









-
Plants have three types of flowers: female, male, and hermaphroditic[1,2]. Different combinations of these three types of flowers on the same plant determine the diversity of plant sex differentiation[3]. According to the types of floral organs and their growth characteristics, plants can mainly be classified into seven sex types: androecious line (producing only male flowers), gynoecious (producing only female flowers), monoecious line (unisexual male and unisexual female flowers), gynomonoecious line (hermaphroditic and unisexual female flowers), andromonoecious line (hermaphroditic and unisexual male flowers), trimonoecious line (hermaphroditic, unisexual male and female flowers), and the hermaphroditic line (producing only hermaphroditic flowers)[4−10]. These sex differentiation types can all be found in cucurbit plants[11,12]. Watermelon (Citrullus lanatus) is an important worldwide horticultural economic crop[13]. Most cultivated varieties of watermelon are the monoecious line, with some varieties exhibiting the andromonoecious line. The hermaphroditic flowers of watermelon are prone to self-pollination. During hybrid breeding, hermaphroditic flowers need to be dehulled as the mother plants, which can easily lead to the generation of pseudo-hybrids due to incomplete dehulling[14,15]. Therefore, sex differentiation research is particularly important for breeders.
The cucurbit plants are regarded as models for studying sexual differentiation because they encompass all seven types of sexual differentiation[3]. To date, the ACS (1-Aminocyclopropane-1-carboxylate synthase) genes identified based on the ethylene-mediated sex determination model in cucurbits include CsACS1G[16], CsACS2[17], CsACS11[18], CmACS7[19], CmACS11[20], and ClACS7/CitACS4[21]. The sexual differentiation of cucumbers is jointly regulated by three sexual-determining genes, F (CsACS1G), M (CsACS2), and A (CsACS11). Among them, the F gene promotes the development of female flowers, while the M gene is expressed in the carpels of female flowers and inhibits the formation of stamens. The mutation m leads to the production of hermaphroditic flowers[16,22]. The A gene is a key gene for carpel development and is a pure male control gene. Its mutant form in cucumber shows pure male flowers without female flowers[18]. Notably, a recent study has shown that auxin and ethylene exhibit a reciprocal relationship during sex determination, while CsARF3 plays an essential role in this process[23]. The sex differentiation of melon is mainly controlled by three pairs of alleles A/a (CmACS7), G/g (CmACS11), and M/m. The single-sex female flower of melon is controlled by the dominant single gene A; the a gene controls the formation of male and hermaphroditic flowers on the same plant; the G gene controls pure female plants, and the g gene is related to female and hermaphroditic flowers; the M gene is related to the differentiation of male flowers[19,20]. The most common four types of sex morphologies in melon are pure female plants (__ggM_), female and male heteromorphic flowers on the same plant (A_G_M), male and hermaphroditic flowers on the same plant (A_G_mm), and hermaphroditic flower plants (_ggmm)[24]. The sex differentiation of watermelon is controlled by three pairs of alleles: a (CitACS4/ClACS7)[3,21], gy (ClWIP1)[12], and tm. The bisexual flower on the same plant as both male and female flowers is controlled by the recessive gene a, the same plant sex type of bisexual male, bisexual female, and bisexual and hermaphroditic flowers is controlled by the recessive gene tm, and a is the recessive supergene of tm[3]. The all-female plant sex type of watermelons is controlled by the recessive single gene gy[12,14].
In summary, the m gene in cucumber, a gene in melon, and a gene in watermelon all regulate the formation of bisexual flowers, and the research on their regulatory mechanisms is relatively in-depth. However, for watermelon, there is a lack of positive genetic evidence for the a gene. Therefore, based on the previous BSA-seq and preliminary positioning, our study expanded the ZXG1555 × COS-F2 population, used the flanking markers from the preliminary positioning to screen for recombinant single plants, developed polymorphic molecular markers within the preliminary positioning interval, combined the genotypes and phenotypes of the recombinant single plants to conduct fine mapping of the key genes controlling the bisexual flower trait of watermelon, established, and verified the watermelon bisexual flower molecular marker-assisted selection technology system, which is of great significance for watermelon gender differentiation breeding.
-
Two watermelon varieties, ZXG1555 (andromonoecious) and COS (Cream of Saskatchewan, monoecious), were used as parental lines to construct the mapping population. In both the spring and autumn of 2021, 187 plants and 137 plants of the F2 population were respectively planted in the Xiangyang Experimental Agricultural Farm and Horticultural Engineering Center greenhouse at the Northeast Agricultural University. Both the spring and autumn populations were used for QTL analysis, with the aim of obtaining stable preliminary mapping intervals. In the spring of 2022, another 710 F2 plants derived from the same parental lines were used for fine mapping. Phenotypic statistics of open female flowers were conducted in the early morning during both spring and autumn, and the number of stamens on each female flower was recorded. At least four female flowers were counted per plant. The andromonoecious index (AI value) of the female flowers was calculated according to the definition of Martínez et al.[25], and the female flowers without stamen development were defined as AI = 1, female flowers with 1 or 2 stamens, or without pollen production were defined as AI = 2, and female flowers with stamens greater than or equal to 3 and producing pollen were defined as AI = 3. The average AI value of each individual plant in the segregating population was calculated, and female flowers with AI ≤ 1.5 were classified as unisexual flowers, and female flowers with AI > 1.5 were classified as non-unisexual flowers. Flower bud tissues of the two parents were collected for the determination of the expression levels of candidate genes.
