








-
Medicinal plants have served as a cornerstone of human health, from ancient traditional systems such as Traditional Chinese Medicine (TCM) to contemporary pharmacology. Approximately 10% of all vascular plant species are used for therapeutic purposes, with their efficacy derived largely from bioactive secondary metabolites that form the basis of many modern pharmaceuticals[1−6]. However, maintaining a stable and sustainable supply of these biologically derived resources has become increasingly challenging under the combined pressures of climate change, habitat degradation, and volatile global markets[7−9].
Few crops illustrate this convergence of opportunity and vulnerability as clearly as Artemisia annua L.—an ancient medicinal plant transformed into a globally strategic pharmaceutical commodity. A member of the Asteraceae family, A. annua is the only commercial source of artemisinin, the key active ingredient in artemisinin-based combination therapies (ACTs) for malaria treatment[10−12]. Artemisinin is synthesized and stored in glandular trichomes through a complex biochemical pathway that is tightly regulated by genetic, developmental, and environmental factors[13−16]. Its discovery marked a turning point in malaria control, enabling effective treatment of drug-resistant Plasmodium strains and saving millions of lives worldwide[17−21].
Despite its medical importance, the global artemisinin supply chain remains structurally fragile. Production is characterized by fluctuating yields, pronounced price volatility, and agronomic constraints that undermine the reliability of raw material availability for pharmaceutical manufacturers[22−28]. The major risks and structural constraints underlying this volatility—spanning agronomic production, supply chain coordination, and market dynamics—are summarized in Table 1, underscoring the need for a more integrated and resilient production framework. Additionally, geographic variations, such as differences in day length, temperature, and climate, also influence the timing of artemisinin accumulation relative to flowering, requiring region-specific cultivation strategies. A comparative overview of how these environmental drivers shape phenological development and define optimal harvest windows across climate zones is provided in Table 2. Meeting an estimated annual demand of 220–250 metric tons, therefore, presents a critical challenge: how to expand cultivation without exacerbating ecological degradation, genetic erosion, and socio-economic precarity among farming communities[29−31].
Table 1. Key challenges and bottlenecks in the global artemisinin supply chain.
Category Key challenges and risk factors Agronomic and environmental • Land-use competition: Cultivation of A. annua competes with staple food crops for arable land, raising food security concerns.
• Climate change impacts: Increased frequency of extreme weather events (droughts, floods) threatens crop viability and yield stability.
• Geographic constraints: Photoperiod sensitivity limits high-yield cultivation in tropical regions, which bear the highest malaria burden.Genetic and biological • Inherent genetic variability: High heterozygosity in wild and cultivated A. annua leads to inconsistent artemisinin content (0–1.1%).
• Cross-pollination: Gene flow from low-yielding wild Artemisia species can dilute the genetic quality of elite cultivars.
• Pest and disease pressure: While generally robust, monocultures are susceptible to emergent pathogen and pest outbreaks.Post-harvest and processing • Critical harvest timing: Artemisinin content peaks within a narrow window before flowering, making precise timing essential to avoid significant losses.
• Post-harvest degradation: Improper drying, handling, and storage conditions lead to the rapid degradation of artemisinin.
• Extraction inefficiencies: Extraction and purification processes can be costly and result in loss of the final product.Economic and supply chain • Price volatility: A historic 'boom-and-bust' cycle, driven by fluctuating demand forecasts and speculative production, creates extreme financial risk for farmers.
• High cost of alternatives: Semi-synthetic artemisinin production remains expensive and complex, limiting its ability to stabilize the market.
• Logistical bottlenecks: Inadequate infrastructure in many production regions hinders the transport and processing of raw materials.Table 2. Environmental and climatic impacts on A. annua phenology and artemisinin accumulation.
Geographic/climate zone Key environmental drivers Observed phenological response Implication for artemisinin yield Example regions
& key citationsTemperate
(e.g., Northern China)• Long summer day lengths
(>14 h).
• Distinct seasonal temperature shifts.• Prolonged vegetative growth phase.
• Flowering is delayed until late summer/autumn as day length shortens.• High potential: Long vegetative period allows for maximum biomass and artemisinin accumulation before the onset of flowering. [25,32] Subtropical
(e.g., Northern Vietnam, Southern Brazil)• Moderate day lengths.
• High humidity and rainfall.
• Less pronounced temperature seasonality.• Vegetative phase is shorter than in temperate zones.
• Flowering time is sensitive to both photoperiod and temperature cues.• Moderate to high potential: Yield depends heavily on using late-flowering cultivars specifically bred for these conditions. [33,34] Tropical/equatorial
(e.g., Kenya, Uganda, Madagascar)• Consistently short-day lengths (~12 h).
• High and stable temperatures.
• Bimodal rainfall patterns.• Rapid induction of flowering, often prematurely.
• Significantly curtailed vegetative growth and biomass accumulation.• Low potential (for non-adapted varieties): Standard cultivars flower too early, resulting in very low yields. Requires day-neutral or highly adapted hybrids. [35,36] High-altitude tropical
(e.g., East African Highlands)• Short day lengths (~12 h).
