









-
Potato (Solanum tuberosum) is the fourth largest food crop after corn (Zea mays), wheat (Triticum aestivum) and rice (Oryza sativa) in the world[1]. With the acceleration of modern life, fresh-cut potatoes have gained growing popularity in people's daily lives[2]. However, fresh-cut potatoes have several problems after cutting, such as softening, an unpleasant aroma and browning[3]. Among these problems, browning is the most common deterioration in the appearance of fresh-cut potatoes. As browning is enzymatic, it severely reduces their shelf life. When oxygen and enzymes are present, the phenolic compounds produce quinones, which then transform into pigments[4]. During the process, polyphenol oxidase (PPO) is the vital enzyme, which catalyzes not only the hydroxylation of monophenols to diphenols but also the oxidation of diphenols to quinones[1]. In recent years, many biological, chemical and physical technologies have been reported that can repress the browning of fresh-cut potatoes. Chemical technologies can significantly alleviate the browning of fresh-cut potatoes, including sulfur dioxide and sulfites, but these have food safety issues[5]. Although physical methods can effectively repress the browning of fresh-cut potatoes, these also have some issues. For example, low oxygen could markedly alleviate the browning of fresh-cut potatoes, but it leads to off-flavors and unpleasant odors[6]. Biological technologies (some natural extracts, natural edible coatings and genetic engineering technologies) can effectively inhibit the browning of fresh-cut potatoes, but most biological technologies cannot be used commercially[7]. Previous studies found that some amino acids can repress the browning of fresh-cut potatoes, including isoleucine and L-cysteine, arginine, etc.[2,8,9]. Moreover, our previous work reported that glutamic acid (Glu, C5H9NO4) treatment markedly alleviates the browning of fresh-cut potatoes[5], but its regulatory mechanism was unclear.
According to the DNA-binding domains, transcription factors (TFs) can be classified into ethylene response factors (ERFs), WRKY transcription factors (WRKYs), NAC transcription factors (NACs), basic leucine zipper (bZIP) proteins, and so on[10]. Among them, bZIPs are named for the conserved bZIP domain, which possesses a leucine zipper dimerization domain and a basic DNA-binding domain[11]. Several studies have reported that bZIPs mediate fruit development, maturity and resistance to biotic and abiotic via directly interacting with its downstream genes[12]. For example, in apple (Malus) 'Pinkspire' fruit, MpbZIP9 enhances anthocyanin biosynthesis by activating MpF3'H expression[13]. In citrus, CsbZIP44 promotes carotenoid biosynthesis via directly activating the expression of CsNCED2, CsBCH1, CsGGPPs and CsDXR[14]. In strawberry (Fragaria × ananassa), FaTRAB1 (a bZIP transcription factor) enhances anthocyanin biosynthesis by repressing FaMYB10 expression, which directly stimulates the expression of FaOMT, FaUFGT, FaF3'H and FaANS[15]. In pear (Pyrus) , PpbZIP44 promotes carbohydrate metabolism and the accumulation of flavonoids and amino acids by directly activating the transcriptional activity of PpProDH1 and PpSDH9[16]. PbrbZIP15 improves the accumulation of fruit sugars by stimulating expression of the glucose isomerase gene PbrXylA1[17]. In banana (Musa acuminata), MabZIP93 overexpression promotes fruit ripening by inducing the expression of MaXTH23, MaXGT1, MaPE1 and MaPL2[11]. Moreover, MabZIP24 promotes fruit ripening by stimulating the expression of MaACO1, MaEXPA15, MaPE42, MaPE51, MaPL5, MaPL15 and MaPG3[18]. In sweet cherry (Prunum avium), PavbZIP6 enhanced fruit anthocyanin accumulation via positively regulating the expression of anthocyanin biosynthesis genes (PavUFGT, PavANS and PavDFR)[19]. Moreover, bZIP proteins also regulate fruit browning. PpbZIP23 and PpbZIP25 mediate the browning of peach (Prunus persica) fruit caused by chilling injury[12]. However, the regulatory mechanism of bZIP TFs regulate the browning of fresh-cut fruit and vegetables was still unknown.
ERFs belong to the AP2 ERF family, which is a main regulator in fruit development, maturity and ripening[20]. In tomato (Solanum lycopersicum), SlERF.F12 negatively mediates fruit ripening by repressing the expression of SlACO2 and SlACS4[21]. SlERF.D3 negatively regulates fruit ripening and leaf senescence[22]. SlERF.D6 overexpression promotes steroidal glycoalkaloids synthesis and fruit ripening[23]. SlERF.D7 promotes fruit ripening by activating SlARF2 expression[24]. SlERF.F12 represses the ripening of tomato fruit by repressing the expression of SlACS2 and SlACS4[21]. SlERF.C1 enhances fruits' resistance to Botrytis cinerea via stimulating the expression of pathogenesis-related (PR) genes[25]. In sweet cherry, PavRAV2 negatively regulates fruit size by directly repressing PavKLUH transcription[26]. In apple, MdERF34 can activate anthocyanin biosynthesis in the fruits[27]. MdERF4 inhibits fruit ripening via suppressing ethylene synthesis[28]. MdERF113-overexpression enhances fruits' cold tolerance and drought resistance[29]. In banana, MaERF95L positively mediates fruit ripening by stimulating the expression of sucrose synthesis and starch degradation genes[30]. MaERF113 promotes fruit ripening by stimulating the expression of cell, starch and chlorophyll degradation genes[31]. MaERF003 mediates fruit ripening via directly inducing the expression of chlorophyll, cell wall and starch degradation genes[32]. In peach, PpERF5 and PpERF7 increase peach fruit aroma via positively regulating PpLOX4 expression[33]. Moreover, ERFs may mediate the browning of vegetables. CaERF4-2, CaAP2L1 and CaERF32/41/43/80/91/114 mediate the browning of sweet pepper (Capsicum anuum)[34]. However, the regulatory mechanism of ERFs in regulating the browning of fresh-cut fruit and vegetables is still unclear.
Herein, we found that the expression of StbZIP53-like2 was closely associated with the browning of fresh-cut potatoes, and its expression was enhanced by Glu treatment. StbZIP53-like2 markedly repressed the expression of StPPO2. StbZIP53-like2 interacts with StERF091, the expression of which was increased by Glu treatment. StERF091 severely repressed the transcription of StPPO2 and StPPO3. Moreover, the interaction of StERF091 and StbZIP53-like2 enhanced the transcriptional activity of StPPO2 and StPPO3. Our study revealed that StbZIP53-like2–StERF091 alleviates the browning of fresh-cut potatoes by inhibiting the expression of StPPO2 and StPPO3.