QTL analysis
-
Based on the resequencing data of the parental lines and the reference genome of watermelon (97103 v2) from the CuGenDB (
http://cucurbitgenomics.org/organism/21 ), flanking sequences (500−700 bp) of 12 SNPs were extracted, which are located in the initial mapping region (Chr3: 29,252,650–30,021,268 bp)[26]. Using information on the restriction sites of 10 enzymes (including EcoRI, MboI, HincII, HindIII, PstI, MspI, XhoI, TaqI, BamHI, and BclI), candidate SNPs were converted into cleaved amplified polymorphic sequence (CAPS) markers with the SNP2CAPS software[27]. Primers were designed using Primer Premier 6.0 (www.premierbiosoft.com/primerdesign ), with a length of 20–30 bp, and GC content of 40%–60% (Supplementary Table S1). Markers that demonstrated co-dominant polymorphism in the P1, P2, and F1 generations were employed for genotyping the F2 population.A total of 12 CAPS markers were developed to genotype the F2 population in both spring and autumn of 2021. Combined with phenotypic data and genotypic information, QTL analysis was conducted using the R/qtl software[28].
Fine mapping
-
Based on the results of QTL analysis and preliminary mapping, the F2 population was expanded to 710 individuals. Recombinant plants were screened using molecular markers flanking the initial target region. Additionally, seven CAPS markers (Supplementary Table S1) were newly developed within this region for genotyping the recombinants. Each recombinant plant was self-pollinated to produce F2:3 families, with each family comprising at least 20 individuals. The trait of bisexual flowers in watermelon was assessed during the flowering period. By integrating genotype data from the F2 plants and phenotype data from the F2:3 families, the key gene governing bisexual flower formation in watermelon was finely mapped.
Candidate gene prediction
-
Using the reference genome of watermelon (97103 v2), the candidate genes within the fine-mapping interval were annotated. The coding sequences of the candidate genes were compared in the resequencing of the two parental lines using the Integrative Genomic Viewer (IGV) software[29] to detect the correlations of SNPs or structural variations with the phenotype.
Flower bud tissues (3 d before flowering) collected from the two parental lines were stored in liquid nitrogen for RNA extraction. Total RNA extraction was carried out according to the instructions of the Sangon (Shanghai, China) RNA Extraction Kit. The extracted RNA was first subjected to agarose gel electrophoresis for detection. Then, cDNA synthesis was performed using the Vazyme Reverse Transcriptase Kit (Nanjing, China). Specific primers (Supplementary Table S2) were designed based on the website for qRT-PCR primer design of Genscript (
www.genscript.com.cn/tools/real-time-pcr-taqman-primer-design-tool ). The analysis of gene expression levels was carried out according to the previous method[30,31].Sequence analysis
-
We obtained the homologous protein sequences of ClACS7 from the cucurbits by conducting a BLAST search in the NCBI database (
www.ncbi.nlm.nih.gov/cdd ). Multiple protein sequence alignment and phylogenetic tree construction were performed using MEGA 11.0 based on the Neighbor-Joining method[32]. The 3D structures of the ClACS7 protein from the two parental lines were modeled with AlphaFold 3 and compared in PyMOL software[33].Marker-assisted selection (MAS) system verification
-
By using molecular markers that are closely linked to the sex-related traits within the fine mapping region, the genotypes of 32 different types of sex differentiated (monoecious and andromonoecious) watermelon germplasm resources were determined. Combined with the phenotypic data of the natural population, a molecular marker-assisted selection technology system for watermelon sex differentiation was established and verified. The 32 watermelon materials are listed in Supplementary Table S3. In addition, sequences 75 bp upstream and downstream of the SNPs were selected, and a Kompetitive Allele Specific PCR (KASP) primer was designed using the previous method as described by Fang et al.[34].
Statistical analysis
-
The results are expressed as mean ± SD from at least three independent replicates. Distinct letters indicate statistically significant differences. Statistical comparisons among groups were performed by one-way ANOVA followed by Tukey's post hoc test, with a significance threshold of p < 0.05. All graphs were generated using Prism 10.0 (GraphPad Inc., USA).