• Cooler night temperatures.• Cooler temperatures can partially inhibit or delay the flowering signal despite short days. • Moderate potential: Altitude can mitigate some effects of the tropical photoperiod, making cultivation more viable than in tropical lowlands. [35,37] Conventional strategies centered on large-scale monoculture expansion are increasingly untenable. Such systems are associated with soil degradation, biodiversity loss, heightened pest and disease pressure, and long-term declines in agroecosystem resilience—particularly in tropical and subtropical regions that simultaneously bear the highest malaria burden and host significant biodiversity[38]. Addressing artemisinin supply instability thus requires a paradigm shift, treating A. annua not as an isolated industrial crop, but as an integrated component of diversified, resilient agroecosystems.
This review responds to that need by proposing an integrated framework for the sustainable expansion of A. annua cultivation. Drawing on evidence from agronomy, ecology, biotechnology, artificial intelligence, and socio-economic governance, the framework seeks to enhance artemisinin yield stability while simultaneously conserving agrobiodiversity and improving farmer livelihoods. Rather than prioritizing yield maximization alone, the approach emphasizes system-level resilience, ecological buffering, and equitable value distribution. In doing so, it provides a holistic roadmap for medicinal plant production that aligns public health imperatives with global goals for biodiversity conservation, climate adaptation, and inclusive rural development.
-
The cultivation of A. annua has been the subject of extensive research, focusing primarily on boosting artemisinin content through genetic improvement and optimizing agronomic inputs[10,11,39,40]. Early research focused on identifying high-yielding cultivars and defining basic parameters for fertilization, irrigation, and plant density[39,40]. More recent studies have delved into the molecular biology of the artemisinin biosynthesis pathway, exploring metabolic engineering in various hosts like Saccharomyces cerevisiae and Nicotiana benthamiana as alternatives to agricultural production[12]. However, these biotechnological approaches have not yet achieved commercial scalability, leaving field cultivation as the only viable source of artemisinin[12].
Simultaneously, a separate body of work has explored the supply chain dynamics, highlighting challenges such as price volatility, post-harvest losses, and the economic risks faced by smallholder farmers[25−27,29,30]. More recently, the advent of precision agriculture has introduced the potential of artificial intelligence (AI), internet of things (IoT), and remote sensing to optimize crop management[41]. Despite this wealth of research, a significant gap remains: the lack of an integrated approach that connects these domains within a framework of agrobiodiversity. Most studies focus on a single aspect—yield maximization, molecular biology, or economic analysis—without systematically considering the ecological trade-offs or synergistic benefits of different strategies. The impact of large-scale A. annua cultivation on local biodiversity, and conversely, how enhanced biodiversity can support stable production, is rarely the central theme. This gap is summarized in Table 3, which synthesizes disciplinary paradigms in A. annua research and their associated systemic gaps. The table provides a comparative analysis of dominant research approaches for enhancing artemisinin production by outlining their primary objectives, representative methodologies, and inherent limitations. Collectively, this synthesis reveals a persistent emphasis on productivity within disciplinary silos, with limited integration of ecological or socio-economic dimensions. The analysis underscores a critical and largely unaddressed need for an integrated framework that places agrobiodiversity and ecosystem resilience at the core of a sustainable artemisinin supply chain.
Table 3. Comparative analysis of research paradigms and identified systemic gaps in A. annua studies.
Research area Dominant paradigm/
primary goalRepresentative methodologies and approaches Systemic limitation (identified gap) Genetics and breeding To engineer elite, high-yield cultivars with superior artemisinin content. • Development of detailed phenological scales to synchronize flowering for controlled cross-pollination[34].
• Quantitative Trait Loci (QTL) mapping via deep transcriptome sequencing for marker-assisted selection[42].
• Assessment of heritability and use of mass selection to breed high-content lines and F1 hybrids[25].Prioritizes the creation of uniform, specialized hybrids for industrial monocultures, overlooking the conservation of in situ genetic diversity and the role of locally adapted landraces in building resilient farming systems. Biotechnology and metabolic engineering To create alternative, non-agricultural industrial production platforms and genetically enhance the plant. • Engineering of novel transcription factors (e.g., AaMYB121) to boost biosynthetic gene expression[43].
• Co-overexpression of multiple biosynthetic enzymes in transgenic A. annua to increase artemisinin levels[15].
• Application of nanobiotechnology (e.g., graphene, iron oxide NPs) as elicitors to increase yield[44,45].
• Engineering of transcription factors (e.g., AabZIP1) to link yield with stress tolerance[46].
• Use of external biostimulants (e.g., strigolactones, seaweed extract) to enhance growth and yield[47−49].
• Leveraging beneficial microbes (e.g., Trichoderma, endophytes) to improve plant growth and soil fertility[50,51].
• Heterologous expression of artemisinic acid or its precursors in microbes (S. cerevisiae, E. coli) and subsequent chemical conversion[52−57].
• Engineering alternative plants (e.g., tobacco) as biofactories for drug production and oral delivery[58].Inherently decouples drug production from agriculture or frames biotechnology as a tool for maximizing yield in a single species. This approach sidesteps the opportunity to build sustainable rural economies, enhance farm-level agrobiodiversity, and foster ecological resilience. Agronomy and cultivation sciences To maximize biomass and artemisinin yield per unit area through optimized physical inputs and practices. • Evaluation of planting density effects on biomass and allelopathic potential[59].
• Assessment of polyploid accession performance and stability in new tropical agroecologies[60].
• Comparative analysis of growth and artemisinin content across diverse agro-ecological zones[61].
• Optimization of nitrogen fertilizer application rates to maximize leaf biomass[62].
• Development of physical and chemical seed treatments to improve germination rates[63].