-
Potatoes were treated via the method of Guan et al.[3]. In brief, potatoes were shredded and rinsed in a 5 × 10−5 g/L NaClO solution for 15 s, followed by draining with sterile gauze at room temperature. The shredded potatoes were then divided into two groups. The control group was immersed in deionized water for 4 min at room temperature, whereas the Glu group was immersed in a 15 g/L Glu solution for 4 min under the same conditions. Subsequently, all samples were stored at 4 °C for subsequent analysis. Each group consisted of three biological replicates. Samples were collected at 0 (before treatment), 12, 24, 48 and 72 h and stored at −80 °C.
Ethylene production
-
For measuring ethylene production, 500 g of fresh-cut potatoes from the control and Glu groups were separately placed in an airtight container equipped with a rubber stopper for 2 h at 25 °C. Subsequently, 1 mL of the headspace gas was collected from each sample in triplicate for determining ethylene. Ethylene production was measured by the method of Zhu et al.[35].
Transcription level analysis
-
Reverse transcription–quantitative polymerase chain reaction (RT-qPCR) was performed, following the protocol of Guan et al.[3]. Briefly, total RNA was extracted from different fresh-cut potatoes using the FastPure Universal Plant Total RNA Isolation Kit and reverse-transcribed into cDNA with the EVO M-MLV Kit. Gene transcription levels were calculated by the 2−ΔΔCᴛ method[36], using β-actin as the internal reference gene[3].
Sequencing and gene annotation
-
RNA-Seq analysis was carried out according to Guan et al.[3]. Briefly, fresh-cut potatoes treated with or without Glu at 0 (before treatment), 12 and 48 h were analyzed by RNA-Seq with three biological replicates per time point. All cDNA libraries were sequenced and analyzed by OE Biotechnology Co., Ltd. (Shanghai, China) on an Illumina Hi-Seq 2500 platform.
Prior to downstream analysis, clean reads from each sample were mapped to the potato reference genome database (
www.ncbi.nlm.nih.gov/datasets/genome/?taxon=4113 ) using TopHat2 software. A heatmap was generated with Tbtools, and differentially expressed genes (DEGs) were identified via clustered profile analysis with thresholds of p ≤ 0.05 and a fold-change of ≥ 2.Subcellular localization assay
-
The full coding sequences (CDSs) of StbZIP53-like2 and StERF091 were cloned into the pBE-GFP vector to obtain StbZIP53-like2-GFP and StERF091-GFP vectors. Then the StbZIP53-like2-GFP, StERF091-GFP and pBE-GFP vectors were individually expressed in tobacco (Nicotiana benthamina) leaves by Agrobacterium tumefaciens mediation. After 36–48 h, the green fluorescence protein (GFP) signals in the tobacco leaves were detected using a fluorescence microscope (Zeiss Axioskop 2 Plus, Leica, Solms, Germany) at 488 nm[3].
Yeast two-hybrid assay
-
The CDSs of StbZIP53-like2 and StERF091 were separately cloned into pGBKT7 and pGADT7 to construct the AD-StbZIP53-like2, AD-StERF091, BD-StbZIP53-like2 and BD-StERF091 vectors. Sequentially, AD-StbZIP53-like2 + BD-StERF091 and BD-StbZIP53-like2 + AD-StERF091 were expressed in yeast strains to confirm their interaction. The transformed yeast strains were grown at 28 °C on a culture medium without adenine (Ade), histidine (His), tryptophan (Trp) and leucine (Leu), and the interaction of StbZIP53-like2 and StERF091 were tested according to their growth status and α-galactosidase activity. Recombinants of T-antigen + lamin and T-antigen + P53, respectively, served as the negative and positive controls.
GST pull-down assay
-
The CDS of StbZIP53-like2 was cloned into the pGEX-4T-1 vector to get GST-StbZIP53-like2 recombinant proteins, which were induced with 0.1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) at 16 °C for 16 h. Moreover, the CDS of StERF091 was cloned into the PET-28a vector to get His-StERF091 recombinant proteins, which were induced with 0.2 mM IPTG at 28 °C for 6 h. GST-StbZIP53-like2 and His-StERF091 proteins were purified according to Song et al.[33]. His-StERF091 was then incubated with gutathione S-transferase (GST) or GST-StbZIP53-like2, and their interaction was detected by Western blotting using anti-His antibodies and anti-GST antibodies (Huaan, Shenzhen, China).
Co-immunoprecipitation assay
-
The CDS of StERF091 was inserted into the pEAQ-HT-GFP vector, and the CDS of StbZIP53-like2 was ligated to the PBI121-Flag vector. The recombinant vectors were then infiltrated into 6-week-old tobacco leaves using A. tumefaciens GV3101. After 48 h of infiltration, the protein was extracted from tobacco leaves and reacted with the anti-GFP rabbit antibody (Cat. No. ab290, Abcam, Cambridge, UK) for 2 h. Then the protein A agarose beads were mixed and incubated for 4 h and washed four times before the the beads were collected for Western blot analysis using anti-Flag (Cat. No. 3165, Sigma) and anti-GFP (Cat. No. ab290, Abcam).
Transient dual luciferase expression assay
-
The CDSs of StbZIP53-like2 and StERF091 were separately cloned into the pGreenII62-SK vector to serve as effector constructs, whereas the promoter regions of StPPO2, StPPO3 and StPPO7 were cloned separately into the pGreenII-0800-LUC vector as reporter plasmids. Subsequently, the reporter and effector vectors were co-expressed in tobacco leaf tissues by A. tumefaciens mediation. Following 48 h of expression, luciferase activity was determined in accordance with the protocol provided by the Dual Luciferase Assay Kit (Promega, Madison, WI, USA). Each dual luciferase reporter assay was conducted with six replicates, and the resulting data are presented as the firefly luciferase/renilla luciferase (LUC/REN) ratio.
Yeast one-hybrid library assay
-
AD-StbZIP53-like2 and AD-StERF091 were obtained as described above. The promoter sequences of StPPO2 and StPPO3 were separately cloned into the pBait-ABAi vector to generate bait plasmids, which were subsequently transduced into the yeast one-hybrid (Y1H) Gold strain. To verify the binding affinity, the AD-StbZIP53-like2 and AD-StERF091 vectors were separately introduced into the bait–reporter yeast strain, and their interaction was validated. Protein–DNA interactions were assessed on the basis of the growth status of co-transformed yeast cells on SD/-Leu medium supplemented with Aureobasidin A (AbA).
Electrophoretic mobility shift assay
-
The CDS of StERF091 was inserted into the pGEX-4T-1 vector to get GST-StERF091 recombinant proteins, which were induced with 0.1 mM IPTG at 16 °C for 16 h. The recombinant protein was purified according to Guan et al.[3]. The GST-StbZIP53-like2 protein was obtained as described above. Probes of StPPO2 and StPPO3 were labeled at the 3' end via the method of Guan et al.[3]. Unlabeled wild-type promoter fragments containing the GCC-box or C-box were used as competitors, whereas mutant probes with the GCC-box or C-box replaced by AAAAAA served as negative controls. The electrophoretic mobility shift assay (EMSA) was conducted according to Guan et al.[3].