-
To obtain a stable preliminary mapping interval for the andromonoecious (a) locus of watermelon, a total of 12 CAPS markers within the initially located interval and genotyping was conducted on the F2 populations in the spring (187 plants), and autumn (137 plants) of 2021, respectively (Fig. 1). The QTL analysis indicated that the a locus was located in the same region (Chr3: 29,252,650–30,021,268 bp, approximately 768 kb), in both spring (Fig. 2a) and autumn (Fig. 2c) populations in 2021. This region was consistent with the initial mapping region. Moreover, there was a significant peak at the marker Chr3_29540662 (for spring, the LOD value was 12.73, with a phenotypic variation of 55.38%; for autumn, the LOD value was 11.98, with a phenotypic variation of 46.18%, Table 1). By analyzing the phenotypes and genotypes of individual plants in the F2 population using this marker, it was found that when the genotype was AA or AB, the phenotypic AI value was greater than 1.5, indicating andromonoecious type (bisexual flowers); when the genotype was BB, the AI value was less than 1.5, indicating monoecious type (unisexual flowers) in the both spring (Fig. 2b), and the autumn of 2021 (Fig. 2d). Taken together, our data demonstrate that the markers located on both sides of the initial mapping interval (Chr03_29252650 and Chr03_30021268) can be used for subsequent fine mapping.

Figure 1.
Sexual differentiation phenotypic characteristics in the parental lines and F1 and F2 populations. AI, andromonoecious index. Bar = 1 cm.
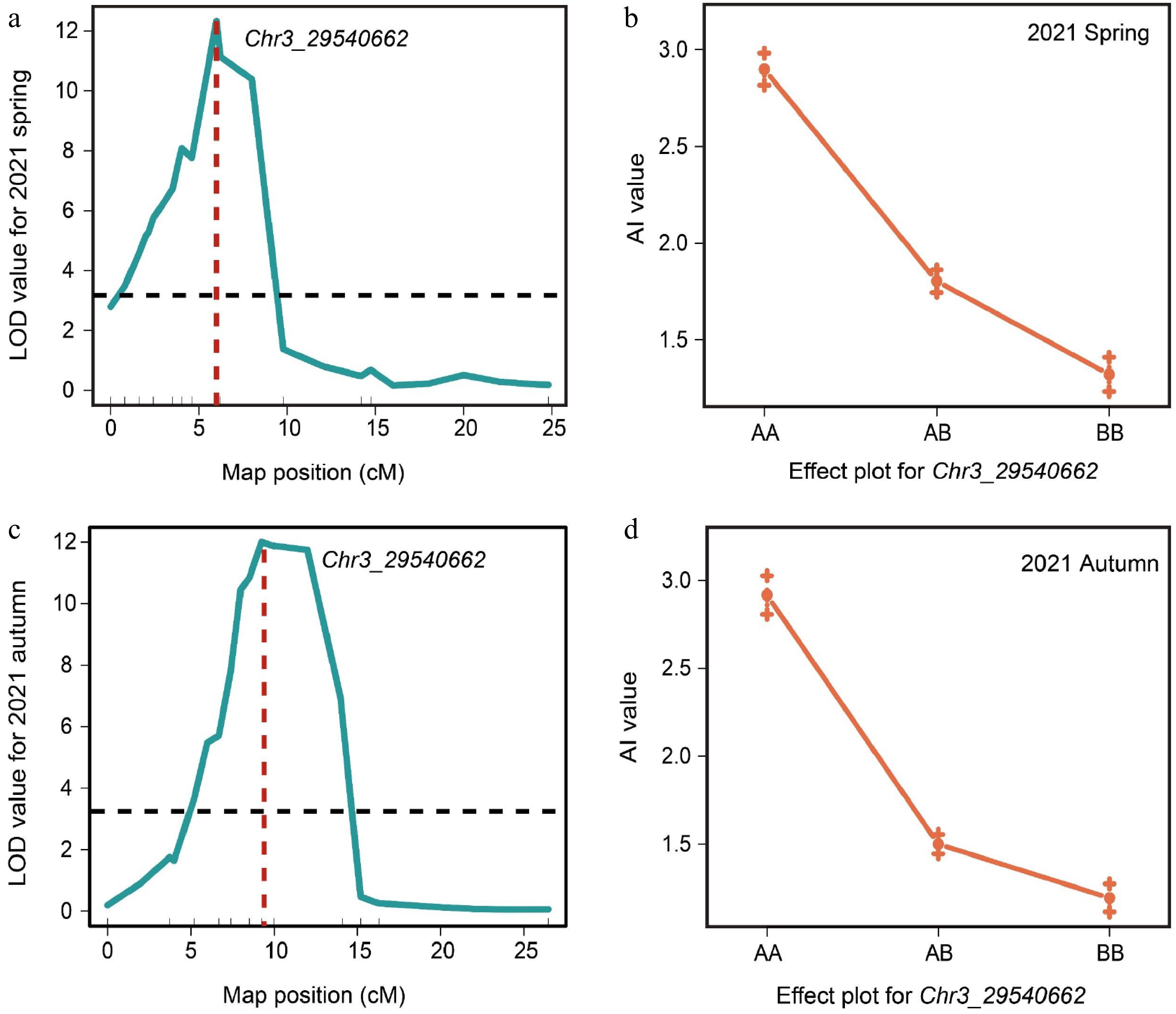
Figure 2.