• Investigation of nitrogen's physiological effect on trichome density and artemisinin concentration[64].
• Identification of suitable hybrid lines and management practices for cultivation in the humid tropics[65].
• Determination of optimal nitrogen and potassium nutrition to balance biomass with artemisinin concentration[66].
• Proposal of Good Agricultural Practices (GAPs) for large-scale monoculture production[39].Frames sustainability primarily as resource-use efficiency for a single crop. This input-centric model is disconnected from broader farm-level ecology, long-term soil health, and the economic risks farmers face from input dependency. AI and technology To enhance the efficiency and precision of managing simplified, large-scale monoculture systems. • Using machine learning (ML) for disease detection and yield prediction, including analysis of adoption barriers[67].
• Application of ML with remote sensing for precision input management and crop monitoring[68].
• Integration of AI with IoT, unmanned aerial vehicles (UAVs), and sensors for automated and data-driven farm management[69,70].
• Development of smart crop management systems to optimize irrigation, fertilization, and pest control[71].
• Systematic reviews of ML algorithms (e.g., Support Vector Machine [SVM], Random Forest) and remote sensing data types in precision agriculture[72].
• Use of deep learning with IoT for predictive analytics in crop management[73].
• Application of information and communication technology (ICT) and wireless sensor networks for predictive decision-making and enhancing resource efficiency[74,75].Applies advanced technology to solve problems within the conventional production paradigm, rather than using it to design, model, or manage complex, biodiverse agroecosystems that are inherently more resilient and less input-dependent. Supply chain and economic analysis To mitigate market instability through downstream financial, logistical, and policy interventions. • Supply chain mapping and analysis of high-value crops to identify constraints and scale-up potential[76].
• Value chain analysis of non-timber forest products to map actors, income distribution, and challenges[77].
• High-level review of industrial biotechnology's role in developing sustainable bio-based economies[78].
• Economic modeling of the Artemisia supply chain to analyze the impact of support prices and semi-synthetic supply on volatility[79].
• Analysis of market volatility and proposal of financial mechanisms like buffer stocks and risk pooling[80].
• Examination of the downstream pharmaceutical supply chain and its logistical disruptions at a national level[81].Treats the agricultural source as an abstract 'black box' of production. This approach fails to connect supply chain stability directly to the ecological stability and agrobiodiversity of the production landscapes, where resilience can be built from the ground up. While our approach aligns with the principles of broader sustainable agriculture frameworks, such as the food and agriculture organization's (FAO's) 'Save and Grow' model, which emphasizes ecosystem-based farming, or the consultative group on international agricultural research’s (CGIAR's) focus on sustainable intensification for food security, it offers a distinct, specialized focus[82,83]. These established models provide foundational principles for general agriculture but often lack specific guidance for high-value, non-food medicinal plants like A. annua, which have unique value chains and ecological considerations. The proposed framework in this literature-based review is novel in its explicit integration of three domains typically addressed in isolation: (1) advanced agronomy with in situ genetic diversity conservation; (2) the application of AI not just for precision input management but for monitoring agrobiodiversity metrics; and (3) a socio-economic governance model (PPFP) tailored to the volatile medicinal plant market. Therefore, this framework is among the first to systematically link AI-driven monitoring with genetic diversity conservation and socio-economic enablers, specifically for a medicinal plant commodity.
-
The analytical framework presented here is based on a systematic literature review. The synthesis was conducted following the principles of the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines to ensure a comprehensive review. A search of prominent academic databases, including Scopus, Web of Science, PubMed, and Google Scholar, was conducted for the period 1995–2025. Search terms included combinations of 'Artemisia annua L.', 'artemisinin', 'sustainable agriculture', 'agrobiodiversity', 'multi-cropping', 'intercropping', 'genetic diversity', 'smallholder farmers', 'precision agriculture', 'phenology', and 'AI in agriculture'. Articles were selected for inclusion if they addressed agronomic, ecological, genetic, or socio-economic aspects of A. annua cultivation. Studies focusing purely on biochemical synthesis or clinical trials were excluded unless they provided context on supply chain demands. This structured approach allowed for the integration of findings from disparate fields into the cohesive strategic model that follows.
Core analytical questions
-
To address the identified gaps, this review employs a systematic literature synthesis and analytical framework structured to answer key questions critical for sustainable expansion. This methodology moves beyond a simple review by integrating findings from diverse fields to build a cohesive strategic model. The analysis is guided by the following core questions:
1. How can A. annua cultivation be adapted across multiple latitudes without compromising genetic diversity or ecological stability?
2. What specific sustainable agronomic practices are most effective for enhancing both artemisinin yield and local agrobiodiversity (e.g., soil health, beneficial insects)?
3. How can AI-driven technologies and intelligent farming systems be applied not just to optimize yield, but also to monitor and enhance sustainability metrics?
4. How does the integration of A. annua into diversified cropping systems (e.g., with staple crops) influence farm income, food security, and the conservation of agrobiodiversity?
5. What are the primary economic and ecological impacts of large-scale A. annua cultivation, and what governance models (e.g., Public-Private-Farmer Partnerships) can mitigate risks and ensure equitable benefits?
By systematically addressing these questions, a comprehensive framework is constructed that is scientifically grounded, ecologically conscious, and practically relevant for stakeholders across the artemisinin value chain.
