









-
Madagascar periwinkle (Catharanthus roseus [L.] G. Don) (MP) is a perennial herbaceous plant that belongs to the Apocynaceae family, which is indigenous to Madagascar, subsequently introduced and naturalized in multiple countries worldwide due to its medicinal and ornamental value[1]. Owing to its rapid growth rate and minute seeds readily dispersed by insects, wind, and water currents, the periwinkle has been designated as an invasive species in numerous Asian, African, American, and Oceanian nations[2]. Historically, C. roseus has garnered considerable scientific interest for its ability to biosynthesize over 150 monoterpenoid indole alkaloids (MIAs), including vinblastine and vincristine, which serve as principal sources of these pharmacologically important compounds[3]. Extracts derived from this plant have demonstrated therapeutic potential in treating certain cardiovascular conditions and specific types of cancer (Fig. 1)[4].
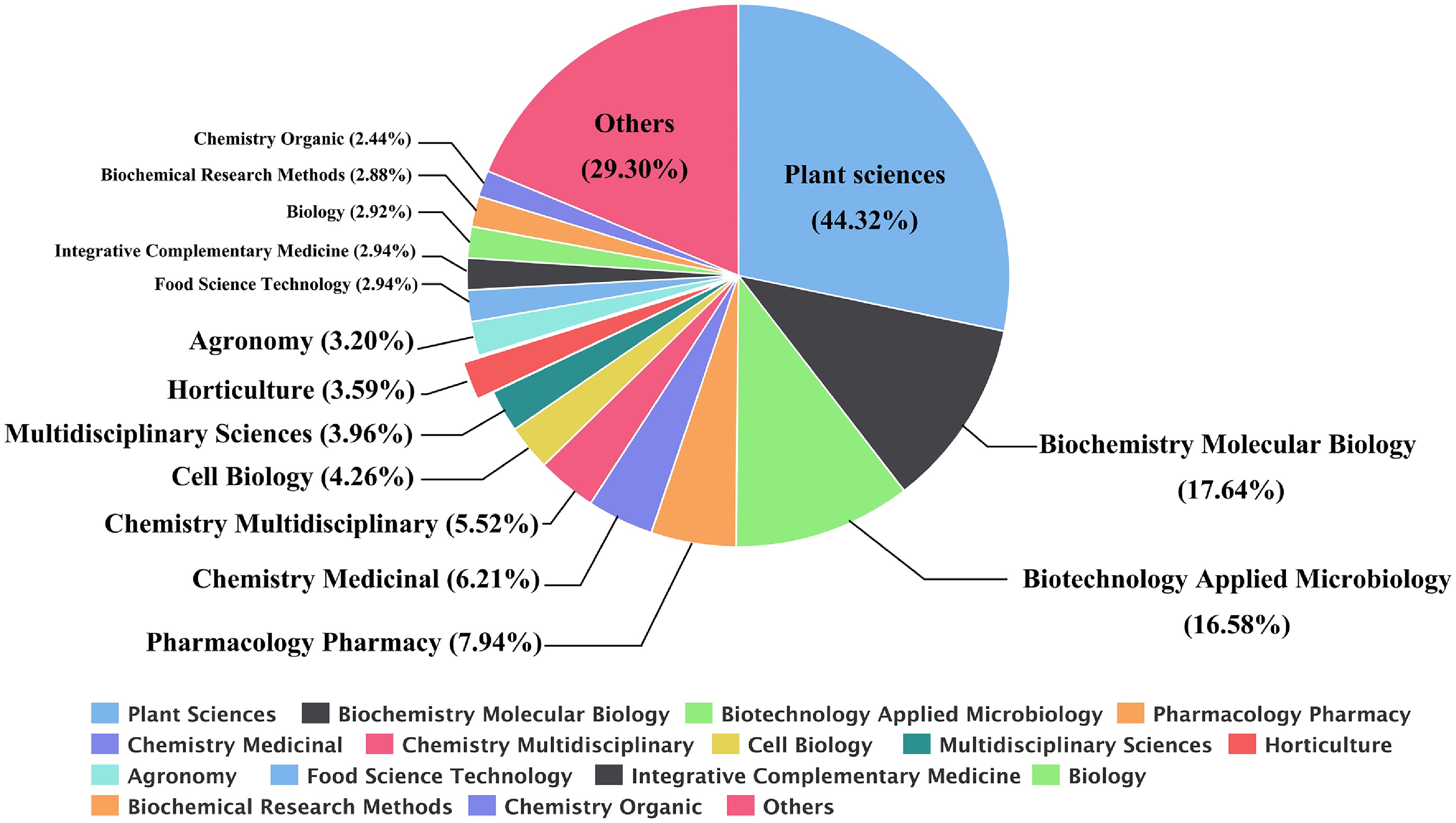
Figure 1.
Statistics of Catharanthus roseus research publications (1963–2025). Distribution (%) of publications on periwinkle in different subject areas during the period 1963–2025 (search with all fields, query preview = 'Madagascar periwinkle' OR 'Catharanthus roseus' at the Web of Science).
Beyond its medicinal applications, C. roseus exhibits efficient vegetative propagation predominantly by seeds, and less by the stem cuttings, facilitating its widespread cultivation as an ornamental plant in tropical and subtropical climates. This popularity is attributed to its prolonged flowering period, diverse floral coloration, tolerance to heat and drought stress, and overall adaptability[5,6]. The periwinkle breeders have circumvented the reproductive constraint of cleistogamy in C. roseus, focused on breeding for dwarf or semidwarf, contrasting colored corolla with its eye color, floriferous, high biomass, resistance to aerial blight and dieback diseases, heat and drought tolerance as the prime targets to develop new cultivars for wider horticultural adoption[7,8], such as the 'Cora® XDR' series of Syngenta® flowers (the USA), and the Vinca 'Titan-ium™' series of PanAmerican Seed (the USA). However, in contrast to the well-established pharmaceutical breeding programs for Catharanthus, the development of ornamental cultivars remains hindered by continued dependence on traditional hybridization techniques, the lack of high-throughput marker-assisted selection (MAS) platforms, and insufficient integrated pest management (IPM) strategies targeting major pathogens and insect pests. These challenges constrain the genetic improvement and broader horticultural deployment of ornamental MP varieties.
Consequently, this review systematically assesses the current status of ornamental MP germplasm resources and genetic breeding, predominantly highlighting the genetic regulation of preliminary traits and the limitations of current breeding methodologies. Furthermore, the review explores prospective avenues for enhancing stress resistance and optimizing complex trait integration, thereby contributing to the advancement of both theoretical knowledge and practical approaches for the improvement of ornamental Catharanthus cultivars.
-
Linnaeus[6] initially established the genus Vinca in 1753, describing V. minor and V. major. In 1759, he incongruously included the tropical V. rosea within this temperate genus, despite morphological inconsistencies in stamen structure. Reichenbach first recognized the generic distinctness of V. rosea from Vinca. Subsequently, George Don redefined the taxonomy: retaining Vinca for V. minor, V. major, and V. herbacea, while erecting the genus Catharanthus with C. roseus (basionym: Vinca rosea L.) as its type species[9]. Consequently, Catharanthus and Vinca constitute closely related genera, with C. roseus historically classified as Vinca rosea. The genus name Catharanthus is derived from the Greek words katharos (pure) and anthos (flower), collectively meaning 'pure flower'; then the specific epithet roseus originates from Latin, denoting 'red', 'rose-like', or 'rosy' in color[10].
Madagascar periwinkle (Catharanthus roseus) is considered a native of the West Indies, but it was originally described from Madagascar[11]. Initially, MP was introduced into Paris in 1757 and has since become naturalized in continental Africa, America, Asia, Australia, and Southern Europe, as well as on some islands in the Pacific Ocean (Fig. 2)[12]. Subsequently, periwinkle is commonly grown as an ornamental plant throughout tropical and subtropical regions of the world (Fig. 2a)[13]. All Catharanthus species in China are introduced rather than native, with no recorded native populations. The C. roseus, as the only species of this genus distributed in China, has become naturalized in southern China (such as Yunnan, Guangdong, and Fujian province), and some eastern provinces (such as Zhejiang, Jiangxi, and Taiwan province) (Fig. 2b).