QTL analysis of the a locus based on the initial mapping region. (a) QTL analysis of F2 population in spring 2021. (b) Correlations between phenotypes and genotypes based on the marker Chr3_29540662 in spring 2021. (c) QTL analysis of F2 population in autumn 2021. (d) Correlations between phenotypes and genotypes based on the marker Chr3_29540662 in autumn 2021.
Table 1. QTL for monoecious/andromonoecious based on interval mapping in the F2 population from both spring and autumn in 2021.
Year Left marker Right marker LOD Phenotypic
variation (%)2021 Spring Chr03_29252650 Chr03_30021268 12.73 55.38 2021 Autumn Chr03_29252650 Chr03_30021268 11.98 46.18 a locus was fine-mapped to an 82.10-kb region
-
Initial markers Chr3_29252650 and Chr3_30021268 were used to screen for recombinant individuals from the additional F2 generation (710 plants) of spring 2022 (Fig. 3c). A total of 17 CAPS markers were used to genotype the recombinant individual plants and detect the recombination events. Discarding the plants that only had recombination at the marker loci on both sides, 26 key recombinant plants were finally obtained. Based on the F2:3 phenotypes and genotypes of the recombinant plants, combined with the F2–217 and F2–233 plants no longer recombined between Chr3_29488876 and Chr3_29570980 (29,488,876–29,570,980 bp), the a locus in watermelon was fine-mapped to be located between the markers Chr3_29488876 and Chr3_29570980 (82.10 kb in physical distance) and contained five candidate genes (Fig. 3d).
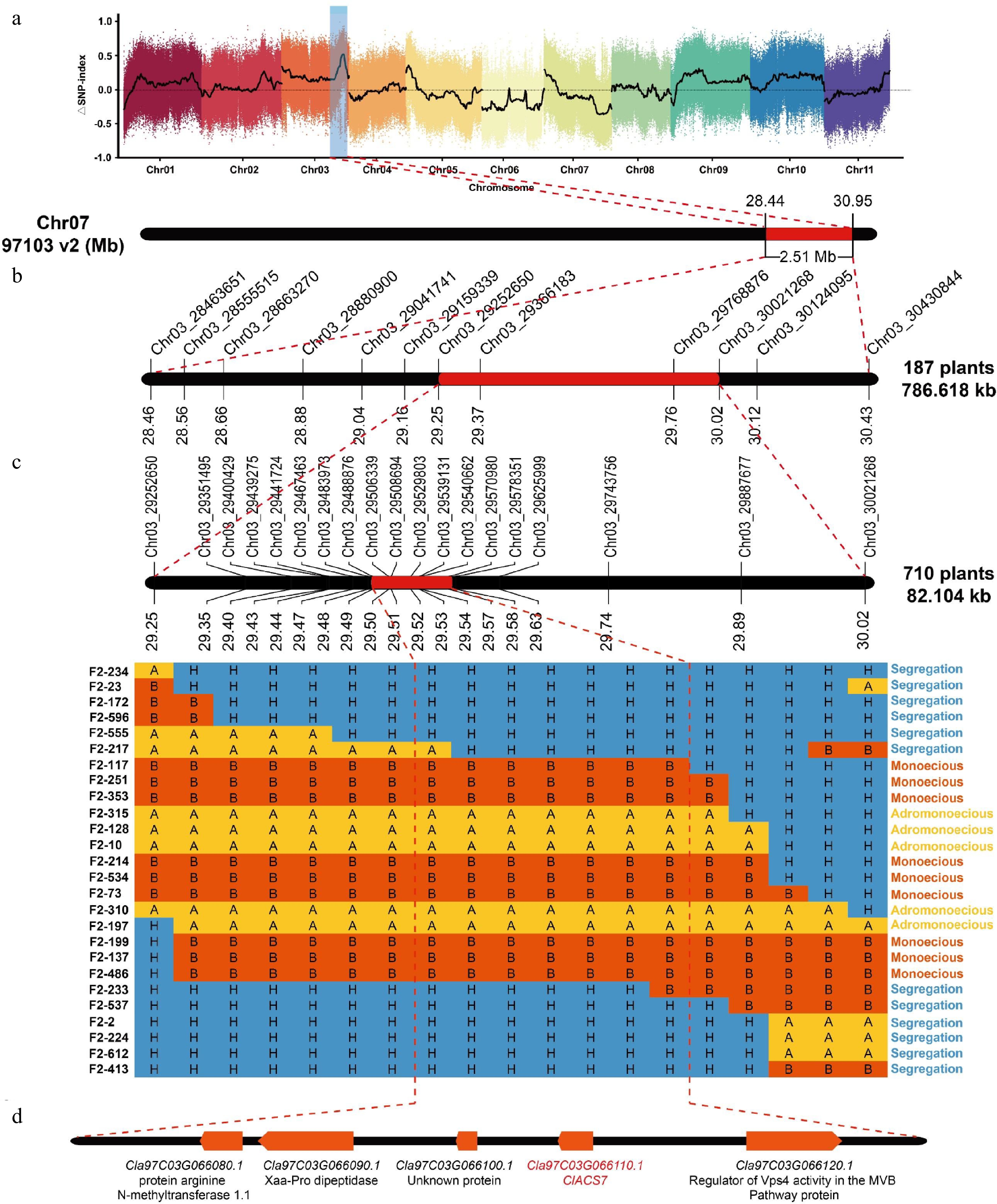
Figure 3.