-
Building on the limitations identified in prevailing productivity-driven approaches, this study advances a systems-based framework for A. annua cultivation that explicitly integrates agronomic, ecological, technological, and governance dimensions. Rather than treating yield optimization as an isolated objective, the framework is structured around four interdependent pillars: (1) biodiversity-centric agronomic practices that enhance productivity while strengthening ecosystem function; (2) climate-adaptive, multi-latitude cultivation strategies that buffer environmental variability; (3) intelligent farming approaches that enable precision management and resource efficiency; and (4) robust ecological and socio-economic governance mechanisms that support long-term resilience and equitable value distribution across the supply chain.
Biodiversity-centric agronomic practices
-
The foundation of a sustainable production framework rests on agronomic practices that enhance yield while simultaneously enriching the agroecosystem, requiring a shift from conventional input optimization toward a more holistic management approach (Fig. 1). As illustrated in Fig. 1, A. annua cultivation typically follows a sequential annual cycle, beginning with seed stock retained from the previous harvest. The cultivation phase involves seed germination and nursery-based plantlet establishment, followed by transplantation into prepared fields where plants undergo maturation supported by essential field management practices such as weeding. Harvesting is strategically timed to coincide with the flowering stage—approximately two months post-transplantation—when artemisinin accumulation in leaf biomass reaches its peak. Following harvest, plant material is dried, and artemisinin-rich leaves are separated from stems and directed to downstream processing steps, including extraction and purification, to yield crystalline artemisinin.
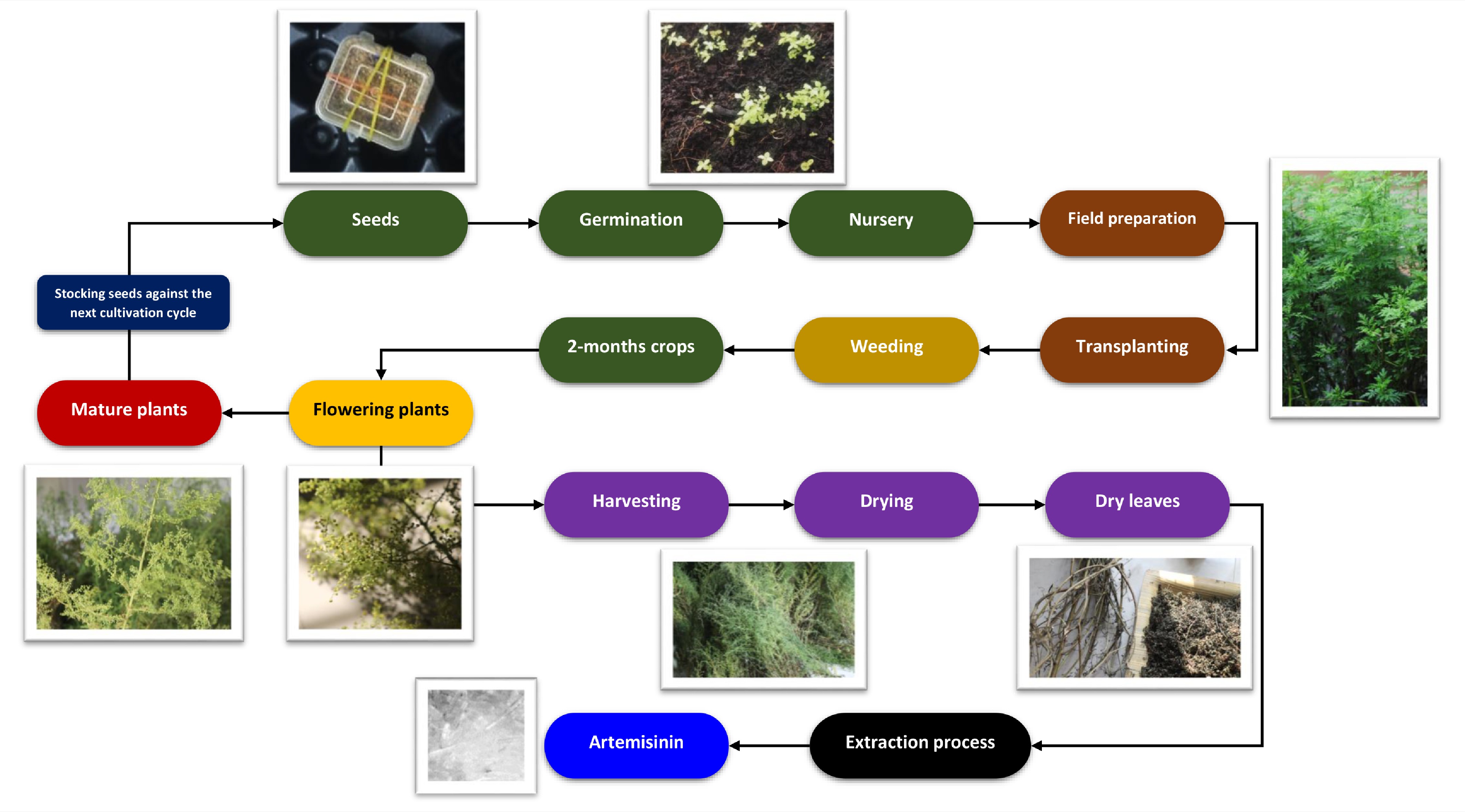
Figure 1.
The annual cultivation and production cycle of A. annua, from nursery establishment to crystalline artemisinin extraction.