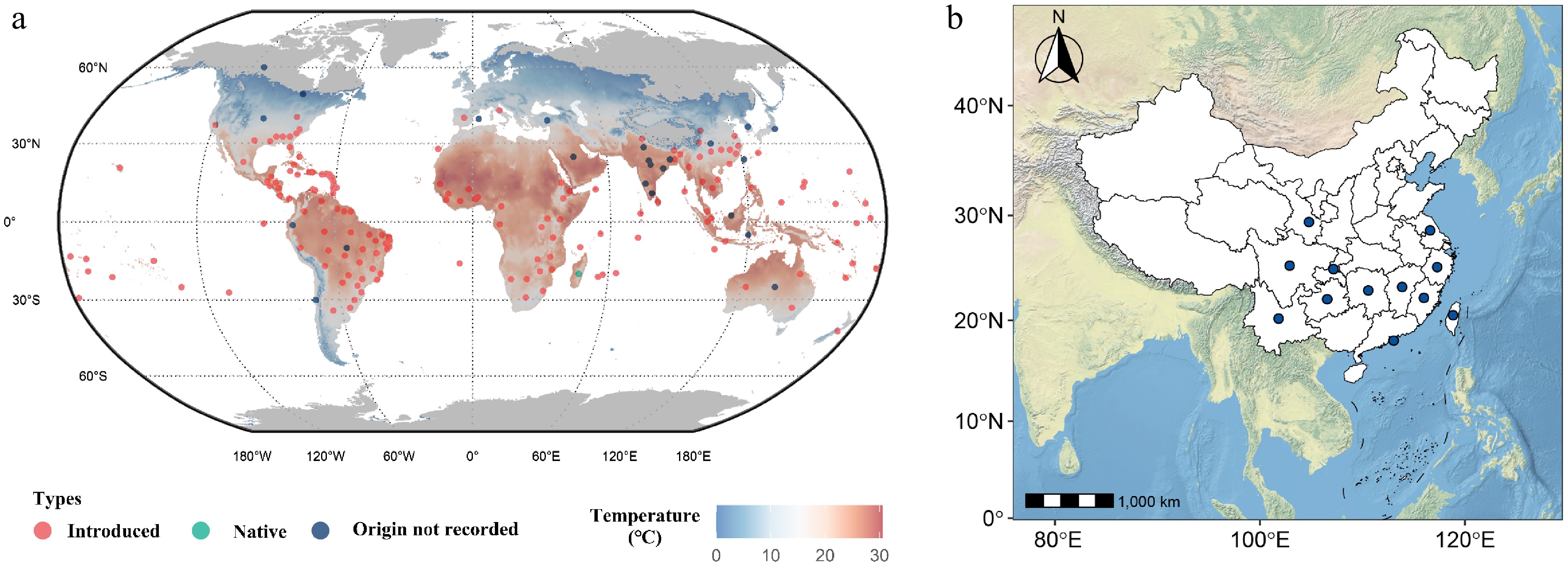
Figure 2.
Distribution map of Catharanthus roseus based on all the information available to CABI Compendium (doi:
10.1079/cabicompendium.16884 ). (a) Distribution map of C. roseus across the globe. (b) Distribution map of C. roseus across China. The sample distribution map was generated using the R package 'rnaturalearth' and RStudio (Version 2025.05.1+513).Wild species resources
-
The Catharanthus is a genus of herbaceous perennial or annual plants, belonging to the Apocynaceae family[14,15]. Moreover, eight species out of the Catharanthus genus are indigenous to Madagascar (C. trichophyllus, C. lanceus, C. coriaceus, C. ovalis, C. longifolius, C. scitulus, and C. roseus), and one (C. pusillus) is restricted to India and Sri Lanka[2]. Previous cytogenetic analyses have established that all species within the genus Catharanthus exhibit a diploid chromosome number of 2n = 2x = 16[16−18]. Among the eight pairs of chromosomes, four pairs were identified with submedian centromeres, while two pairs exhibited subterminal centromeres, and the remaining two pairs possessed median centromeres[18,19].
Two distinct morphotypes of MP occur in the wild: one characterized by pink flowers and reddish stems, and another by white flowers and green stems[20,21]. These two forms are often found growing sympatrically. Furthermore, the wild species of MP exhibits remarkable tolerance to abiotic stresses such as drought and salinity (up to 2,000 ppm), enabling it to thrive in diverse habitats, including sandy soils, shrublands, grasslands, riverbanks, savanna dunes, wastelands, residential areas, roadsides, beaches, and limestone outcrops[22,23]. The extent of tolerance varies as per genotypes' adaptability in agro-geographical zones[24].
Cultivated germplasm resources
-
The MP has been bred as a horticultural plant since the 1920s[20]. Interestingly, the MP has been naturalized worldwide for medicinal applications, yet it is regarded as either an invasive weed or a decorative ornamental in different regions[25]. During the 1970s, breeding efforts for Madagascar periwinkle primarily focused on ornamental traits, particularly through natural selection for corolla color. Popular varieties included C. roseus cv. 'Rosea' (purple), C. roseus cv. 'Alba' (white), and a white-flowered form with a purple eye—often commercialized as mixed populations exhibiting substantial variation in alkaloid content[26]. Before the 1980s, systematic breeding programs had not yet been established, and pure lines were generally absent in C. roseus[27]. During the 1980s, breeding programs in MP were spearheaded largely by commercial horticultural seed companies, with an emphasis on creating novel recombinant lines through inter-cultivar hybridization and selective breeding. Furthermore, the primary objectives in breeding included the creation of novel combinations of corolla and eye zone coloration, improvements in leaf morphology (encompassing attributes such as shape, size, and pigmentation), and the reduction of plant height[28].
Therefore, horticulturists and seed companies have utilized conventional breeding techniques, particularly hybridization of C. roseus with related species, to generate F1 progeny over the past two decades. These efforts have led to the introduction of numerous horticulturally improved cultivars and varieties for bedding, pot culture, hanging basket, and mass effect[29,30]. Currently, there are over 250 known commercial varieties of periwinkle (Table 1). Furthermore, the breeding efforts have not only enhanced floral characteristics—such as expanding the color spectrum, increasing bloom quantity and size, and improving plant architecture—but have also augmented resistance to diseases and cooler climates, while simultaneously boosting herbage production and alkaloid yield (Fig. 3).
Table 1. The major varieties of Madagascar periwinkle.