Fine mapping of the a locus. (a) ΔSNP index plot from BSA-seq. (b) Initial mapping of the a locus to a 786.62-kb region[26]. (c) Fine mapping of the a locus to an 82.10-kb region with 26 recombinant plants. 'A' denotes the ZXG1555 genotype, 'B' represents the COS genotype, and 'H' signifies the heterozygous genotype. (d) The five candidate genes in the fine mapping region.
ClACS7 was predicted to be the candidate gene for a locus
-
Using the reference genome of watermelon in the Cucurbit Genomics Database (97103 v2) (
http://cucurbitgenomics.org/organism/21 ), the candidate genes within the fine mapping interval (29,488,876–29,570,980 bp) were annotated. A total of five candidate genes were included. The homologous gene ClACS7 of CmACS7, which has already been reported, is included. This gene has also been verified in watermelons to be related to the sex differentiation of bisexual flowers[35−37]. Based on the resequencing data of the two parental lines, the coding regions of the candidate genes were first compared in the IGV software, and it was found that four candidate genes had non-synonymous mutations in their coding regions. ClACS7 (Cla97C03G066110, encodes 1-aminocyclopropane-1-carboxylate synthase 7) has two non-synonymous mutations. The first one occurs at the 792nd bp (SNP264), where the base changes from C to G, and the amino acid is replaced from asparagine to lysine; the second non-synonymous mutation is located at the 1,185th bp (SNP384), where the base changes from G to C, and the amino acid is replaced from glutamic acid to glutamine (Fig. 4a). Conserved domain analysis revealed that both SNPs in ClACS7 were located within the PLN02607 domain (corresponding to amino acids 1-440, identified as 1-aminocyclopropane-1-carboxylate synthase, Supplementary Fig. S1). To investigate whether these SNPs affect the 3D protein structure, we predicted the structures using AlphaFold 3 and performed comparisons with PyMOL. The results indicated that neither SNP caused alterations in the protein structure (Fig. 4b). Further hydrogen bond analysis of these SNP sites (Fig. 4c) showed that they altered the number of hydrogen bonds between the protein and its ligand (for SNP264, the number was 2 in COS, and 3 in ZXG1555; for SNP384, it was 5 in COS and 4 in ZXG1555). This change may affect the protein's ligand-binding capacity or overall function[38]. Furthermore, as the box7 domain is a known key regulator of ACS enzyme activity[39], and SNP384 is situated in close proximity to this region, suggesting that SNP384 may be a critical site influencing the enzymatic activity of ClACS7. The annotations and mutation information of the other four genes are listed in Supplementary Table S4.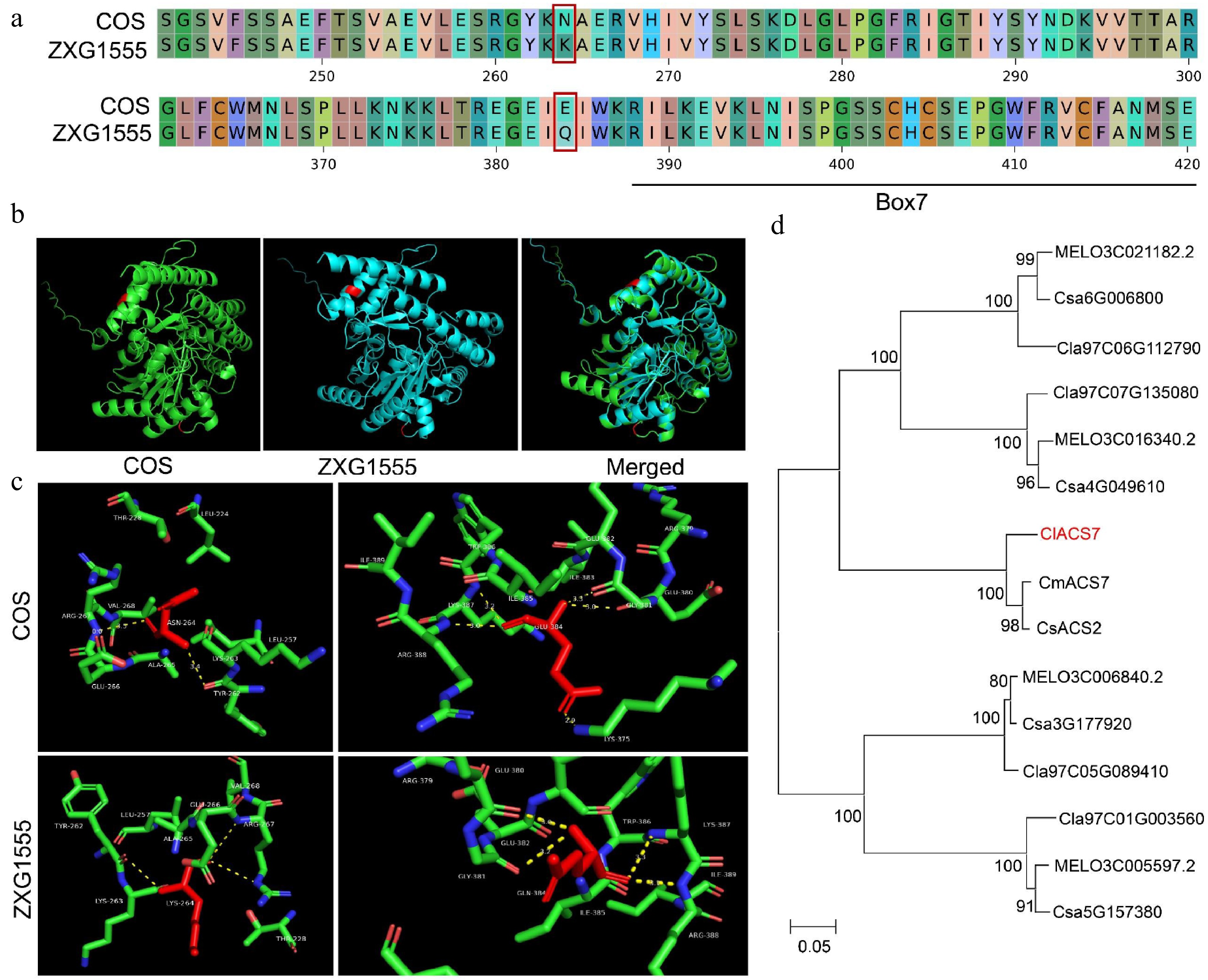