Statistical analysis
-
All experiments were performed with three or six independent biological replicates. Data were analyzed with Duncan's multiple range test using SPSS 25 or the t-test, and the results are presented as the mean ± standard deviation (SD). All primers are listed in Supplementary Table S1.
-
As shown in Fig. 1a, ethylene content increased first and then decreased, and Glu treatment significantly induced its content. Eleven bZIP genes with differences in expression were identified by our RNA-Seq analysis. The expression of StbZIP17, StbZIP17-like, StbZIP53-like1 and StbZIP60 was reduced with incrased storage time, and the expression of StbZIP53, StbZIP53-like, StbZIP53-like2, StbZIP53-like3, StbZIP53-like5 and StbZIP53-like6 was increased with increased storage time. The expression of StbZIP17, StbZIP17-like, StbZIP53, StbZIP53-like, StbZIP53-like3, StbZIP53-like4 and StbZIP60 was inhibited by Glu treatment at 12 h but was enhanced by Glu treatment at 24 h. However, the expression of StbZIP53-like6 was induced by Glu treatment at 12 h but was repressed by Glu treatment at 24 h. Moreover, only the expression of StbZIP53-like1 and StbZIP53-like2 was induced by Glu treatment at both 12 h and 24 h, and the expression level of StbZIP53-like2 was the highest (Fig. 1b). Thus, it was selected for further study. RT-qPCR analysis confirmed this result, showing that the expression first increased and then decreased, and that Glu treatment markedly induced its expression (Fig. 1c). Moreover, StbZIP53-like2 contains 166 amino acids and a bZIP_plant_GBF1 domain (Supplementary Fig. S1) and is located in the nucleus (Fig. 1d).
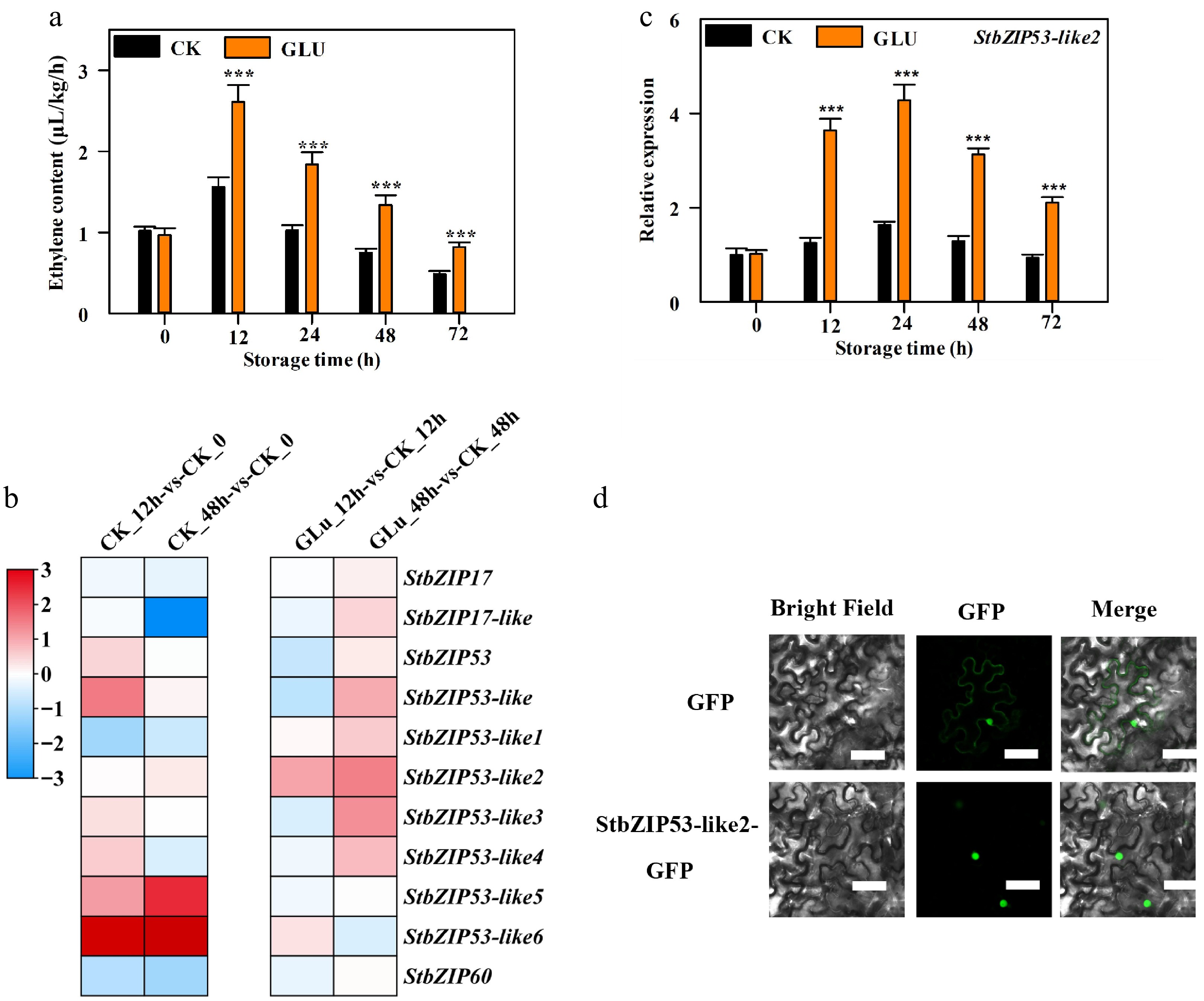
Figure 1.
Ethylene content and the expression and subcellular location of StbZIP53-like2. (a) Ethylene content. (b) Expression profiles of 11 StbZIP genes in the RNA-Seq analysis during storage. (c) The expression of StbZIP53-like2. The expression levels of StbZIP53-like2 at different points are relative to 0 h, which was set as 1. *** indicates a significant difference at p < 0.001. (d) The subcellular location of StbZIP53-like2. Bars = 50 μm.
StbZIP53-like2 mediates the expression of StPPO3
-
Our previous study found that the transcription of StPPO2, StPPO3 and StPPO7 was closely related to the browning of fresh-cut potatoes[3]. Therefore, we aimed to find out whether the expression of StPPO2, StPPO3 and StPPO7 was regulated by StbZIP53-like2. A DLR assay was performed; the promoters of StPPO2, StPPO3 and StPPO7 were separately cloned into pGreenII 0800-LUC effector vector; and the CDS of StbZIP53-like2 was cloned into the pGreenII 62-SK reporter vector (Fig. 2a). The promoter activity of StPPO3 was severely repressed by StbZIP53-like2, but the promoters of StPPO2 and StPPO7 were not regulated by StbZIP53-like2 (Fig. 2b). EMSA was used to verify StbZIP53-like2 binding of the promoter of StPPO3. The data indicated that GST-StbZIP53-like2 could directly interact with the C-box motif of StPPO3 and caused mobility shifts, but GST protein alone did not cause a mobility shift. Moreover, the shifted bands were reduced when the unlabeled competitor increased, but not the mutant probes (Fig. 2c). In addition, the Y1H assay further verified that StbZIP53-like2 could directly bind to StPPO3 (Fig. 2d). These results indicated that StbZIP53-like2 inhibited the expression of StPPO3 by directly and specifically interacting with the C-box motif.