Although wild A. annua populations typically exhibit low and highly variable artemisinin content (0%–0.9% dry weight), sustained breeding and selection efforts have led to the development of late-flowering hybrids with substantially improved performance. Notably, cultivars developed in Brazil demonstrate markedly enhanced biomass production (> 30 t·ha−1 fresh leaves) alongside elevated artemisinin concentrations, reaching up to 1.0975%[39,40,84−86].
The following management practices are designed to ensure that improved A. annua cultivars achieve their full productive potential while maintaining long-term agroecosystem sustainability. Effective soil health management is foundational, as A. annua can tolerate a broad soil pH range (5.0–8.0), although certain genotypes exhibit sensitivity below pH 5.5. Rather than relying exclusively on synthetic fertilizers, an integrated soil fertility management strategy is recommended. While balanced macronutrient applications (approximately 100 kg·ha−1 each of N, P, and K) have been shown to significantly enhance dry biomass production, micronutrient supplementation has not demonstrated a consistent effect on artemisinin biosynthesis[40]. These findings support prioritizing practices that build soil organic matter—such as compost application, cover cropping, and crop rotation—which promote gradual nutrient release and improve soil structure and resilience.
Crop establishment strategies should be adapted to local production contexts, particularly in smallholder systems where direct broadcast seeding is often more cost-effective than transplanting. To achieve uniform seed distribution, seeds may be mixed with an inert carrier such as sand and sown at the onset of the rainy season into finely tilled soil at an optimal depth of approximately 5 mm, followed by immediate irrigation to prevent desiccation[39]. Maintaining an appropriate planting density (typically 1–2.5 plants m−2) is critical for balancing resource use efficiency, canopy development, and weed suppression.
Water management plays a dual role in supporting vegetative growth and modulating secondary metabolite accumulation. Adequate irrigation is particularly important during early growth stages; however, evidence indicates that inducing moderate water stress shortly before harvest can enhance artemisinin concentration in leaf tissues[40]. Weed pressure can be effectively managed during early establishment through inter-row cultivation and mulching, while canopy closure at later stages suppresses weeds naturally through shading and the allelopathic properties of A. annua, thereby reducing reliance on chemical herbicides[87,88].
Harvesting and post-harvest handling are critical determinants of final artemisinin yield and quality. Harvest timing must balance total biomass accumulation with peak artemisinin concentration, which occurs primarily in the leaves and inflorescences. In regions with favorable climatic conditions, multiple harvests per season may be achieved by cutting plants back to stimulate regrowth, substantially increasing annual productivity. Post-harvest processing protocols, summarized schematically in Fig. 2, are essential for preserving artemisinin content. Optimal practice involves sun-drying harvested biomass in the field for no longer than one week to minimize leaf loss, followed by shade drying. Leaves are then separated from stems through threshing and sieving (typically using 5 and 3 mm mesh sizes). Fine grinding is avoided, as artemisinin is stored in fragile glandular trichomes on the leaf surface that are easily damaged. For storage before extraction, maintaining a relative humidity of 12%–13% is critical to prevent degradation of the active compound[40].
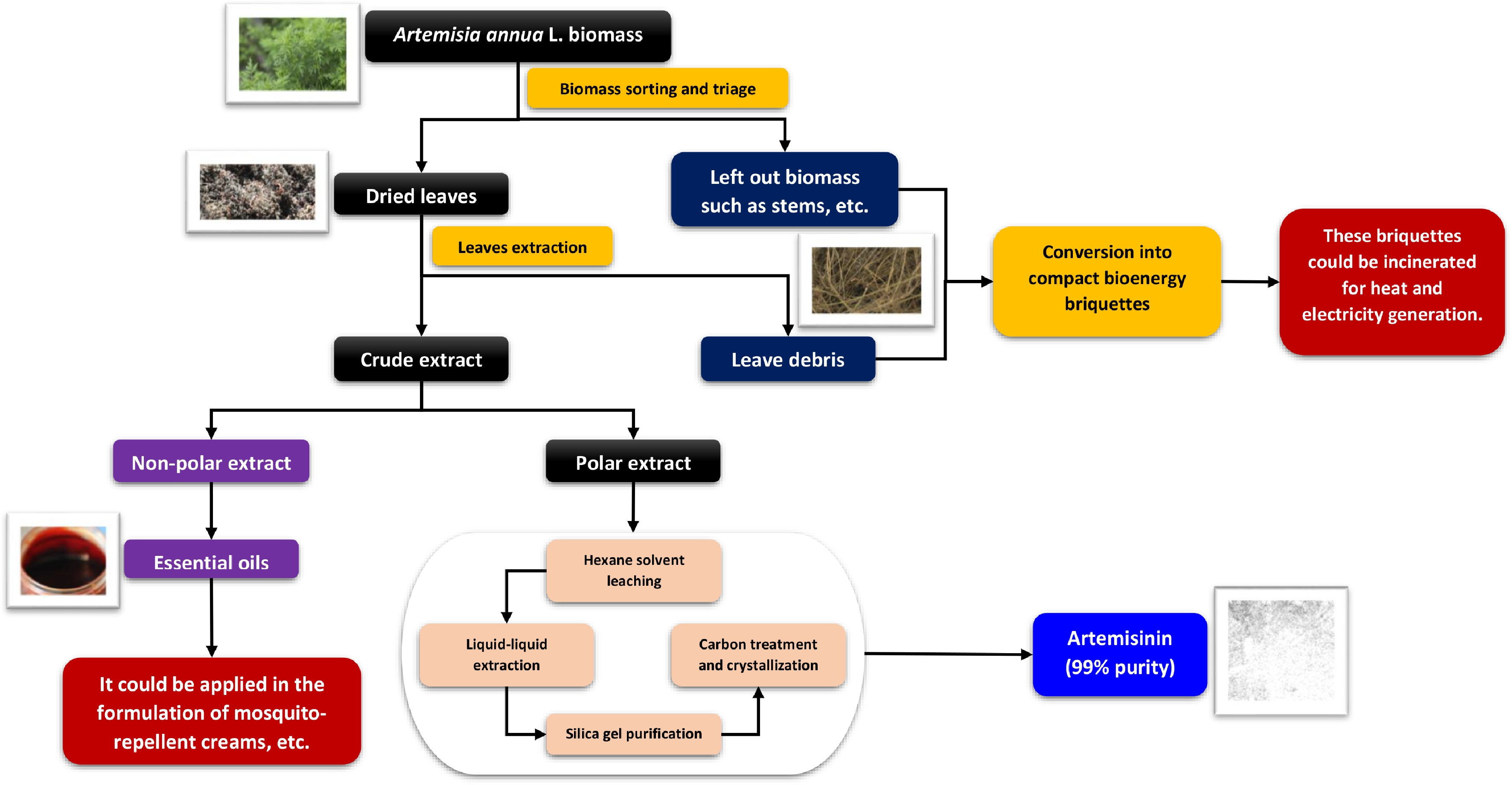
Figure 2.