Series Supplier Cultivars Blockbuster™ Syngenta® Flowers (originally developed by Floranova) 'Blockbuster™ Apricot', 'Blockbuster™ Blue', 'Blockbuster™ Blush', 'Blockbuster™ Burgundy', 'Blockbuster™ Crimson', 'Blockbuster™ Dark Red', 'Blockbuster™ Mix', 'Blockbuster™ Patriot Mix', 'Blockbuster™ Peppermint', 'Blockbuster™ Punch', 'Blockbuster™ Red With Eye', 'Blockbuster™ White' Carpet Sakata Ornamentals 'Rose Carpet', 'Pink Carpet', 'Magic Carpet', 'Dawn Carpet' Cobra Syngenta® Flowers (originally developed by Floranova) 'Cobra Apricot', 'Cobra Orchid Eye', 'Cobra Passion Fruit, 'Cobra Peppermint', 'Cobra Purple', 'Cobra Purple Eye', 'Cobra Red', 'Cobra Red Eye', 'Cobra Rose', 'Cobra Strawberry Red', 'Cobra White', 'Cobra Lavender Halo', 'Cobra Mix', 'Cobra Orange' Cooler Pan America Seed 'Cooler Apricot', 'Cooler Blush', 'Cooler Coconut', 'Cooler Deep Orchid', 'Cooler Grape', 'Cooler Hot Rose', 'Cooler Icy Pink', 'Cooler Lavender Halo', 'Cooler Mixture', 'Cooler Orchid', 'Cooler Orchid Deep', 'Cooler Peppermint', 'Cooler Peppermint Improved', 'Cooler Pink', 'Cooler Raspberry Red', 'Cooler Red', 'Cooler Rose', 'Cooler Strawberry' Cora® Cascade XDR Syngenta® Flowers 'Cora® Cascade XDR Apricot', 'Cora® Cascade XDR Bright Rose', 'Cora® Cascade XDR Lavender with Eye', 'Cora® Cascade XDR Lilac', 'Cora® Cascade XDR Mix', 'Cora® Cascade XDR Polka Dot', 'Cora® Cascade XDR Punch', 'Cora® Cascade XDR Shell Pink', 'Cora® Cascade XDR Strawberry', 'Cora® Cascade XDR Violet', 'Cora® Cascade XDR White' Cora® XDR Syngenta® Flowers 'Cora® XDR Apricot', 'Cora® XDR Cranberry', 'Cora® XDR Deep Strawberry', 'Cora® XDR Hotgenta', 'Cora® XDR Light Pink', 'Cora® XDR Magenta Halo', 'Cora® XDR Mix', 'Cora® XDR Orchid', 'Cora® XDR Pink Halo', 'Cora® XDR Polka Dot', 'Cora® XDR Red Glow', 'Cora® XDR Rose Punch', 'Cora® XDR White' First Kiss Benary 'First Kiss Apricot', 'First Kiss Blueberry', 'First Kiss Blush', 'First Kiss Cherry red', 'First Kiss Coral', 'First Kiss Icy Pink', 'First Kiss Orchid', 'First Kiss Peach', 'First Kiss Polka Dot', 'First Kiss Raspberry', 'First Kiss Rose', 'First Kiss Ruby', 'First Kiss Sunrise', 'First Kiss Think Pink', 'First Kiss White' Heat Wave HEM Genetics (originally developed by John Bodger & Sons Co.) 'Heatwave Apricot', 'Heatwave Blue W/Eye', 'Heatwave Burgundy', 'Heatwave Cherry', 'Heatwave Deep Rose', 'Heatwave Formula Mixture', 'Heatwave Grape', 'Heatwave Midnight Mix', 'Heatwave Orchid', 'Heatwave Peach', 'Heatwave Peppermint', 'Heatwave Pink', 'Heatwave Raspberry', 'Heatwave Red', 'Heatwave Rose', 'Heatwave Santa Fe', 'Heatwave Southwest Mix', 'Heatwave White' Jaio Murakami Seed Co., Ltd. 'Jaio Bicolor Deep Rose', 'Jaio Cherry Red', 'Jaio Dark Red', 'Jaio Vermillion', 'Jaio Vermillion Eye', 'Jaio Peppermint', 'Jaio White', 'Jaio Salmon Pink', 'Jaio Sky Blue' Little Syngenta® flowers 'Little Linda', 'Little Salmon', 'Little Blanche', 'Little Bright Eye', 'Little Delicata', 'Little Pinkie' Mediterranean XP Pan America Seed 'Mediterranean XP Burgundy Halo', 'Mediterranean XP Dark Red', 'Mediterranean XP Hot Rose', 'Mediterranean XP Peach', 'Mediterranean XP Red', 'Mediterranean XP Rose Halo', 'Mediterranean XP Strawberry', 'Mediterranean XP White', 'Mediterranean XP Mixture' Mega Bloom Ameriseed 'Mega Bloom Dark Red', 'Mega Bloom Polka Dot', 'Mega Bloom White', 'Mega Bloom Lavender', 'Mega Bloom Icy Pink', 'Mega Bloom Strawberry', 'Mega Bloom Burgundy with Eye', 'Mega Bloom Peach Pink', 'Mega Bloom Orchid Halo', 'Mega Bloom Grape', 'Mega Bloom Pink', 'Mega Bloom Red', 'Mega Bloom Raspberry', 'Mega Bloom Apricot' Nirvana® XDR Syngenta® Flowers 'Nirvana® XDR Watermelon', 'Nirvana® XDR Coral', 'Nirvana® XDR Apricot Splash', 'Nirvana® XDR Cranberry Halo', 'Nirvana® XDR Blackberry', 'Nirvana® XDR Blush Splash', 'Nirvana® XDR Blue Halo' Pacifica XP Pan America Seed 'Pacifica XP Apricot', 'Pacifica XP Blush', 'Pacifica XP Burgundy', 'Pacifica XP Burgundy Halo', 'Pacifica XP Cherry Red Halo', 'Pacifica XP Dark Red', 'Pacifica XP Deep Orchid', 'Pacifica XP Magenta Halo', 'Pacifica XP Orange', 'Pacifica XP Pink', 'Pacifica XP Polka Dot', 'Pacifica XP Punch', 'Pacifica XP Really Red', 'Pacifica XP Red Halo', 'Pacifica XP Rose Halo', 'Pacifica XP White', 'Pacifica XP Bold Mixture', 'Pacifica XP Mixture' Soirée® Monrovia 'Soirée® Cerise White Eye', 'Soirée® Hot Pink', 'Soirée® Pink White Eye', 'Soirée® Light Pink Dark Eye', 'Soirée® White' Soirée® Crown Monrovia 'Soirée® Crown Pink', 'Soirée® Crown Rose' Soirée® Double Suntory flowers 'Soirée® Double Appleblossom', 'Soirée® Double Pure White', 'Soirée® Double Pink Improved', 'Soirée® Double Orchid Improved', 'Soirée® Double White', 'Soirée® Double Pink', 'Soirée® Double Orchid' Soirée® Flamenco Suntory flowers 'Soirée® Flamenco Electric Salmon Eye', 'Soirée® Flamenco Strawberry Picotee', 'Soirée® Flamenco Plum Velvet', 'Soirée® Flamenco Salsa Red', 'Soirée® Flamenco Salmon Glow', 'Soirée® Flamenco Cheeky Pink', 'Soirée® Flamenco Senorita Pink', 'Soirée® Flamenco Pink Twist' Soirée® Kawaii Suntory flowers 'Soirée® Kawaii Hot Pink', 'Soirée® Kawaii Lilac Dream', 'Soirée® Kawaii Paprika Red', 'Soirée® Kawaii Paprika Red', 'Soirée® Kawaii Berry Blast', 'Soirée® Kawaii Lady Liberty', 'Soirée® Kawaii Blueberry Kiss', 'Soirée® Kawaii Coral Reef', 'Soirée® Kawaii Red Shades', 'Soirée® Kawaii White Peppermint', 'Soirée® Kawaii Light Purple', 'Soirée® Kawaii Pink', 'Soirée® Kawaii Lavender', 'Soirée® Kawaii Coral' Solar HEM Genetics 'Solar Apple Blossom', 'Solar Apricot', 'Solar Blueberry', 'Solar Blush Pink', 'Solar Cherry with Eye', 'Solar Formula Mixture', 'Solar Fresh Red', 'Solar Lilac', 'Solar Orange with Eye', 'Solar Orchid with Eye', 'Solar Pink', 'Solar Raspberry with Eye', 'Solar Red', 'Solar Red with Eye', 'Solar White' Stardust Pan America Seed 'Stardust Orchid', 'Stardust Pink', 'Stardust Mix', 'Stardust Rose' SunStorm® Syngenta® Flowers 'SunStorm® Apricot', 'SunStorm® Blush', 'SunStorm® Bright Red', 'SunStorm® Deep Lilac', 'SunStorm® Deep Orchid', 'SunStorm® Deep Orchid', 'SunStorm® Deep Pink', 'SunStorm® Light Blue', 'SunStorm® Mix', 'SunStorm® Orchid Halo', 'SunStorm® Pure White', 'SunStorm® Purple', 'SunStorm® Red', 'SunStorm® Red Halo', 'SunStorm® Rose With Eye', 'SunStorm® Tropical Mix', 'SunStorm® White With Eye' Tattoo™ Pan America Seed 'Tattoo™ Black Cherry', 'Tattoo™ Blueberry', 'Tattoo™ Orange', 'Tattoo™ Papaya', 'Tattoo™ Raspberry', 'Tattoo™ Tangerine', 'Tattoo™ American Pie Mixture' Titan-ium™ Pan America Seed 'Titan-ium™ Apricot', 'Titan-ium™ Blush', 'Titan-ium™ Dark Red', 'Titan-ium™ Polka Dot', 'Titan-ium™ Punch', 'Titan-ium™ Really Red', 'Titan-ium™ White', 'Titan-ium™ Mixture' Titan™ Pan America Seed 'Titan™ Apricot', 'Titan™ Blush', 'Titan™ Burgundy', 'Titan™ Cranberry', 'Titan™ Dark Red', 'Titan™ Icy Pink', 'Titan™ Lavender Blue Halo', 'Titan™ Lilac', 'Titan™ Polka Dot', 'Titan™ Punch', 'Titan™ Pure White', 'Titan™ Really Red', 'Titan™ Rose', 'Titan™ Rose Halo', 'Titan™ Bubble Gum Mixture', Titan™ 'Clear Mixture', 'Titan™ Mixture', 'Titan™ Summer Breeze Mixture' Tropicana University of Connecticut 'Tropicana Apricot', 'Tropicana Blush', 'Tropicana Bright Eye', 'Tropicana Pink', 'Tropicana Rose' Valiant™ Pan America Seed 'Valiant™ Apricot', 'Valiant™ Burgundy', 'Valiant™ Lilac', 'Valiant™ Magenta', 'Valiant™ Orchid', 'Valiant™ Punch', 'Valiant™ Pure White', 'Valiant™ Mixture' Victory Sakata Ornamentals 'Victory Apricot', 'Victory Blue', 'Victory Bright Eye', 'Victory Carmine', 'Victory Carmine Rose', 'Victory Cranberry', 'Victory Deep Apricot', 'Victory Deep Pink', 'Victory Grape', 'Victory Lavender', 'Victory Light Pink', 'Victory Pure White', 'Victory Purple', 'Victory Red', 'Victory Mix' Viper Sahola flowers 'Viper Apricot', 'Viper Grape', 'Viper Orchid Halo', 'Viper Pink', 'Viper Purple', 'Viper Purple Halo', 'Viper Red', 'Viper Red Eye', 'Viper Rose', 'Viper Watermelon' Virtuosa Sakata Ornamentals 'Virtuosa White', 'Virtuosa Polka Dot', 'Virtuosa Peach', 'Virtuosa Apricot', 'Virtuosa Deep Purple', 'Virtuosa Raspberry', 'Virtuosa Deep Red', 'Virtuosa Cranberry', 'Virtuosa Lavender', 'Virtuosa Pink', 'Virtuosa Punch', 'Virtuosa Rosa', 'Virtuosa Orchid', 'Virtuosa Mix' Vitesse Syngenta flowers (originally developed by Floranova) 'Vitesse Apricot', 'Vitesse Blue', 'Vitesse Dark Red', 'Vitesse Lavender Morn', 'Vitesse Orange', 'Vitesse Peppermint', 'Vitesse Pink', 'Vitesse Purple', 'Vitesse Rose', 'Vitesse Rose Morn', 'Vitesse White' 
Figure 3.