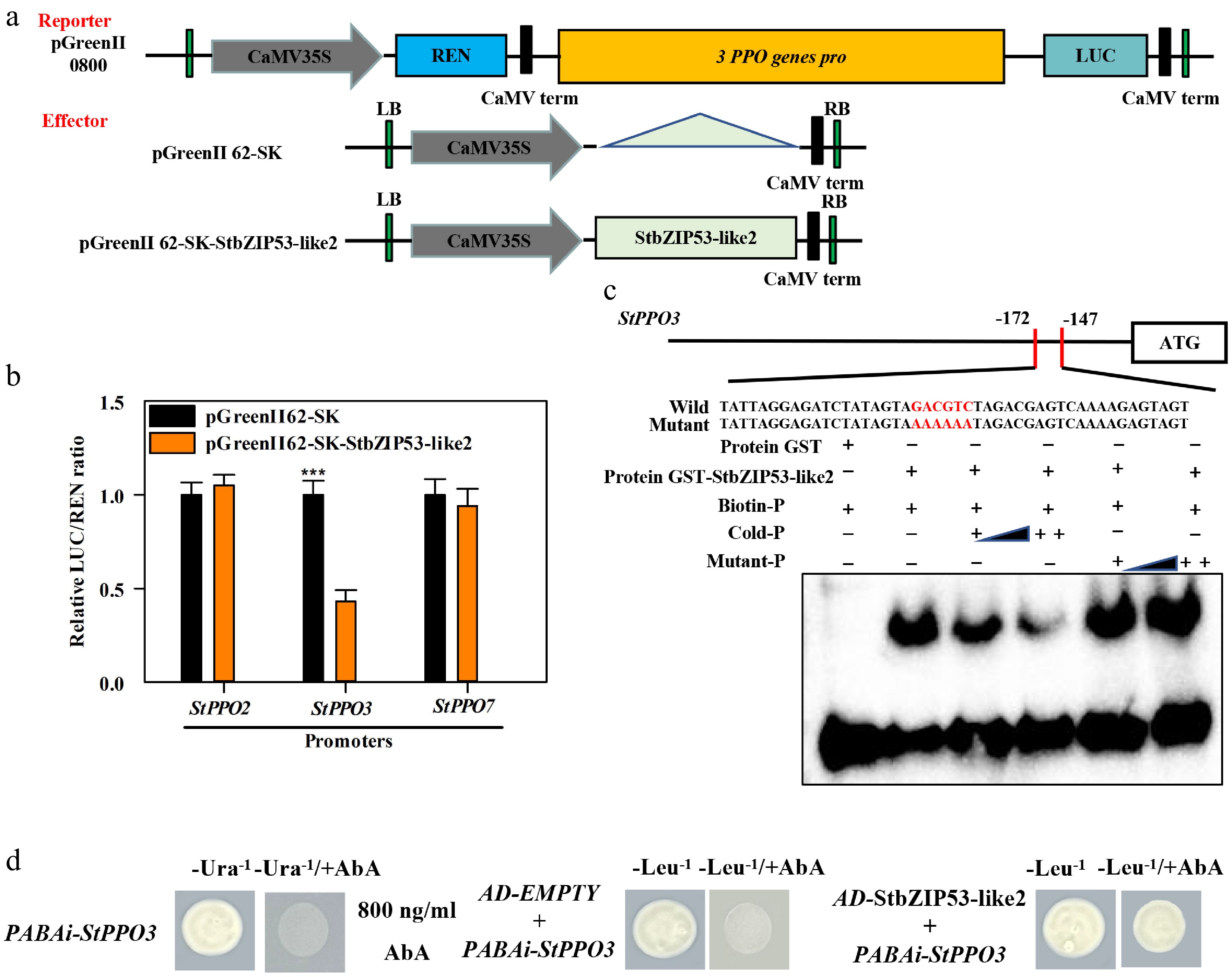
Figure 2.
Effect of StbZIP53-like2 mediates the transcriptional activity of StPPO2, StPPO3 and StPPO7. (a) Picture of the reporters and the effector. (b) The promoter activity of StPPO2, StPPO3 and StPPO7 was regulated by StbZIP53-like2. The empty vector co-expressed with promoters was used as a control (set as 1). *** indicates a significant difference at p < 0.001. (c) The cooperation of StbZIP53-like2 with the promoter of StPPO3 in the EMSAs. GST-StbZIP53-like2 protein was mixed with the probes of StPPO3 including the C-box motif and the mutant probes AAAAAA, which are shown in red letters. ++ indicates increasing amounts of the probe; – represents absence; + represents presence. (d) StbZIP53-like2 interacts with the promoter of StPPO3 in vivo. Yeast growth assays indicated the interaction of StbZIP53-like2 with the promoters of StPPO3.
StbZIP53-like2 interacts with StERF091
-
We aimed to detect whether StbZIP53-like2 interacts with other proteins in the browning of fresh-cut potatoes. As shown in Supplementary Fig. S2, BD-StbZIP53-like2 does not have self-activation ability, and it was used to screen the cDNA library. StERF091 protein was identified. The yeast two-hybrid assay (Y2H data) indicated that BD-StERF091 also does not have self-activation ability, and StERF091 interacted with StbZIP53-like2 in the yeast strain (Fig. 3a). In addition, GST-StbZIP53-like2 could reduce His-StERF091 but not GST protein, which indicated that StERF091 cooperated with StbZIP53-like2 in vitro (Fig. 3b). Moreover, the co-immunoprecipitation (Co-IP) assay also indicated that StERF091 could immunoprecipitate StbZIP53-like2 protein. The data indicated that StERF091 interacted with StbZIP53-like2 in vitro and in vivo.