Schematic of an integrated biorefinery for A. annua biomass valorization and zero-waste processing.
Figure 2 further illustrates a zero-waste biorefinery pathway designed to maximize value recovery from harvested A. annua biomass. The process separates dried material into two complementary streams. The primary stream focuses on high-value leaf processing, in which dried leaves are extracted to yield a crude product that is fractionated into non-polar and polar components. The non-polar fraction provides essential oils suitable for commercial applications such as mosquito-repellent formulations, while the polar fraction undergoes a multi-step purification cascade—including hexane leaching, liquid–liquid extraction, silica gel chromatography, and carbon treatment—culminating in the crystallization of high-purity (> 99%) artemisinin. The secondary stream valorizes residual biomass, including stems and leaf debris, by converting it into compacted bioenergy briquettes, thereby supporting renewable energy generation and reinforcing the circular, zero-waste nature of the production system.
Multi-latitude cultivation: balancing productivity and genetic diversity
-
A. annua is a short-day plant, meaning the transition to flowering is triggered by decreasing day length. This photoperiod sensitivity is a major barrier to cultivation in tropical regions with shorter day lengths (particularly during the summer season), as premature flowering curtails vegetative growth and biomass accumulation[89,90]. To expand cultivation across multiple latitudes, a multifaceted strategy focusing on genetic potential and agronomic management is essential.
The genetic diversity within A. annua offers significant opportunities for adaptation. For instance, detailed phenological studies by Marchese et al.[34] revealed critical differences in flowering triggers between genotypes. While a Chinese genotype required only a short day to initiate flowering, a Vietnamese genotype needed the dual stimuli of both short days and low-to-moderate temperatures. This highlights that adaptation is not just about day length but also about temperature interactions, a crucial factor in climate-resilient breeding.
Furthermore, that same study challenged the long-held belief that artemisinin concentration peaks exclusively at flowering. It demonstrated that plants maintained in a vegetative state under long photoperiods exhibited higher densities of glandular trichomes and, consequently, greater artemisinin concentrations than plants induced to flower under short days[34]. This finding opens up a key agronomic strategy: manipulating the cultivation calendar to prolong the vegetative phase. For example, by delaying the transplantation of seedlings to the field until after the critical inductive photoperiod has passed, farmers can prevent early flowering and maximize biomass and artemisinin yield. This level of phenological control is fundamental to optimizing the crop for different latitudes.
Building on this understanding, breeding programs must focus on developing late-flowering or day-neutral hybrid varieties that are less sensitive to photoperiod. Advanced techniques such as genome selection, speed breeding, and gene editing can accelerate the development of cultivars with fine-tuned photoperiod sensitivity[91,92]. The success of Brazilian hybrids, which can produce over 30 t·ha−1 of fresh leaves, demonstrates the potential of this approach[39]. Importantly, these breeding programs must draw from a wide genetic base, including wild populations, to maintain genetic diversity and explore other adaptive traits, such as a potential gradual day-length sensing mechanism that could allow for more dynamic responses to seasonal changes[93,94].
Integrating A. annua into existing multi-cropping systems is also a cornerstone of sustainable expansion[95] (Box 1). In regions with bimodal rainfall or structured irrigation scheduling, A. annua can be grown in rotation or intercropped with staple food crops like maize, legumes, or cassava. This diversification enhances agrobiodiversity by creating more complex habitats for beneficial insects and soil microbes. It also improves food security, provides farmers with diversified income streams, and reduces the economic risk associated with relying on a single commodity crop. For example, planting nitrogen-fixing legumes alongside A. annua can naturally enrich the soil, reducing the need for synthetic fertilizers.