Figure 4.
Sequence analysis of ClACS7. (a) The amino acid alignment of ClACS7 in the two parental lines. The red box lines indicate the amino acid mutations caused by non-synonymous mutations. (b) Prediction and comparison of 3D protein structure of the ClACS7. Red color represents the locations where the two SNPs (SNP264 and SNP384) are located. (c) Hydrogen bond analysis of SNP264 and SNP384. The yellow dotted line represents the number of hydrogen bonds. (d) The phylogenetic tree of ClACS7 homologous genes. CmACS7: MELO3C015444.2, CsACS2: Csa1M580750.
In order to clarify the homologous evolutionary relationship of ClACS7, we identified its homologous genes from melon and cucumber and constructed a phylogenetic tree (Fig. 4d). The results showed that ClACS7 has a high degree of homology with CmACS7 and CsACS2, forming a monophyletic evolutionary branch, indicating that ClACS7 is the direct homolog gene of CmACS7 and CsACS2.
To further validate the associations of candidate genes with the andromonoecious trait, we conducted qRT-PCR analysis on five candidate genes in the two parental lines. The results show that the expression level of ClACS7 in female flower buds is significantly higher than that in bisexual flower buds, while the expression levels of the other four candidate genes show no significant differences between the two parental lines (Fig. 5a).
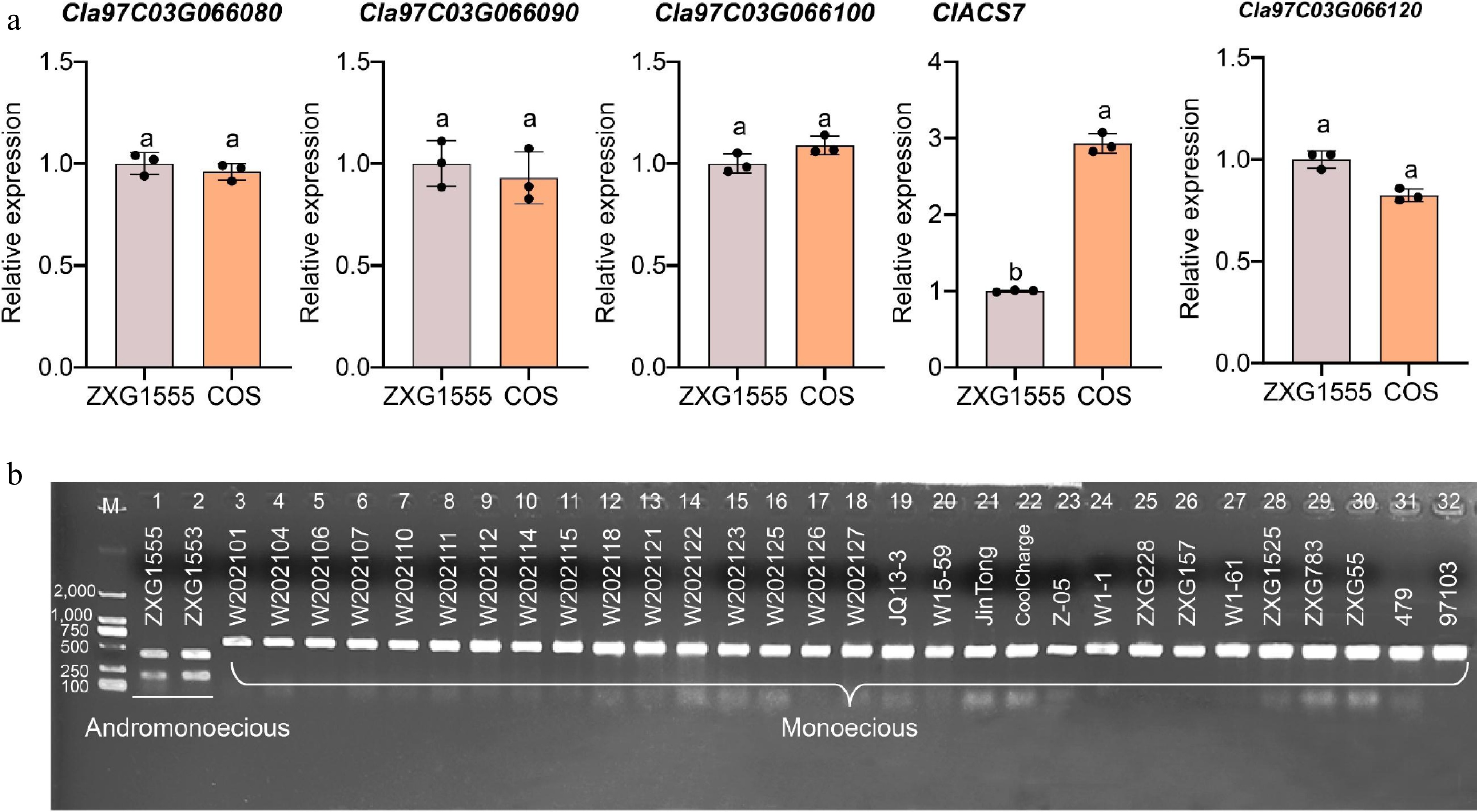
Figure 5.
Determination of the expression levels of five candidate genes and establishment of a molecular marker-assisted selection technology system for monoecious/andromonoecious flowers in watermelon. (a) The expression levels of five candidate genes analysis in the flower buds of the two parental lines. The different letters denote significance (p < 0.05), as determined by analysis of variance. (b) Validation of marker Chr3_29539131 in natural populations of watermelon. M, DL2000 marker. The numbers on the left indicate the size of the marker, bp.
Chr3_29539131 can be used for MAS validation for a locus
-
Using the CAPS marker Chr3_29539131, which is closely linked to the key genes of andromonoecious in watermelon, the genotypes of 32 natural watermelon accessions were determined. In addition, due to the advantages of high throughput and ease of operation of KASP markers[40], we converted SNP29,539,131 into KASP markers and developed three KASP primers, which are listed in Supplementary Table S5. Combined with the field phenotypes, it was found that the enzyme-cut fragments of the andromonoecious materials were 459 and 198 bp, while those of the monoecious materials were 657 bp. This result indicates a strong correspondence between phenotype and genotype (Fig. 5b), indicating that this diagnostic marker can distinguish bisexual and monoecious flowers in watermelon.