The abundant cultivars of flower color and shape in Madagascar periwinkle (Catharanthus roseus). (a) Different colors of MP cultivars`petals. (b) Different eye zone colors of MP cultivars. (c) Wavy petal of MP cultivars. (d) Double-flowered and tiny-flowered MP cultivars. (The publicly accessible websites for downloading these images are listed in Supplementary Table S1).
Furthermore, breeding objectives for mainstream cultivated varieties of periwinkle diverge significantly across regions. European and American breeders prioritize flower color, disease resistance, and growth habit, as exemplified by the 'Titan™' series (Pan America Seed), the 'Pacifica XP' series (Pan America Seed), and the 'Cora® Cascade XDR' series (Syngenta® Flowers). The remarkable diversity of commercial periwinkle varieties underscores significant achievements in targeted breeding. The 'Cora® XDR' series (Syngenta Flowers), for instance, is distinguished by its exceptional resistance to soil-borne pathogens such as Phytophthora and Pythium, making it a robust choice for landscape plantings in humid climates. Meanwhile, the 'Cora® Cascade' series (Syngenta Flowers) is characterized by a semi-trailing growth habit and prolific blooming, traits that render it ideally suited for hanging baskets and container gardens. Furthermore, Japanese breeding has placed a greater emphasis on floral innovation, as exemplified by the 'Soirée®' series (Suntory Flowers). This series includes the 'Soirée® Double' group, which produces fully double flowers with petaloid stamens, and the 'Soirée® Flamenco' group, noted for its distinctive wavy petals. These cultivars, detailed in Table 1, collectively demonstrate how specific market demands have driven the development of specialized traits in ornamental periwinkle.
-
Using the query preview (All fields = 'Madagascar periwinkle' OR 'Catharanthus roseus') a literature search was conducted for publications from nearly 60 years, spanning 1963 to 2025 (Fig. 1). The 4,348 retrieved publications showed that most studies focus on plant sciences (44.32%), followed by biochemistry and molecular biology (17.64%), and biotechnology applied to microbiology (16.58%). However, there is a notable lack of research on the horticultural aspects of MP (3.59%) (Fig. 1), which presents an important opportunity for further research into the mechanisms of molecular breeding and the development of ornamental traits (Fig. 4).
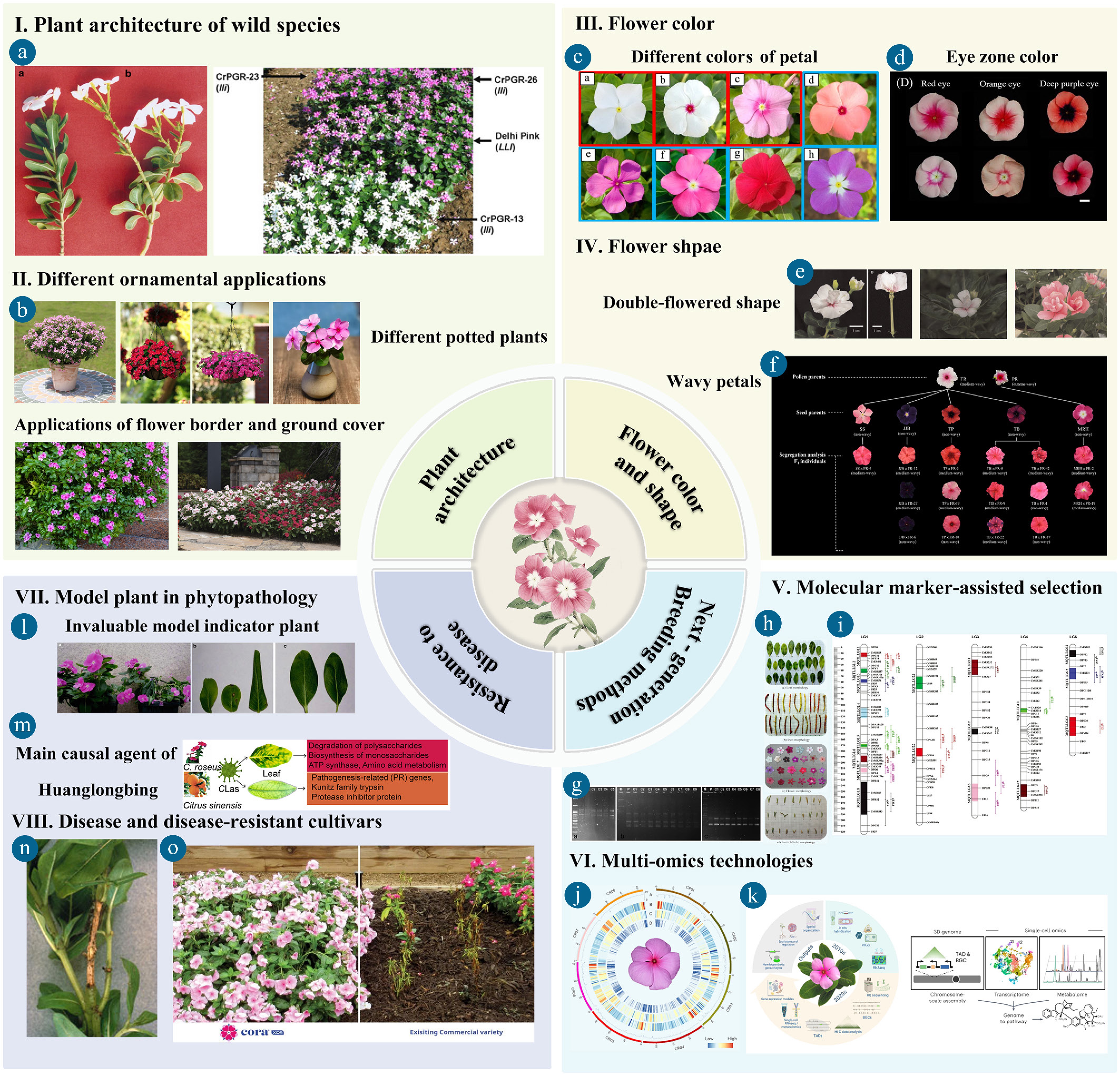
Figure 4.