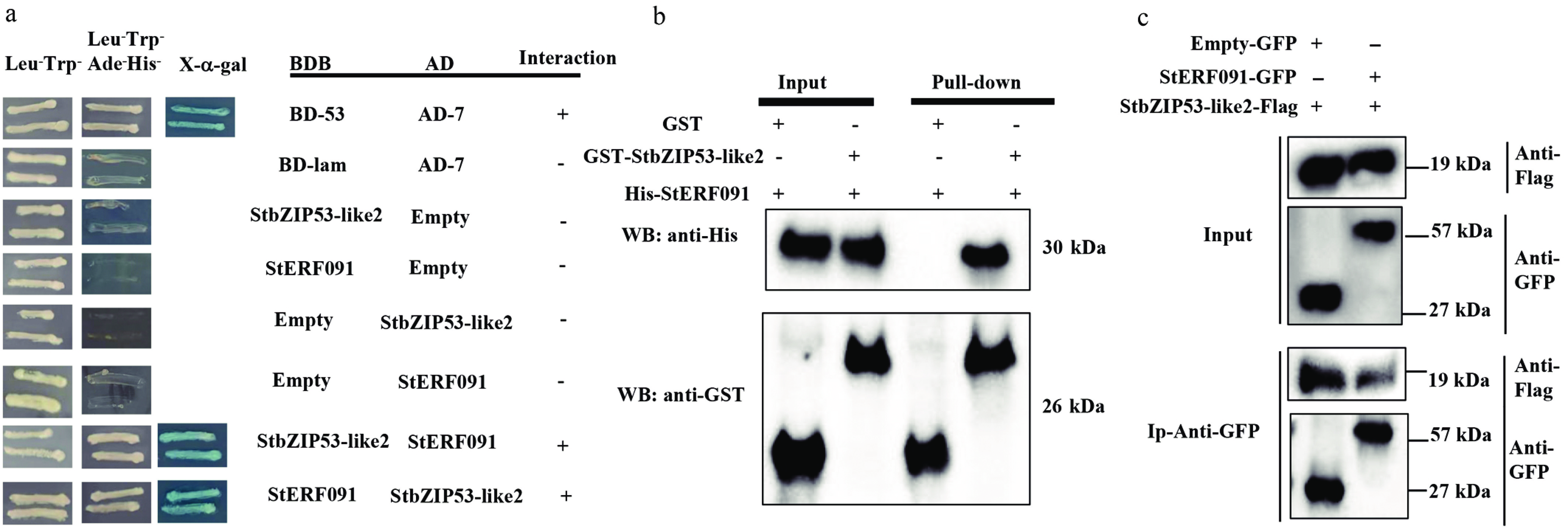
Figure 3.
The cooperation between StbZIP53-like2 and StERF091 proteins. (a) The Y2H assay confirmed the cooperation of StbZIP53-like2 and StERF091. Yeast cells grew on SD/-Leu-Trp-Ade-His- with 125 μM Aureobasidin A and turned blue in the presence of 4 mg/mL X-α-Gal; this served as a positive interaction. (b) GST pull-down assay of the cooperation between StbZIP53-like2 and StERF091. His-StERF091 protein was incubated with GST-StbZIP53-like2 or GST. The anti-His antibody and anti-GST antibody were used for the immunoblotting assays. (c) The co-immunoprecipitation assay confirmed the interaction between StbZIP53-like2 and StERF091. Here, – represents the absence and + represents the presence of the fusion proteins. The anti-Flag and anti-GFP antibodies were used for immunoprecipitation.
Glu induced StERF091 expression and repressed StPPO2 and StPPO3 expression
-
The expression of StERF091 first increased and then decreased, which was enhanced by Glu treatment (Fig. 4a). Moreover, StERF091 contained 261 amino acids and the AP2 domain (Supplementary Fig. S3) and it was located in the nucleus (Fig. 4b).
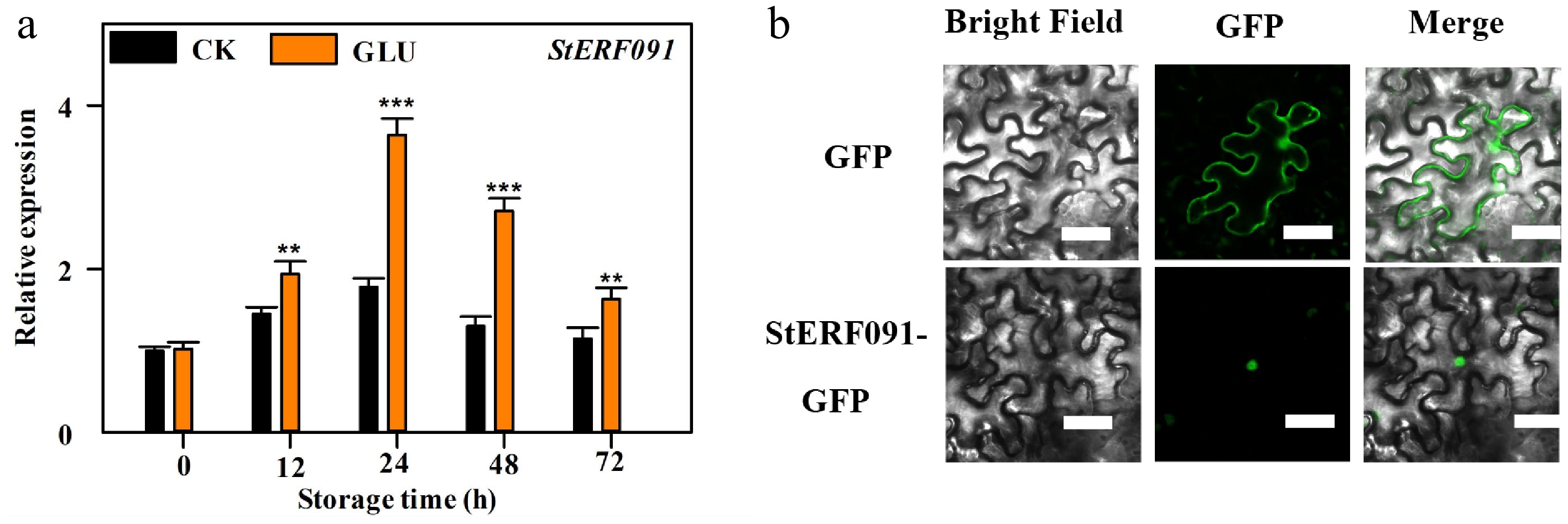
Figure 4.
The expression and subcellular location of StERF091. (a) The expression of StERF091. The expression levels of StERF091 at different points are relative to 0 h, set as 1. ** and *** indicate significant differences at p < 0.01 and p < 0.001, respectively. (b) The subcellular location of StERF091. Bars = 50 μm.
To detected whether StERF091 mediates the promoter activity of StPPO2, StPPO3 and StPPO7, a DLR assay was performed. The promoters of StPPO2, StPPO3 and StPPO7 were separately cloned into the pGreenII 0800-LUC reporter vector, and the CDS of StERF091 was cloned into the pGreenII 62-SK effector vector (Fig. 5a). The promoter activity of StPPO2 and StPPO3 was severely repressed by StERF091, but the promoter of StPPO7 was not regulated by StERF091 (Fig. 5b). EMSA was used to verify whether StERF091 bound the promoter of StPPO2 and StPPO3. The data indicated that GST-StERF091 could directly interact with the GCC-box motif of StPPO2 and StPPO3 and caused mobility shifts, but GST protein alone did not cause a mobility shift. Moreover, the shifted bands were reduced as the unlabeled competitor increased, but not the mutant probes (Fig. 5c). Additionally, the Y1H assay further verified that StERF091 could directly bind to StPPO2 and StPPO3 (Fig. 5d). These results indicated that StERF091 inhibited the expression of StPPO2 and StPPO3 by directly and specifically interacting with the GCC-box motif.