Box 1: Case study - Cultivation of Artemisia annua L. in India
Artemisinin accumulation in Artemisia annua L. is a complex, multifactorial process governed by genetic, environmental, developmental, agronomic, and stress-related factors. Biosynthesis of the compound occurs in glandular trichomes and is tightly regulated by key genes, including AaADS, AaCYP71AV1, AaALDH1, and AaDBR. Key environmental parameters—such as light intensity, temperature, and nutrient availability—along with agronomic practices like nitrogen optimization and soil conditioning, significantly modulate artemisinin synthesis and production efficiency. Moreover, biotic and abiotic stresses can trigger enhanced secondary metabolite production as part of the plant's defense response, suggesting that stress-induction pathways could be exploited to maximize artemisinin yields.
The adaptability of A. annua to diverse agro-climatic conditions is exemplified by the development and successful establishment of the 'Jeevanraksha' cultivar in India's Indo-Gangetic Plain[96]. Studies by Kumar et al.[96] demonstrated that artemisinin biosynthesis initiates in March and April, peaks from May to June, and remains elevated throughout the rainy season (July–September) (Table 4). Over 90% of the compound accumulates in leaves and fine stems during the vegetative phase. Notably, the roots do not store artemisinin, reinforcing the importance of optimal harvest timing. Multi-harvesting (ratooning) techniques, involving three to four sequential harvests over ≥ 30 weeks, yielded 44.1 ± 14.2 kg·ha−1 to 74.2 ± 15.6 kg·ha−1 of artemisinin, significantly surpassing the previously reported maximum of 25 kg·ha−1 [97]. Early planting, coupled with multiple harvests, maximized leaf biomass while minimizing excessive stem growth, thereby optimizing the overall artemisinin yield.
Table 4. Phenological and agronomic timeline for Artemisia annua L. cv. 'Jeevanraksha' in subtropical India.
Timeline (month) Key phenological stage Critical agronomic activities Artemisinin dynamics and key considerations Dec–Jan Nursery stage • Prepare nursery beds.
• Sow seeds under protected conditions.• No significant artemisinin production.
• Focus is on ensuring high germination rates and healthy seedling development.Feb–Mar Seedling and establishment • Transplant healthy seedlings to the main field.
• Conduct initial irrigation and gap-filling.
• Implement early-stage weed management.• Artemisinin biosynthesis begins as temperatures rise, primarily in young leaves.
• The flexible transplantation window (Feb-Aug) can be used to manage plant density and timing.Apr–Jun Rapid vegetative growth • Apply targeted fertilizers to drive robust biomass accumulation.
• Ensure consistent moisture through irrigation.
• Monitor for pests and diseases.• Artemisinin content steadily increases. Younger, upper leaves consistently show higher concentrations than older, lower leaves.
• Maximizing leaf biomass during this stage is critical for final yield.Jul–Sep Peak vegetative and pre-flowering • Optimal harvest window: Begin harvesting leaves at peak artemisinin content.
• Manage water drainage during the monsoon season.
• Cease nutrient application pre-harvest.• Peak artemisinin accumulation: Artemisinin levels in leaves reach their maximum.
• Delaying harvest into this period ensures both high biomass and high potency.
• Over 90% of total artemisinin is located in leaves and fine stems.Sep–Oct Full flowering stage • Continue harvesting if targeting floral parts.
• Reduce irrigation.
• Prepare for seed collection from selected plants.• Leaf artemisinin content begins to decline as it is partially translocated to the flowering heads (capitula).
• Capitula become a significant source of artemisinin (up to 40% of total).Nov–Dec Seed maturation and senescence • Harvest mature seeds from unharvested plants.
• Clear remaining plant biomass from the field.• Artemisinin content in vegetative parts drops significantly.
• Seeds contain negligible amounts of artemisinin.Post-harvest Drying and processing • Summer/winter harvest: Shade-dry leaves to preserve artemisinin.
• Rainy season harvest: Use aerated drying chambers or forced-air systems to prevent fungal growth and artemisinin degradation.• Proper drying is critical. Rapid, high-temperature drying can cause significant artemisinin loss.
• Shade drying is the preferred method to maintain quality.Post-harvest handling is critical for preserving artemisinin content. Shade drying proved effective during summer and winter, whereas specialized techniques were required during the rainy season to prevent degradation. Efficient post-harvest logistics—including drying, separation, and moisture-proof storage—are essential for commercial extraction and supply chain stability. Despite these advancements, Kumar et al.[96] identified persistent inefficiencies in multi-harvesting methods and proposed several strategies to further boost yields, including genetic improvements, enhancing the leaf-to-stem ratio, and optimizing harvest schedules.
Beyond its agronomic potential, the 'Jeevanraksha' cultivar demonstrated compatibility with staple crop rotations in subtropical agro-climatic zones. Viable cropping systems, such as Wheat/Brassica/Bengal Gram/Lentil/Potato–A. annua and Rice–Potato–A. annua, effectively balance artemisinin production with food security. Furthermore, contract farming models that link cultivators with manufacturers offer a scalable pathway to ensure consistent seasonal production while improving farmer profitability.
The successful integration of A. annua into multi-cropping systems highlights the potential to expand its cultivation beyond traditional temperate regions. Targeted breeding and optimized agronomic strategies can drive higher artemisinin yields while offering smallholder farmers and large-scale cultivators additional revenue streams. This synergistic approach fosters agricultural resilience, enhances resource efficiency, and stabilizes the global supply of artemisinin—an essential compound in malaria treatment—underscoring the need for sustainable and scalable production frameworks. Table 4 below provides a month-by-month guide for the cultivation of the 'Jeevanraksha' cultivar, outlining the key plant development stages, corresponding agricultural practices, and the dynamics of artemisinin biosynthesis. The timeline is specifically adapted for the subtropical agro-climatic conditions of the Indo-Gangetic plains, providing a framework for optimizing yield and harvest timing. Adapted from data in Kumar et al.[96].