Current major areas in ornamental Madagascar periwinkle research (I–V). (a) Hyper-branched leafless inflorescence of 'lli' genotype and wild type, and field view of some new ornamental genotypes of periwinkle C. roseus carrying the 'lli' mutation (adapted from Kumar et al.[92]). (b) Different ornamental applications of ornamental periwinkle, such as potted flowers, flower baskets, cutting flowers, flower border, and ground cover. (c) Different colors of ornamental periwinkle cultivars (adapted from Makki et al.[96]). (d) Different eye zone color of ornamental periwinkle cultivars (adapted from Tsai et al.[45]). (e) Double-flowering phenotype of C. roseus cv. 'Taoyuan No. 1 Rose Girl' (adapted from Chen & Yeh and Chen et al. [1,77]). (f) Top view of flowers of the pollen parents, seed parents, and F1 individuals selected for F2 generation wavy flower from segregation analysis (adapted from Huang et al.[79]). (g) Clonal fidelity of in vitro regenerated plantlets of C. roseus revealed by RAPD (TDG-CU-5), ISSR (18[GT], and 65[AG]), and ISSR (GTGTGTGT and AGAGAGAG) markers, respectively (adapted from Das et al.[97]). (h) Morphological characteristics images of 25 accessions of C. roseus. (adapted from Rani et al.[67]). (i) Chromosomal position of the detected MQTL on the C. roseus genome associated with morphological traits and pharmaceutical alkaloids with 95% confidence interval (adapted from Shariatipour et al.[98]). (j) Chromosome-level features of C. roseus (BioProject: PRJNA841429) (adapted from Sun et al.[35]) (k) The progressive improvement of the single-cell multi-omics era in C. roseus (adapted from Li et al. and Burlat et al.[34,99]). (l) Disease symptoms induced in C. roseus plants 2 months after inoculation with Xylella fastidiosa (right) (adapted from Lacava & Azevedo[100]). (m) Differences in CLas symptoms of Citrus sinensis and its agent C. roseus, and their proposed genetic basis (adapted from Xu et al.[33]). (n) Botrytis stem canker on vinca (C. roseus) (adapted from Daughtrey et al.[101]). (o) Performance assessments between disease-resistant C. roseus cv 'Cora® XDR' series against existing commercial varieties (
www.syngentaflowers.com/ams/coraxdr ).Genomics of Catharanthus roseus
-
Although MP is an important medicinal and ornamental species, its terpenoid indole alkaloid (TIA) pathway, key ornamental phenotypes, and their regulatory mechanisms remain poorly understood. Previous studies estimated the nuclear DNA content (1C-value) of C. roseus at 0.70 pg (696 Mbp) by Zonneveld et al.[31], and 0.76 pg (738 Mbp) by Guimarães et al.[18]. The rapid progress of high-throughput sequencing technologies has enabled the release of multiple high-quality C. roseus genome assemblies, allowing for detailed analysis of its biologically important traits (Table 2). Firstly, a draft genome of C. roseus assembled with short-read sequencing technology was published in 2015[32]; however, it remains highly fragmented and incomplete. Then, the application of advanced third-generation sequencing and single-cell omics technologies has enabled the assembly of near-complete, high-quality genomes of C. roseus[33−35]. These genomic resources have significantly elucidated the biosynthetic pathway of monoterpenoid indole alkaloids (MIAs). However, as an ornamental plant, the formation mechanisms of many important ornamental traits in MP, such as flower color, flower shape, and plant architecture, remain poorly understood. The acquisition of a fully assembled telomere-to-telomere (T2T) genome of C. roseus using advanced high-throughput sequencing and assembly technologies will facilitate the elucidation of its key ornamental traits. This effort will establish a critical theoretical framework for screening germplasms with both ornamental and medicinal value, and support the expansion of their industrial applications.
Table 2. Genome assemblies and annotations among sequenced Catharanthus species.
Species Assembled
genome size (Mb)Contig
N50 (Mb)Scaffold
N50 (Mb)BUSCO of
assemblyChromosome-scale Annotated
gene numberBUSCO of annotation Repeat Ref. Catharanthus roseus 541.13 0.08 2.60 96.00% No 34,363 90.70% – Kellner et al.[32] C. roseus 581.45 12.20 12.20 97.10% No – – – Cuello et al.[61] C. roseus 572.90 2.90 71.90 91.53% Yes 37,297 – – Sun et al.[35] C. roseus cv. 'Sunstorm Apricot' 572.20 11.30 71.20 98.50% Yes 26,347 96.10% 70.25% Li et al.[34] C. roseus cv. 'Pacifica XP Burgundy' 561.70 20.90 71.10 98.00% Yes 30,085 96.90% 73.71% Xu et al.[33] Interspecific hybridization and male sterility
-
The flower of MP exhibits cleistogamous floral morphology, characterized by a stigmatic head positioned inferior to the anthers[36]. This receptive structure secretes adhesive exudates that capture shed pollen. Consequently, MP demonstrates reproductive incompatibility with other species within the Catharanthus genus. Although self-compatible, effective intra-floral self-pollination is impeded by the basal localization of the stigmatic receptivity zone, necessitating alternative pollen transfer mechanisms[7]. However, natural interspecific hybridization has been documented between C. roseus and related species in Madagascar, with the majority of such crosses occurring between C. longifolius and C. roseus[37,38]. Reciprocal crossing experiments revealed asymmetric compatibility between C. roseus and C. trichophyllus. Specifically, when C. roseus was used as the female parent, fruit set failed completely, resulting in no successful introgression from C. trichophyllus into C. roseus[39].
Male sterility refers to a condition wherein functional anthers, pollen, or male gametes, fail to develop, whether through natural mechanisms or artificial induction[40,41]. Although Schnell[42] reported that MP has both self-compatible and self-incompatible lines within the genus, Veyret[37] later observed that all interspecific hybrids were self-fertile—a finding further supported by Levy et al.[43]. The absence of self-incompatibility, dioecy, or male sterility in any known Catharanthus species renders commercial hybrid seed production expensive. Subsequently, a functionally male-sterile mutant (EMS1-10) derived from the parental variety 'Nirmal' exhibits non-dehiscent anthers while retaining fertility through artificial selfing, enabling its use in hybrid seed production without the need for a restorer line[44]. The EMS1-10 mutant displays a wavy leaf margin and reduced anther size with fewer pollen grains, yet sets normal fruit and seed upon self-pollination. Field trials demonstrated effective hybrid seed yield when the male-sterile line was used as the female parent surrounded by the pollinator variety[45]. Sterility is controlled by a recessive gene linked to early-expressing morphological markers, allowing for pre-transplant identification. The mutant can be propagated via selfing, stem cuttings, or micropropagation[46]. These materials would provide valuable resources for heterosis breeding in Catharanthus.
Mutation breeding
-
Over the past three decades, mutation breeding techniques have successfully generated numerous new MP varieties with different characteristics, many of which have been commercialized. Subsequently, three phenotypically distinct reduced-height mutants, designated as 'dwarf', 'semi-dwarf', and 'bushy', have been identified in MP, exhibiting height reductions of approximately 60%, 40%, and 30%, respectively, compared to the parental cultivar 'Nirmal'[47]. The 'dwarf' and 'semi-dwarf' mutants are conferred by monogenic recessive genes, named dw1 and dw2, respectively, which are allelic. Both mutants accumulated significantly higher root alkaloid content than the parental variety[48].
The MP is also widely appreciated as an ornamental garden plant owing to its diverse floral coloration. Consequently, mutants influencing floral attributes such as color, bloom density, and longevity have attracted considerable research interest. A notable mutant of Catharanthus roseus, characterized by the development of flowers on nodes devoid of leaves and referred to as the 'leafless inflorescence' phenotype, has been reported. The genetic locus responsible for this trait, designated as 'lli', has been successfully mapped[49]. The mutant produced more flowers per plant than its parent, thus further increasing its horticultural value. Additionally, a double-flowered mutant, designated 'TYV1' ('Taoyuan No.1-Rose Girl'), was obtained from a cross between a self-pollinated C. roseus cv. 'Pacifica Polka Dot' population and a line derived from self-pollinated C. roseus cv. 'Titan Burgundy'[1]. This mutant was isolated and characterized for the morphology and genetic inheritance of its double-flowered trait. Consequently, new horticultural cultivars with enhanced ornamental characteristics were generated through hybridization of this mutant with genotypes exhibiting distinct flower colors and plant architectures.
Polyploid induction
-
In the previous studies, colchicine has become a widely used mutagenic agent for polyploidy induction in plants[50,51]. Thus, polyploid induction breeding in C. roseus, a species with significant ornamental and medicinal value, is expected to facilitate the development of elite germplasm exhibiting both enhanced ornamental traits and increased secondary metabolite accumulation.
Numerous studies on polyploidy in MP have been published in previous related research[16,52]. Seeds or apical buds of young seedlings were treated with colchicine solutions at concentrations ranging from 0.01% to 1.0% to conduct the polyploid induction of MP[27]. Furthermore, the treatment of apical buds resulted in higher tetraploid induction efficiency compared to seed treatment[53]. In addition, the stamen length and pollen grain diameter were considered as the two most reliable characters to identify autotetraploids of MP[54]. Additionally, the autotetraploids of periwinkle were also found to have low pollen fertility (32%–43%), poor fruit set (17.5%–22.5%), and low seed production as compared with diploids[16,54]. Pre-fertilization abnormalities, such as undeveloped ovules, aberrant or delayed embryo sac development, and delayed fertilization, were recognized as primary causes of seed failure (haplontic sterility) in autotetraploids of MP[55].