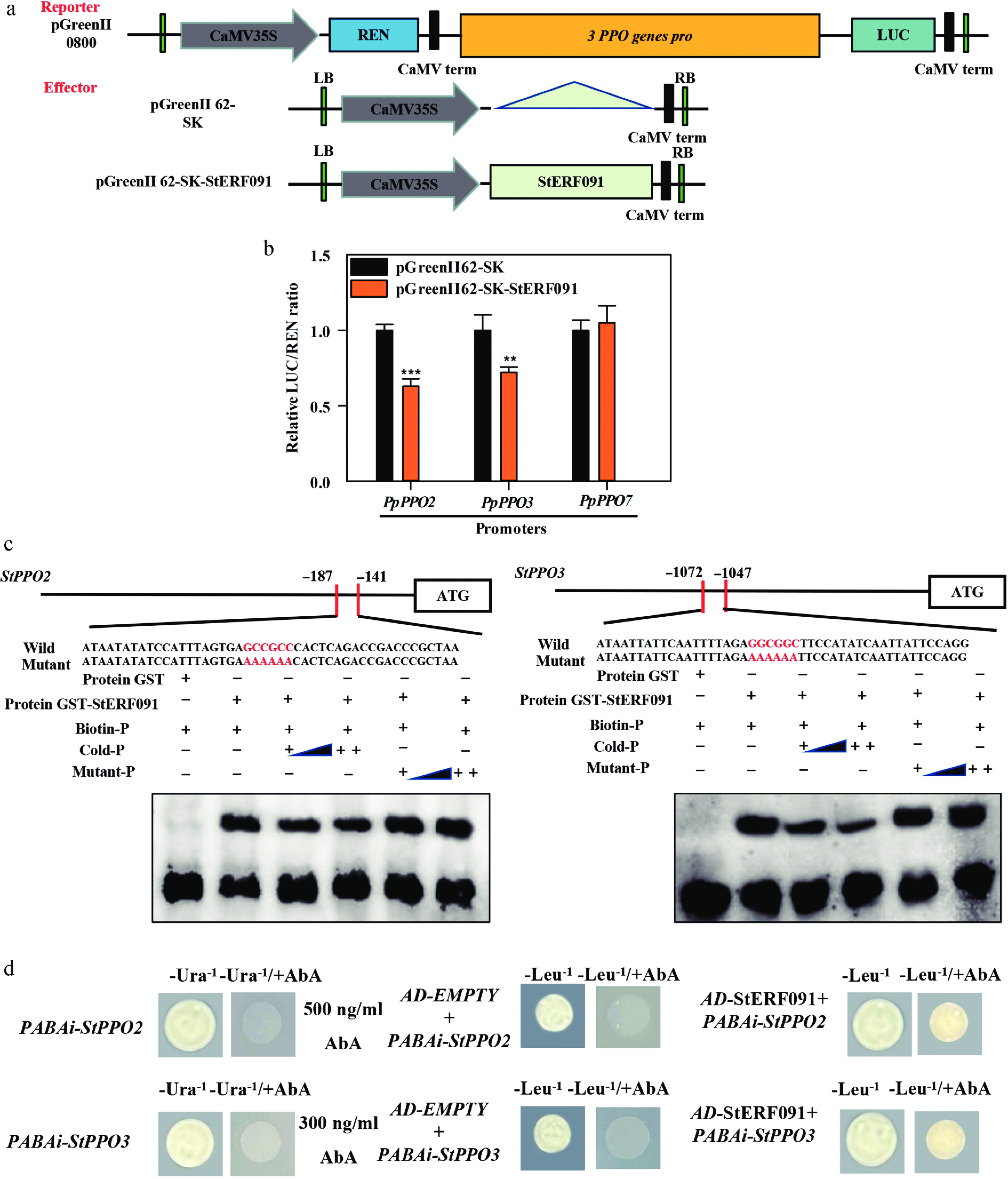
Figure 5.
StERF091 mediates the transcriptional activity of StPPO2, StPPO3 and StPPO7. (a) Picture of the reporters and the effector. (b) The promoter activity of StPPO2, StPPO3 and StPPO7 was regulated by StERF091. The empty vector co-expressed with the promoters was used as a control (set as 1). ** and *** indicate significant differences at p < 0.01 and p < 0.001. (c) The cooperation of StERF091 with the promoter of StPPO2 and StPPO3 in EMSAs. GST-StERF091 protein was mixed with the probes of StPPO2 and StPPO3 including the GCC-box motif and the mutant probes AAAAAA (shown in red letters). ++ indicates increasing amounts of the probe, – represents absence and + represents presence. (d) StERF091 interacts with the promoter of StPPO2 and StPPO3 in vivo. Yeast growth assays indicated the interaction of StERF091 with the promoters of StPPO2 and StPPO3.
StbZIP53-like2 interacted with StERF091 to inhibit StPPO2 and StPPO3 expression
-
The above data indicated that StbZIP53-like2 interacted with StERF091, StbZIP53-like2 repressed StPPO3 expression and StERF091 repressed StPPO2 and StPPO3 expression. Therefore, we explored whether the cooperation of StbZIP53-like2 and StERF091 affects the regulation of its downstream genes. The DLR results showed that when StbZIP53-like2 and StERF091 were co-expressed in tobacco leaves, the transcriptional activity of StPPO2 and StPPO3 was lower than when StbZIP53-like2 or StERF091 was expressed alone (Fig. 6).
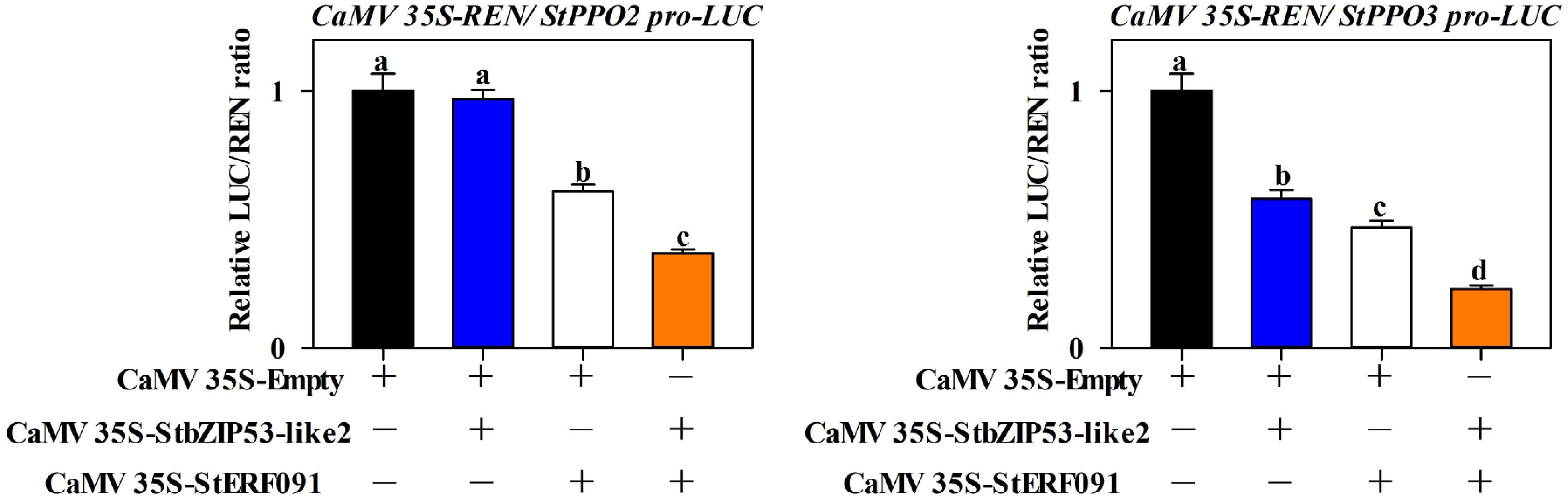
Figure 6.