-
The sex types of cucurbit plants are highly correlated with ethylene[41−45]. During the biosynthesis of ethylene, methionine is converted into s-adenosyl-methionine, which serves as a precursor in the production of polyamines. The 1-aminocyclopropane-1-carboxylic acid synthase (ACS) catalyzes the conversion of AdoMet into 1-aminocyclopropane-1-carboxylic acid (ACC). Therefore, ACS is the rate-limiting enzyme in ethylene biosynthesis[46−48]. Previous studies have confirmed that in the cucurbits, the function of the ACS gene is relatively conserved in the process of regulating sex differentiation[49,50]. In watermelon, ClACS7 is the homologous gene of CmACS7, CsACS2, and CpACS27A[19,25,51,52]. Like the other three genes, ClACS7 has a similar gene structure (Fig. 4), indicating that these different genes evolved from the same ancestral sequence[35]. In this study, we successfully precisely localized the a gene for watermelon sex differentiation to the 82.10 kb interval on watermelon chromosome 3, and this interval contains ClACS7 (Fig. 3), providing positive genetic evidence for the regulation of watermelon bisexual flower sex differentiation by ClACS7.
To further confirm the correlation between ClACS7 and the andromonoecious phenotype of watermelon, we measured the expression level of ClACS7 in the flower bud tissues of the two parental lines. The results showed that the expression level of ClACS7 in the female flower buds was significantly higher than that in the hermaphroditic flowers (Fig. 5a). Among melon, cucumber, and pumpkin, the homologous genes are specifically transcribed in the pistil flowers, and their expression levels are higher in the pistil flowers than in the staminate flowers[19,25,51]. This indicates that the expression patterns of these homologous genes have also been preserved through evolution. In addition, Boualem et al.'s transcriptional results of ClACS7 in different parts of watermelon indicated that ClACS7 was specifically expressed in the carpels of female flowers and hermaphroditic flowers, but not expressed in male flowers, leaves, and stems[35]. More importantly, the enzyme activity test confirmed that the active ClACS7 enzyme would cause the development of female flowers in hermaphroditic varieties, while the reduction of enzyme activity would result in hermaphroditic flowers in the androgyne varieties. This indicates that the ethylene production mediated by ClACS7 in the primordium of the ovary can prevent the development of stamens in hermaphroditic flowers[36]. In this study, the expression levels of ZXG1555 and COS for ClACS7 correspond to this and suggest that it performs the same function.