Polyploid induction technologies provide an excellent way to increase yields of valuable metabolites. Previous studies have indicated that autoploidization can enhance secondary metabolism in MP, thereby promoting the accumulation of terpenoid indole alkaloids[56]. Furthermore, the vincristine and vinblastine content were significantly altered and markedly increased in autotetraploids of C. roseus cv. 'Pacifica XP Red Really' and C. roseus cv. 'Pacifica XP Polka Dot', compared to their diploid counterparts[57]. This finding provides a theoretical foundation for implementing polyploid induction in cultivated varieties of C. roseus, screening elite germplasms with both ornamental and medicinal value, and further enhancing the industrial chain and added value of the species.
Next-generation breeding of Catharanthus roseus
-
Advances in next-generation sequencing (NGS) technologies over the past decade have dramatically enhanced the efficiency and cost-effectiveness of DNA sequencing, greatly accelerating genomic research[58]. Furthermore, the rapid integration of advanced technologies in recent years, such as third-generation long-read sequencing (Oxford Nanopore, Pacbio), integrated single-cell multi-omics, biosynthetic engineering, and high-throughput phenomics, has propelled crop breeding into a new era of intelligence-driven improvement[59].
As an important medicinal and ornamental species, C. roseus has encountered significant challenges in the next-generation molecular breeding stage due to a lack of high-throughput genomic data. In 2006, Murata et al.[60] built cDNA libraries from the leaf base and root tips of C. roseus and generated expressed sequence tags (ESTs). This work provided the initial platform for identifying microsatellite markers, thereby paving the way for molecular breeding efforts in this species. Although the first whole-genome assembly of C. roseus cv. 'Sunstorm Apricot' has been made publicly available; however, the current version remains fragmented, with 68,690 gaps that present opportunities for further refinement[32,61].
The MAS is an advanced breeding approach that integrates molecular biological techniques with conventional plant breeding practices[62]. The MAS strategy allows breeders to precisely and efficiently identify plants carrying target genes or quantitative trait loci (QTLs) during early developmental phases, thus streamlining cultivar development and enhancing the overall effectiveness of breeding programs. The initial genetic linkage map for C. roseus was constructed by Gupta et al.[63], which integrated six morphological markers and 125 molecular markers (79 RAPD,7 ISSR,2 EST-SSR,37 other PCR-based DNA markers) into 14 linkage groups covering a total length of 1,131.9 cM. Subsequent linkage maps were established by Shokeen et al.[64], featuring 114 markers distributed across eight linkage groups (632.7 cM), and by Chaudhary et al.[49]—later expanded by Sharma et al.[65]—incorporating both morphological and molecular markers. These foundational maps facilitated the detection of QTLs associated with alkaloid production in various plant tissues. Further advancing this work, Chaudhary et al.[66] identified 20 QTLs linked to the accumulation of key alkaloids such as catharanthine and vindoline, underscoring the potential for marker-assisted selection in MP breeding. It should be noted, however, that the expression and effects of these QTLs may vary across different genetic backgrounds and growing conditions, highlighting the need for further validation. The relatively slow adoption of marker-assisted breeding in C. roseus likely reflects its secondary economic status compared to major global crops.
Nevertheless, the development of molecular markers linked to key ornamental traits in MP remains limited[67]. By employing genetic markers tightly linked to agronomically important traits, MAS enables precise identification and selection of individuals possessing desirable alleles, thus significantly enhancing the efficiency of developing improved cultivars[68]. Utilizing the recently established high-quality reference genome to identify markers associated with traits such as flower color, floral morphology, and plant architecture could greatly enhance the efficiency of marker-assisted breeding in MP. This strategy promises to facilitate broader application within the ornamental breeding sector.
-
Molecular breeding techniques have facilitated the successful transfer of genes in numerous ornamental species, resulting in the development of transgenic plants with targeted traits. Progress in molecular biology has transcended historical cross-species ability limitations, enabling gene exchange among previously incompatible species. Recognized for its precision and efficiency, molecular breeding offers a superior alternative to conventional breeding and induced mutagenesis. It allows specific traits to be altered in a highly directed manner while preserving the integrity of other desirable characteristics. Ornamental qualities such as flower color and morphology, plant architecture, postharvest durability, disease resistance, control of flowering, and tolerance to abiotic stress, represent viable targets for genetic enhancement through this approach, serving as key focal points for modern ornamental breeding programs (Fig. 5).
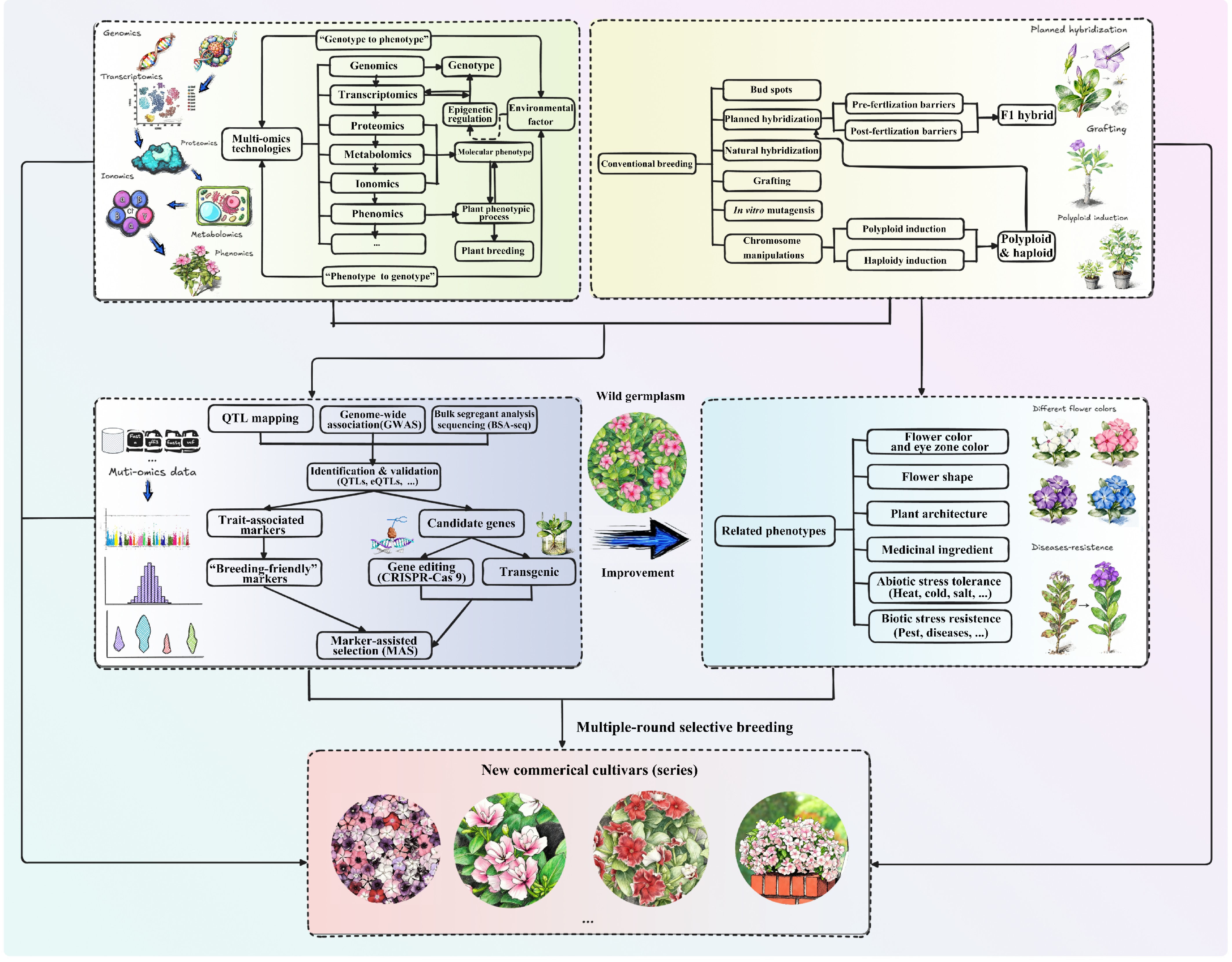
Figure 5.
