheat, and clarify the mechanism by which DOF transcription factors affect the shoot regeneration ability of plants by regulating the expression of TaWOX5 and TaGIF1, auxin signaling transduction, especially cytokinin homeostasis.
We would like to thank Prof. Jiajie Wu and Prof. Fei Ni (College of Agronomy, Shandong Agricultural University) for providing the PC186 vectors. This research was funded by the Biological Breeding-National Science and Technology Major Project (2024ZD04077).
-
The authors confirm their contributions to the paper as follows: research design: Su Y, Wang C, Wang YP; research performing: Wang YP, Zheng J, Peng J; research assistance: Wu J, Zhang Z; data analysis: Wang YP; manuscript writing: Wang C, Yu H. All authors reviewed the results and approved the final version of the manuscript.
-
The RNA-seq data discussed in this study have been deposited at the National Genomics Data Center Genome Sequence Archive (GSA; https://ngdc.cncb.ac.cn/gsa/) under Accession No. PRJCA039631. Gene sequence data mentioned in this study can be found at Wheatomics 1.0 (http://wheatomics.sdau.edu.cn/) under the following Accession Nos: TaDOF4.7-A (TraesCS4A02G234000), TaDOF4.7-B (TraesCS4B02G081500), TaDOF4.7-D (TraesCS4D02G080100).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yi Peng Wang, Jiahao Zheng, Jing Peng
- Supplementary Table S1 The primers used in this study.
- Supplementary Table S2 The list of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 3d (D3 vs P3).
- Supplementary Table S3 The list of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 5d (D5 vs P5).
- Supplementary Table S4 The list of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D8 vs P8).
- Supplementary Table S5 The list of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D11 vs P11).
- Supplementary Table S6 The list of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D14 vs P14).
- Supplementary Table S7 Go enrichment analysis of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 3d (D3 vs P3).
- Supplementary Table S8 Go enrichment analysis of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 5d (D5 vs P5).
- Supplementary Table S9 Go enrichment analysis of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D8 vs P8).
- Supplementary Table S10 Go enrichment analysis of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D11 vs P11).
- Supplementary Table S11 Go enrichment analysis of DEGs in TaDOF4.7-B-OE vs. PC186 empty vector at CIM 8d (D14 vs P14).
- Supplementary Table S12 The list of genes in each cluster in K-means.
- Supplementary Table S13 Go enrichment analysis of genes in cluster 3.
- Supplementary Table S14 Go enrichment analysis of genes in cluster 4.
- Supplementary Table S15 Go enrichment analysis of genes in cluster 5.
- Supplementary Fig. S1 The chromatin accessibility of the promoters of TaDOF4.7-A/B/D in wheat during the early stage of callus induction using the published data.
- Supplementary Fig. S2 RNA-sequencing for immature embryos of Fielder wheat after transforming Ubi::TaDOF4.7-B-6MYC vector or PC186 empty vector at CIM 3d, CIM 5d, CIM 8d, CIM 11d, CIM 14d.
- Supplementary Fig. S3 The expression levels of TaWOX5 and TaGIF1 in the callus after transformation Fielder wheat immature embryos with the Ubi::TaDOF4.7-B-6MYC vector.
- Supplementary Fig. S4 TaDOF4.7-B indirectly regulate the auxin signaling pathway to promote wheat regeneration.
- Supplementary Fig. S5 The expression level and motif analysis of the selected CKXs.
- Supplementary Fig. S6 Identification of putative T0 progeny of TaDOF4.7-B-OE transgenic lines in Fielder by amplification of transgenic-specific PCR product (a) and RT-qPCR analysis (b).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang YP, Zheng J, Peng J, Wu J, Zhang Z, et al. 2025. TaDOF4.7-B enhances wheat regeneration via regulating cytokinin homeostasis. Seed Biology 4: e019 doi: 10.48130/seedbio-0025-0018 

TaDOF4.7-B enhances wheat regeneration via regulating cytokinin homeostasis
- Received: 11 June 2025
- Revised: 05 August 2025
- Accepted: 25 August 2025
- Published online: 03 November 2025
Abstract: Wheat is a globally important staple crop. However, the weak regeneration ability of wheat significantly restricts the progress of wheat molecular design breeding. Screening for key factors that can enhance the regeneration ability of wheat is crucial for the development of the wheat industry. In this study, a DOF transcription factor, TaDOF4.7-B, was identified as a crucial factor regulating the wheat regeneration ability. Transcriptome analysis results indicated that TaDOF4.7-B could improve the regeneration ability of wheat by directly promoting the expression of regeneration related genes, such as TaWOX5, or indirectly regulating the expression level of TaARFs. Importantly, combined with the experimental results of ChIP-qPCR and luciferase activation assays, TaDOF4.7-B can specifically bind to the promoters of multiple TaCKXs and inhibit their expression. The cytokinin concentration in overexpression TaDOF4.7-B wheat was significantly higher than that in the control. On the SIM medium without cytokinin, the wheat immature embryos transformed with the Ubi::TaDOF4.7-B-6MYC vector maintained a higher regeneration frequency and regeneration shoot frequency. Furthermore, the T2-generation of TaDOF4.7-B overexpression transgenic wheat lines still hold a highly regenerative capacity. The present findings demonstrate a novel regeneration factor, TaDOF4.7-B, and shed light on the mechanism by which DOF transcription factors promote plant regeneration through regulating cytokinin homeostasis.
-
Key words:
- TaDOF4.7-B /
- Regeneration /
- Cytokinin /
- Cytokinin oxidase/dehydrogenase /
- Wheat