Combinatory effects of StbZIP53-like2 and StERF091 in mediating the expression of StPPO2 and StPPO3. The empty vector co-expressed with the promoters was used as a control (set as 1). The letters a, b, c and d indicate significant differences at p < 0.05.
-
The browning of fruit and vegetables severely reduces its commercial value. TFs play a key role in regulating the browning of vegetables and fruit. PuMYB21 and PuMYB24 regulate the peel-browning of 'Nanguo' pear by stimulating the transcriptional activity of PuPLDβ1 by directly binding to its promoter[37]. CAMTA5 alleviates the browning of loquat (Eriobotrya japonica) fruit by inhibiting the expression of EjLOX5 and EjPLC6-like, which are the key genes in membrane lipid metabolism[38]. In pineapple (Ananas comosus), AcbHLH144 represses the internal browning of fruit by inhibiting transcription of the phenolic biosynthesis gene Ac4CL5[39]. In peach, PpERF17 alleviates fruit browning by activating the jasmonic acid (JA) signaling pathway[40]. In banana, transient overexpression of MaWRKY18/33/40/60 significantly increases browning of the fruit peel[41]. In apple, MdHY5 and MdHYH inhibits fruit browning by repressing the expression of MdPOD and MdPPO, and enhances MdPAL expression[42]. MdWRKY31 increases the browning of fruit by stimulating the transcription of MdLAC7[43], and MdWRKY3 also enhances the browning of fruit by activating MdPPO7 expression[44]. All this information indicates that there is a complex of transcriptional regulation acting in the browning of fruit and vegetables.
Some research has reported that bZIP TFs regulate fruit and vegetable browning. JrbZIP55, JrbZIP70, JrbZIP72 and JrbZIP88 play a key role in explant browning[45]. In apple, MdbZIP44 aggravates the browning of fruit by stimulating the expression of MdPPO2[46]. However, how bZIP TFs regulate the browning of fresh-cut fruit and vegetables was not reported. Herein, the data first identified the bZIP TF StbZIP53-like2, whose expression was enhanced by the Glu treatment, and StbZIP53-like2 could repress the expression of StPPO3 by directly binding its promoter (Figs 1 and 2).
Plant hormones play a key role in fruits' development, maturation and ripening. Ethylene promotes fruit ripening in banana, tomato and papaya (Carica papaya)[47]. Additionally, some studies found that ethylene also affects the ripening of fresh-cut fruit and vegetables. For example, ethylene represses the internal browning of 'Gala' apples[48]. Ethylene inhibits the peel browning of 'Huangguan' pear by inhibiting the activity of PPO[49]. The browning of 'Yali' pear was reduced by reducing ethylene production[50]. However, ethylene also induces the browning of fruit and vegetables. For example, ethylene induces the browning of pomegranate (Punica granatum) peel by regulating the antioxidant and fatty acid content[51]. Ethylene increases the browning of apple fruit flesh by inducing the expression of MdPPO and MdERF106[52]. In this study, Glu treatment enhanced the ethylene content in fresh-cut potato (Fig. 1). Moreover, several studies have reported that ERFs regulate fruit and vegetable browning. NnERF4/5 were the main regulators in regulating the browning of fresh-cut lotus root[53]. Our previous study found that StERF-BR1-like positively regulates StPPO2 expression by directly binding its promoter, and the expression of StERF-BR1-like was repressed by Glu treatment[3]. In this study, the expression of StERF091 was increased by Glu treatment, and StERF091 significantly inhibited the expression of StPPO2 and StPPO3 by directly binding their promoter (Figs 4 and 5). All these data indicate that Glu treatment alleviates the browning of fresh-cut potatoes by enhancing the expression of StbZIP53-like2 and StERF091, which further inhibits the expression of StPPO2 and StPPO3.
Protein interactions play a key role in fruit and vegetable development, maturation and resistance to biotic and abiotic stresses. In peach, PpERF5 interacts with PpERF7 and enhances fruit aroma by activating PpLOX4 transcription[34]. PpZAT6 cooperates with PpWRKY46 to form a complex enhancing the fruit's cold tolerance by activating the expression of proline synthesis genes[54]. PpWRKY22 cooperates with PpWRKY70 to form a complex improve fruits' resistance to Monilinia fructicola by enhancing the gamma-aminobutyric acid shunt[55]. In banana, MaEBF1 interacts with MaABI5-like to enhance MaABI5-like, regulating starch and cell degradation[56]. In tomato, SlWRKY42 interacts with SlMYC2, and increased SlWRKY42 regulates SlSPDS2 expression and promotes spermidine accumulation[57]. In lychee (Litchi chinensis), LcWRKY1 interacts with LcNAC1 to form a complex regulating LcAOX1 expression[58]. In apple, MdWRKY41 cooperates with MdMYB16 to form a complex to regulate anthocyanin and proanthocyanidin biosynthesis by repressing the expression of MdANR and MdUFGT[59]. Moreover, protein interactions also affects TFs regulating the browning of fruit and vegetables. PuMYB21 interacts with PuMYB24, and this interaction mediates the browning of 'Nanguo' pear by enhancing the transcription of PuPLDβ1[37]. However, protein interactions mediating the browning of fresh-cut fruit and vegetables are not yet widely reported. In this study, the data shown that StbZIP53-like2 interacts with StERF091 to form a complex to mediate the browning of fresh-cut potatoes by inhibiting the expression of StPPO2 and StPPO3 (Fig. 6).
-
This study found two new transcription repressors, StbZIP53-like2 and StERF091, whose transcription was enhanced by Glu treatment. StbZIP53-like2 repressed the expression of StPPO3, and StERF091 repressed the expression of StPPO2 and StPPO3 by directly interacting with their promoters. Additionally, StbZIP53-like2 interacts with StERF091 to form a complex, which enhanced the inhibition of StPPO2 and StPPO3, thereby alleviating the browning of fresh-cut potatoes (Fig. 7). Taken together, the data uncover a StbZIP53-like2–StERF091 module that alleviates the browning of fresh-cut potatoes under Glu treatment.