The hypothetical comprehensive breeding scheme for ornamental periwinkle integrating conventional with modern breeding strategies. Icons were created using Excalidraw (
https://excalidraw.com ), and AI-Doubao (www.doubao.com/chat ).Genetics of flower color
-
Developing novel and appealing flower colors remains a key objective in ornamental plant breeding, driven by its profound influence on consumer choices and commercial value[69]. Flower (corolla) color represents one of the most visually prominent and highly variable traits in MP populations. The horticultural significance of MP largely stems from its diverse and varied floral coloration, which provides valuable resources for ornamental breeding. In natural populations of MP, three predominant corolla color patterns are typically observed: pink, white, and a distinct red-eyed form characterized by a white corolla with a prominent red central eye[70]. Beyond the common corolla variants, less frequent hues, including violet, orange-red, scarlet-red, magenta, and rose, have also been reported. The full spectrum of observed colors (spanning pink, red-eyed, pale pink, white, violet, orange-red, scarlet-red, magenta, and rose) is regulated by interactions among alleles across seven genetic loci: A, R, W, B, I, O/Om, and J[71,72]. Initially, a phenotypic segregation ratio of nine pink:three red-eyed:four white was observed in a cross between pink- and white-flowered plants[73]. Genotypically, the pink phenotype was associated with R-W-, the red-eyed phenotype with R-ww, and the white phenotype with both rrW- and rrww. The I allele also promotes anthocyanin accumulation, though at reduced levels, and in the absence of the W allele, it produces pale pink corollas with a red eye[72]. Gene B acts as a co-pigmentation factor that induces a spectral blue shift in anthocyanin pigments. Additionally, genes A and R act as complementary genes essential for corolla pigmentation; the absence of either gene results in the formation of a white corolla[74]. For another inhibitory gene J, while non-pigmenting on its own, partially suppresses gene Om, resulting in the formation of rose-colored corollas[70]. Moreover, recent studies have demonstrated that the F1 progeny from a cross between the inbred lines 'Vi-15-1' (which has magenta corollas with white centers) and 'Vi-29' (white-flowered) consistently produced pale purple corollas. Additionally, segregation analysis of the F2 generation identified three distinct phenotypic classes aligned with a typical 1:2:1 monogenic ratio: (i) magenta with white center (OmOm); (ii) pale purple (OmW); and (iii) white (WW)[5]. Similar results were found by Awad Hamza et al.[21], where crosses between pink- and white-flowered parents in periwinkle yielded light pink F1 progeny, with the F2 generation segregating in a 1:2:1 ratio, indicating monogenic inheritance with incomplete dominance of the pink flower color allele.
Although the inheritance patterns governing flower color in MP have been elucidated, the molecular mechanisms underlying petal pigmentation remain inadequately characterized. A recent multi-omics study on three C. roseus varieties with white (CW), red (CR), and purple (CP) petals identified 10 anthocyanins as key color determinants[75]. The petals of CP were characterized by malvidin 3-O-glucoside and petunidin 3-O-glucoside, while petals of CR accumulated peonidin 3-O-glucoside and delphinidin 3-O-glucoside. Transcriptome analysis revealed 4173 differentially expressed genes (DEGs), including 1,003 genes commonly regulated across samples, with notable upregulation of chalcone synthase (CHS), flavonoid 3'-hydroxylase (F3'H), and dihydroflavonol 4-reductase (DFR) in colored petals. Furthermore, the expression patterns of MYB and bHLH transcription factors exhibited significant correlations with observed color phenotypes. The competitive enzymatic interaction between flavonol synthase (FLS) and flavonoid 3'-hydroxylase/flavonoid 3',5'-hydroxylase (F3'H/F3'5'H) for the substrate dihydrokaempferol (DHK) likely impedes anthocyanin biosynthesis in the petals of CW. Collectively, these findings elucidate the molecular framework of petal coloration in periwinkle and identify potential genetic targets for the development of pigmented varieties. Therefore, the combined application of multi-omics approaches, genome editing techniques, and traditional breeding methods holds significant potential to enhance the comprehension of corolla color development, elucidate the biochemical mechanisms underlying floral pigmentation, and enable the creation of new flower color phenotypes of MP.
Flower shape
-
The growth and development of angiosperms (flowering plants), particularly of their floral organs, are fundamental characteristics[76]. Similar to numerous species within the Apocynaceae family, the flower of MP exhibits a five-lobed, planar corolla that emerges from a tubular structure enclosing the stamens and pistil[36]. Commercial MP cultivars currently exhibit limited variation in flower morphology, as most feature a common flat-petaled structure that reduces their novelty. In 2001, a double-flowered mutant designated 'Taoyuan No.1-Rose Girl (TYV1)' emerged from self-pollinated (S1) lines of the C. roseus cv. 'Pacifica Polka Dot'[1]. Subsequently, this mutant provided the genetic foundation for breeding the commercial doubled-flower cultivar[77]. Owing to its exceptional ornamental value, the double-flowered trait has received a great deal of attention from breeders in the MP industry. Many horticultural companies, such as Suntory Flower Co. (Japan), have introduced specialty MP varieties like the 'Soirée® Kawaii' series, featuring miniature flowers, and the 'Soirée® Flamenco' series, known for its wavy petals[20]. However, studies on the description or inheritance pattern of the wavy flower and miniature flower form in MP remain scarce.
Wavy petals are common in many ornamental flowers due to their enhanced aesthetic appeal, making them a frequent target for breeders[78]. Neither resulting from the typical homeotic conversion of stamens into petals nor following a dominant inheritance pattern, the 'TYV1' mutation was identified as being governed by a single recessive allele. Recent research indicates that the inheritance of these flower morphologies is governed by a single allele, WAVY (Wv), proposed to exhibit incomplete dominance[79]. Specifically, the non-wavy, extreme-wavy, and medium-wavy phenotypes result from the recessive homozygous (wvwv), dominant homozygous (WvWv), and heterozygous (Wvwv) genotypes, respectively, enabling breeders to predict offspring flower-type ratios based on parental genotypes. Furthermore, notable characteristics such as broadly wrinkled leaves, an acute leaf apex, and a loose bud structure allows for early identification and selection of extreme-wavy MP flowers during breeding. Consequently, these results would provide a foundational framework for breeders to design crossing strategies aimed at generating a broader range of flower forms, thereby enhancing the diversity available to consumers.
However, the molecular regulatory mechanisms behind the diverse floral morphologies (such as wavy and double petals) in MP remain poorly understood. In contrast, studies in the model ornamental plant petunia have shown that different MADS-box transcription factors and TCP transcription factors mainly control the development of its double and wavy petals[80−82]. Therefore, discovering and functionally validating homologous genes within this transcription factor family in periwinkle will offer important insights into the molecular basis of its floral morphology development, while also helping to clarify the conservation and specificity of floral patterning pathways across species.
Plant architecture
-
Plant architecture, defined here as the three-dimensional organization of the shoot system in higher plants, is primarily shaped by shoot branching patterns, plant height, and inflorescence morphology[83]. In particular, plant height plays a fundamental role in plant architecture, collaborating with shoot branching and inflorescence morphology to define the overall structure[84]. As a dominant ornamental crop widely used in landscaping for borders, bedding, and pot culture, the MP remains poorly understood genetically, particularly regarding the inheritance of key traits and their correlation with quantitative characters such as flower yield, yet elucidating the co-segregation of these traits could greatly facilitate crop improvement.
An extremely tall mutant (EMS 18-12), a bushy mutant (EMS 24-5), their parental variety 'Nirmal', and a double mutant recombinant derived from these materials were used to examine the individual and combined effects of genes that exert opposing influences on plant height in MP[48]. The results demonstrated that plant height in the extremely tall mutant (EMS 18-12) is regulated by an epistatic inhibitory interaction between two independently inherited dominant genes, Et and H, where Et promotes extreme height while H suppresses its expression. Moreover, both genes were observed to segregate independently of the plant height-reducing recessive gene present in the bushy mutant (EMS 24-5). At various developmental stages, the double mutant recombinant (Etby) exhibited greater height than the bushy mutant (EMS 24-5) and the variety 'Nirmal', but remained shorter than the extremely tall mutant (EMS 18-12). Recently, a study was performed using 30 F2 segregating lines of Catharanthus developed from diallel crossing of six genetically dissimilar parents varied in many traits. Phenotyping of the population results indicated a significant positive association between days to flowering and plant height. Although major horticultural companies have introduced varieties with diverse plant architectures, such as 'Solar' (Hem Genetics, Netherlands) and 'Cora® Cascade XDR' series, which are suitable for hanging baskets, the molecular mechanisms controlling their growth habits are still not fully understood. Therefore, using multi-omics approaches to uncover the molecular regulatory mechanisms behind different plant architectures in ornamental MP will help develop more precise selection and hybrid breeding strategies in the future.