ClACS7 not only affects the sex differentiation of bisexual flowers in watermelon, but is also associated with a decrease in fruit yield[53]. This gene also influences the longitudinal and transverse growth rates of the ovary and fruit, which means that the fruit from hermaphroditic plants are more round than those from monoecious plants. Therefore, ClACS7 plays a multifaceted regulatory role in the complete development of the pistil and the early development of the fruit. This phenomenon can be traced back to 1928 when Rosa et al. discovered that the fruits produced by hermaphroditic plants were rounder[54]. Poole & Grimball also detected genetic correlations between round fruit and the hermaphroditic trait, as well as between oval fruit and the monoecious plants[55]. Similar multiple effects have also been reported in cucumbers and melons[56−59]. Hence, ClACS7 plays a crucial role in regulating important traits such as pollination, fruit shape, and yield in watermelon. Therefore, in the subsequent studies on fruit shape or yield of watermelon, different types of watermelon varieties with sex differentiation can be utilized, without the intervention of artificial pollination. This will enable a more thorough investigation into the fruit shape and yield of watermelon.
-
This study precisely located the a locus of bisexual flowers in watermelon. The gene controlling andromonoecious is located in an 82.10 kb region on chromosome 3. The analysis of parental sequences and qRT-PCR indicated that Cla97C03G066110 (ClACS7) is the best candidate gene for a locus.
This work was supported by the National Natural Science Foundation of China (Grant Nos 32472747 and U23A20208), the Talent Introduction Project of Northeast Agricultural University (Grant No. 24YJQA02) and the China Agriculture Research System of MOF and MARA (Grant No. CARS-25).
-
The authors confirm their contributions to the paper as follows: study conception and design: Luan F, Liu S; data collection: Fang X, Zhang J, Xu Z, Xu R; analysis and interpretation of results: Guan Y, Wang M, Wang X, Yang Z, Dai Z; draft manuscript preparation: Fang X, Luan F, Liu S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest. Authors Zhongzhou Yang and Zuyun Dai are affiliated with Anhui Jianghuai Horticulture Technology Co., Ltd, which contributed to the analysis and interpretation of results. The company had no commercial or financial interest related to the content of this manuscript.
-
accompanies this paper online at: https://doi.org/10.48130/vegres-0026-0009.
-
# Authors contributed equally: Xufeng Fang, Jing Zhang, Zongwei Xu
- Supplementary Table S1 Primers sequence for QTL analysis and fine mapping processes (97103 v2).
- Supplementary Table S2 Primer sequences used for qRT-PCR.
- Supplementary Table S3 Watermelon accessions used in this study.
- Supplementary Table S4 Information on five predicted genes in the fine mapping region (97103 v2).
- Supplementary Table S5 Information of KASP primers for SNP29,539,131.
- Supplementary Fig. S1 Conserved domain identification for the ClACS7 protein sequence based on NCBI.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fang X, Zhang J, Xu Z, Xu R, Guan Y, et al. 2026. Fine mapping of ClACS7 gene regulating bisexual flowers in watermelon (Citrullus lanatus L.). Vegetable Research 6: e014 doi: 10.48130/vegres-0026-0009 

Fine mapping of ClACS7 gene regulating bisexual flowers in watermelon (Citrullus lanatus L.)
- Received: 04 December 2025
- Revised: 03 February 2026
- Accepted: 22 February 2026
- Published online: 30 April 2026
Abstract: Monoecious and andromonoecious cultivars of watermelon are characterised by the production of male and female flowers, or male and hermaphrodite flowers, respectively. Although the gene responsible for andromonoecy, designated as the a locus, has been identified, its fine-scale genetic mapping remains incomplete. In this study, we built upon our initial mapping of the a locus by performing QTL analysis on F2 populations from spring 2021 (187 plants), and autumn 2021 (137 plants), revealing a stable preliminary region for the a locus on Chr3: 29,252,650–30,021,268 bp. By expanding the ZXG1555 × COS–F2 population to 710 plants, we fine-mapped the a locus to an 82.10 kb interval (Chr3: 29,488,876–29,570,980 bp), which contains five candidate genes. Sequence comparison and transcriptional expression analysis indicated that ClACS7, which encodes 1-aminocyclopropane-1-carboxylate synthase 7, is the most probable candidate gene for this locus. Furthermore, we genotyped 32 natural watermelon populations (2 andromonoecious and 30 monoecious) using the molecular marker Chr03_29539131, which is closely linked to the andromonoecious trait. The results demonstrate that Chr03_29539131 can be effectively used for screening andromonoecy in watermelon and for molecular marker-assisted breeding.
-
Key words:
- Watermelon /
- ClACS7 /
- Sex differentiation /
- Fine mapping