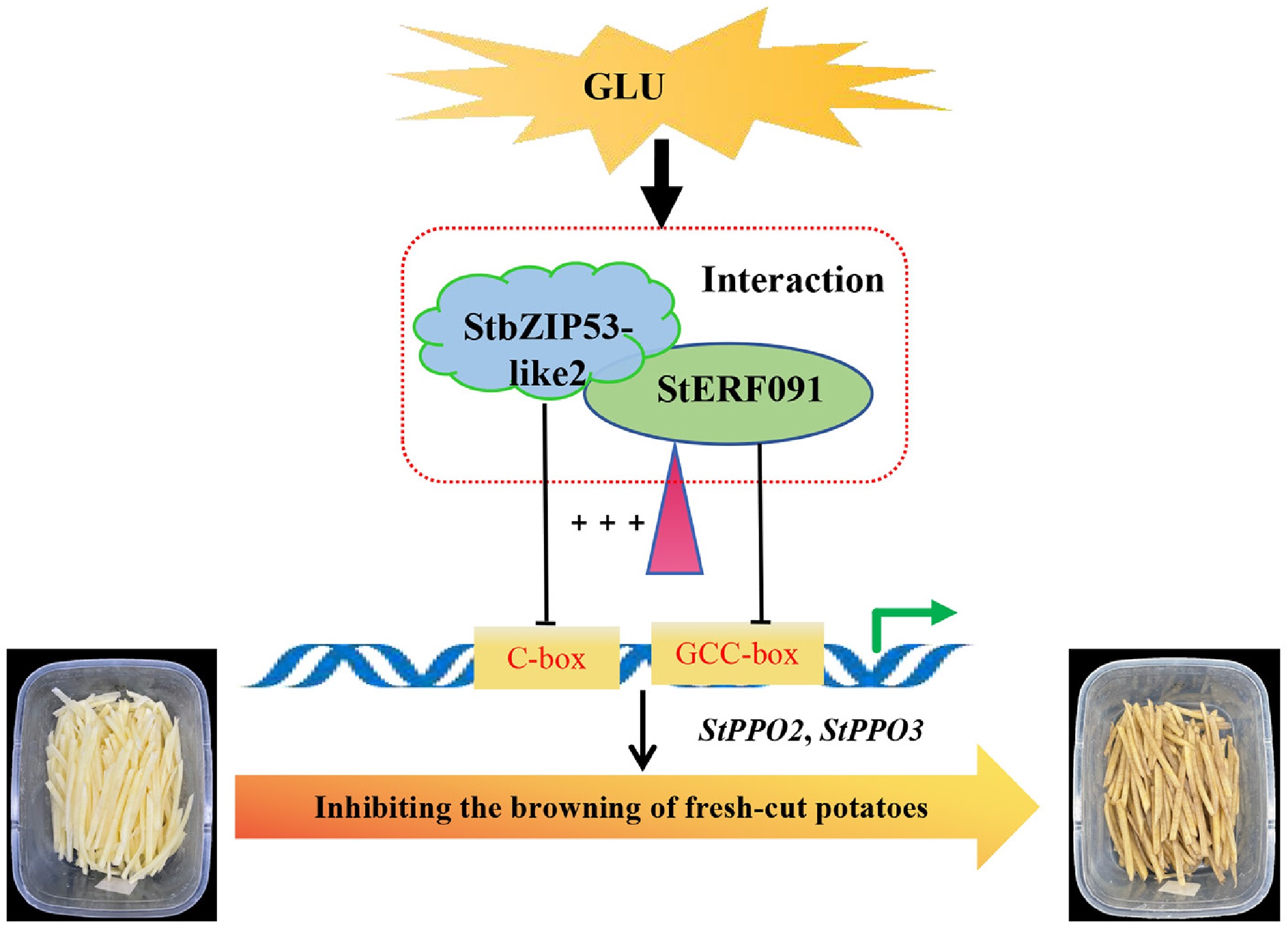
Figure 7.
Glu alleviates the browning of fresh-cut potatoes by enhancing the expression of StbZIP53-like2 and StERF091, thus inhibiting the transcriptional activity of StPPO2 and StPPO3.
-
The authors confirm their contributions to the paper as follows: methodology, formal analysis, draft manuscript preparation: Li W; data curation: Li W, Cao J, He Y, Wang Y; investigation: Li W, Cao J; validation: Cao J, Shi J, Song Z; conceptualization, writing − review & editing, supervision, resources: Shi J, Song Z. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
This work was supported by the Natural Science Foundation of Shandong Province (ZR2023QC010), the Key R & D project of Shandong Province (Grant No. 2021TZXD007) and the Potato Industry Innovation Team for Modern Agricultural Industry Technology System of Shandong Province (SDAIT-16-11).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Wenhui Li, Jixuan Cao
- Supplementary Table S1 Primers used in present study.
- Supplementary Fig. S1 The construction of StbZIP53-like2 protein, which was predicted by CD-search (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi).
- Supplementary Fig. S2 cDNA library quality testing.
- Supplementary Fig. S3 The construction of StERF091 protein, which was predicted by CD-search (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi).
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Chongqing University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li W, Cao J, He Y, Wang Y, Shi J, et al. 2026. A StbZIP53-like2–StERF091 module regulates the browning of fresh-cut potatoes. Plant Hormones 2: e007 doi: 10.48130/ph-0026-0004 

A StbZIP53-like2–StERF091 module regulates the browning of fresh-cut potatoes
- Received: 20 January 2026
- Revised: 07 February 2026
- Accepted: 24 February 2026
- Published online: 20 March 2026
Abstract: With the acceleration of modern life, fresh-cut potatoes (Solanum tuberosum) are very popular in our daily lives. However, surface browning is the major issue reducing the quality of fresh-cut potatoes during storage. Our previous study found that glutamic acid (Glu) treatment could alleviate the browning of fresh-cut potatoes; however the regulatory mechanism was still unclear. In this study, Glu treatment markedly enhanced the expression of StbZIP53-like2, which was a nuclear protein. StbZIP53-like2 repressed the transcriptional activity of StPPO3. StbZIP53-like2 cooperated with an ethylene responsive factor (ERF) transcription factor, StERF091. The transcript level of StERF091 was also enhanced by Glu treatment, and StERF091 was a nuclear protein. Moreover, StERF091 inhibited the transcriptional activity of StPPO2 and StPPO3. Moreover, the interaction of StbZIP53-like2 and StERF091 reduced the transcriptional activity of StPPO2 and StPPO3 compared with StbZIP53-like2 and StERF091 alone. Taken together, these results uncovered a dynamic regulatory module of StbZIP53-like2–StERF091 mediating the browning of fresh-cut potatoes.
-
Key words:
- Fresh-cut potatoes /
- StbZIP53-like2 /
- StERF091 /
- Browning