Although these traits follow monogenic recessive inheritance, their underlying molecular determinants remain uncloned. Research in other ornamentals provides strong mechanistic clues: dwarf phenotypes often implicate gibberellin biosynthesis, or signaling genes (e.g., GA20ox, GA3ox in chrysanthemum)[85], and bushy phenotypes frequently correspond to strigolactone signaling deficiencies (e.g., in MAX2 orthologs)[86]. Consequently, a critical next step for MP architecture research is hypothesis-driven functional validation of candidate genes within these conserved pathways.
Resistance to disease
-
Periwinkle plants are known to be susceptible to phytoplasma infection from different crops, and exhibit highly distinctive symptoms such as root necrosis, water-soaked lesions on stem tissue, and uniform wilting. Especially for die-back disease, induced by Pythium aphanidermatum or Phytophthora nicotianae, severely affects tropical and subtropical regions during the rainy season, with documented mortality rates reaching 70%–80%[27,87]. The Phytophthora aerial blight in annual MP advances quickly under environmental conditions such as high soil moisture, prolonged rainfall, or frequent irrigation[88]. The MP wilting and death may occur as quickly as 1–2 weeks after initial symptom emergence; however, the root systems of infected plants often remain intact[89]. Notably, a die-back-resistant variety, C. roseus cv. 'Nirmal' was developed through pure line selection from an individual plant that survived a severe die-back epidemic[90].
The genetic basis of die-back resistance was investigated employing a resistant dwarf mutant originating from the green-stemmed cultivar 'Nirmal', in comparison with a susceptible purple-stemmed accession designated 'OR'[91]. Furthermore, the qualitative analyses consistently demonstrated that resistance is governed by a single gene, exhibiting a broad-sense heritability of 0.85, and is inherited independently of loci controlling dwarfism and stem pigmentation. The over-flowering 'lli' trait was combined with variation in plant height, petal and eye colors, and tolerance to the common fungal diseases[92]. Consequently, a growing number of new cultivars with superior ornamental traits, including enhanced tolerance to common fungal diseases, heat, and drought, have been widely commercialized. Notable examples include the 'Cora® Cascade XDR' series.
-
An integrated analysis of current commercial varieties (Table 1), and relevant literature reveals several distinct trends in the current commercial breeding of Madagascar periwinkle. First, innovations in plant architecture have significantly expanded its horticultural applications. Breeders have successfully developed series with trailing habits (e.g., 'Cora® Cascade' series), transforming the plant from a traditional bedding flower into a premium material suitable for hanging baskets and container cultivation. Second, the continuous diversification of floral aesthetic traits represents another major driver, including double and mini flowers (e.g., the 'Soirée® Double' and 'Soirée® Kawaii' series), wavy petals (e.g., the 'Soirée®Flamenco' series), and unique eye patterns and complex color formations, all of which markedly enhance its ornamental value and market appeal. Furthermore, disease resistance has become a core breeding objective, particularly for open landscape applications. For example, Syngenta's 'Cora® XDR' series aims to address key production constraints such as die-back caused by pathogens like Phytophthora. Driven by market globalization and the proliferation of e-commerce platforms, these breeding trends underscore the genetic gains achieved through market forces. Leading commercial series efficiently integrate multiple desirable traits to meet the demands of the modern horticulture industry.
However, this intensive commercial breeding exerts a dual influence on the genetic diversity of Catharanthus roseus. While reliance on elite parental lines, risks genetic erosion and a narrowed genetic base, modern techniques such as interspecific hybridization and gene editing, create novel combinations. Thus, future strategies must balance the use of advanced technologies with the conservation of traditional germplasms to ensure long-term breeding sustainability.
-
Although multi-omics approaches have profoundly advanced the study of medicinal alkaloid biosynthesis in Catharanthus roseus, the molecular basis of its ornamental traits remains largely unexplored[34,35]. Key aesthetic characteristics of periwinkle, including corolla pigmentation, petal morphology (such as minuteness, waviness, and doubleness), and plant architecture, are still poorly defined genetically. The integration of CRISPR-mediated gene editing with multi-omics profiling provides a powerful strategy to systematically unravel these traits[93,94]. Notably, the recent application of an intron-optimized Cas9 (zCas9i) system in C. roseus has enabled highly efficient multiplex editing of jasmonate-responsive transcription factors (JAM2, JAM3, RMT1), achieving 100% editing efficiency via Agrobacterium-mediated hairy root transformation[95]. This breakthrough not only validates the cross-species applicability of the intron-optimized Cas9 platform but also provides a critical toolset for targeting key genetic regulators in future functional studies, such as MADS-box genes for flower form, MYB factors for pigment patterning, and susceptibility genes like MLO for stem rot resistance.
As an important bedding plant, the MP maintains a stable presence in the global ornamental market, with increasing adoption in tropical and subtropical regions owing to its extended bloom period and thermal resilience[75]. Its widespread commercial presence is evidenced by the extensive variety series developed by major international breeders such as Syngenta®Flowers, PanAmerican Seed, and Suntory Flowers.
Future commercial growth will largely hinge on the introduction of novel cultivars with improved visual and performance-related traits. In parallel, enhancing resistance to major diseases such as die-back and stem rot through integrated strategies, combining biological controls and genome-edited resistance, will be crucial for sustainable production.
In summary, the overarching goal for next-generation periwinkle breeding lies in the holistic design of elite cultivars that harmonize ornamental appeal, durable disease resistance, and high alkaloid content, thereby maximizing the dual horticultural and pharmaceutical potential of this multifaceted species.
This work was supported by the Foundation for Cultivated Young Talents of Fujian Province, China (Grant No. 2025350417), and the High-Quality Agricultural Development '5511' Collaborative Innovation Project Special Topic of Fujian Provincial People's Government & Chinese Academy of Agricultural Sciences (Grant No. XTCXGC2021003).
-
The authors confirm contribution to the paper as follows: conception and supervision: Zhong H; data collection: He J, Lai N; image collection and processing: He J, Liu S, Zhong S; manuscript suggestions: Liu S, Chen Y, Luo Y, Fan R, Ye X, Wu J, Zhan R; draft manuscript preparation: He J; manuscript reviewing and editing: He J, Zhong H. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are available from the author on reasonable request.
-
The authors declare that they have no conflict of interest. Siyang Liu and Ruiqi Zhang are affiliated with Tangchao Horticulture Technology (Xiamen) Co. Ltd, which contributed to image collection and manuscript suggestions. The company had no commercial or financial interest related to the content of this review.
- Supplementary Table S1 Publicly accessible websites for downloading the images shown in the Figure 3.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
He J, Liu S, Zhong S, Chen Y, Luo Y, et al. 2026. Germplasm resource and genetic breeding of ornamental Madagascar periwinkle (Catharanthus roseus): a brief review. Ornamental Plant Research 6: e003 doi: 10.48130/opr-0025-0049 

Germplasm resource and genetic breeding of ornamental Madagascar periwinkle (Catharanthus roseus): a brief review
- Received: 28 September 2025
- Revised: 04 November 2025
- Accepted: 19 November 2025
- Published online: 27 January 2026
Abstract: Madagascar periwinkle (Catharanthus roseus [L.] G. Don) is a valuable medicinal and ornamental plant, prized for its diverse alkaloid compounds and aesthetic traits. While extensively studied for pharmaceutical applications, its current understanding of preliminary genetic inheritance of ornamental traits and ornamental breeding research are less advanced compared to other horticultural crops. This review systematically examines the germplasm resources, genetic diversity, and breeding advancements in ornamental periwinkle, covering the utilization of wild and cultivated varieties, mutation and polyploidy breeding, and recent genomic sequencing efforts. The potentially genetic mechanisms underlying key ornamental characteristics such as flower color, flower type, and plant architecture are discussed, and emerging approaches evaluated, including marker-assisted selection, interspecific hybridization, and CRISPR-based editing integrated with multi-omics platforms, for trait decoding and precision breeding. Strategies are also highlighted to enhance resistance to prevalent diseases such as die-back and root rot. Finally, future research directions aimed at developing resilient, high-alkaloid cultivars suitable for sustainable ornamental horticulture are proposed. This work provides a comprehensive resource for accelerating molecular breeding and improving ornamental traits in C. roseus.
-
Key words:
- Ornamental breeding /
- Catharanthus roseus /
- Germplasm resources /
- Molecular breeding