Intelligent farming for enhanced productivity and sustainability
-
Artificial intelligence (AI) and the Internet of Things (IoT) offer transformative tools to optimize Artemisia annua L. cultivation sustainably. By integrating data-driven insights with traditional agronomic knowledge, intelligent farming systems enable a shift from broad-spectrum, calendar-based management to localized, precision-oriented decision-making that enhances productivity while reducing environmental impacts.
At the field level, precision monitoring and input management are central to this transformation. IoT sensors deployed across production systems can continuously monitor soil moisture, nutrient availability, and microclimatic conditions in real time[41]. When integrated into AI-powered decision-support systems, these data streams enable the precise application of water and fertilizers only when and where they are needed, minimizing resource waste, lowering production costs, and reducing environmental damage associated with nutrient runoff. Complementarily, drone-based multispectral imaging allows for early detection of crop stress, diseases, or pest infestations, facilitating targeted interventions and reducing reliance on broad-spectrum pesticide applications[41].
AI also enables predictive analytics for harvesting and yield optimization. Machine learning (ML) models trained on historical datasets that combine plant phenology, weather patterns, and soil characteristics can predict the optimal harvest window for maximizing artemisinin content[41]. This data-driven approach replaces fixed, calendar-based harvesting schedules, ensuring peak phytochemical potency and reducing uncertainty for producers. In addition, yield forecasting models provide more reliable production estimates, supporting supply chain stability and allowing pharmaceutical manufacturers to better anticipate raw material availability[41].
Beyond the farm gate, AI-driven supply chain optimization enhances traceability and quality control throughout the A. annua value chain. By digitally tracking production batches from field to factory, AI systems can verify compliance with quality standards and post-harvest handling protocols, thereby minimizing artemisinin degradation and ensuring consistency of pharmaceutical-grade raw materials[41].
Importantly, the transformative potential of AI extends beyond optimizing inputs for a single crop. Within this integrated framework, intelligent systems can be applied to the design and management of complex, biodiverse agroecosystems. Rather than using ML solely to predict monoculture yields, advanced models can assess the synergistic outcomes of intercropping combinations, identifying plant assemblages that enhance pest suppression, nutrient cycling, and overall system resilience. Furthermore, remote sensing data from drones and satellites, when analyzed with AI, can monitor not only crop health but also broader biodiversity indicators—including habitat complexity, soil organic matter dynamics, and pollinator activity—at a landscape scale. In this way, digital technologies shift from tools that manage simplified systems to instruments that help understand, monitor, and enhance ecological complexity, a core principle of sustainable agrobiodiversity-based production.
Despite these advantages, the adoption of intelligent farming technologies is constrained by structural and economic barriers. High capital costs associated with drones, proprietary software, and dense IoT sensor networks risk creating a 'digital divide' that favors large-scale industrial farms while marginalizing smallholder producers[98,99]. Evidence from emerging markets further highlights a 'technology mismatch', where expensive, proprietary systems are poorly aligned with the financial and infrastructural realities of rural farming communities[100]. Without deliberate intervention, these dynamics threaten to undermine the inclusivity goals of sustainable A. annua cultivation frameworks.
To address this challenge, the framework advocates a strategic shift from land-scale ownership of digital assets toward service-scale and shared-access models[100]. In such systems, digital services—rather than hardware ownership—become the primary unit of access. Cooperative platforms or service providers centralize capital investment and technical expertise while decentralizing benefits to individual farmers, thereby reducing financial risk and improving scalability.
Central to this approach is the concept of frugal innovation, defined as the deployment of low-cost, high-impact technologies tailored to resource-constrained environments[101]. Scalable alternatives that democratize data-driven farming include SMS-based advisory services delivering timely alerts on weather forecasts, pest outbreaks, and optimal planting or harvesting windows[102,103]; community-owned automated weather stations that provide localized, high-resolution climatic data; and AI-enabled mobile applications that use smartphone cameras to diagnose pests and diseases and offer integrated pest management recommendations. In parallel, farmer cooperatives can collectively invest in and share higher-cost technologies, such as drones, making precision agriculture tools accessible without requiring individual ownership[104]. These tiered technology pathways—ranging from high-end remote sensing to frugal, mobile-based solutions—ensure that the benefits of intelligent farming can be realized across diverse economic contexts. Table 5 below summarizes a tiered framework for deploying intelligent farming technologies in A. annua production systems, ranging from high-end remote sensing to frugal, mobile-based solutions and service-oriented models. The framework emphasizes scalability, inclusivity, and alignment with local socio-economic conditions, aiming to minimize digital exclusion while maximizing agronomic efficiency, artemisinin yield optimization, and agroecosystem resilience[98,105].
Table 5. A tiered approach to intelligent farming technologies for A. annua.
Technology tier Core technologies and tools Functional role in A. annua systems Benefits for smallholder and
mixed-scale producersKey constraints and risks High-end
(capital-intensive)Drone-based multispectral and hyperspectral imaging; satellite remote sensing integrated with AI analytics Landscape-scale detection of crop stress, pest and disease outbreaks, nutrient deficiencies, and spatial variability in biomass and phenology; supports site-specific interventions and biodiversity monitoring Enables early, non-invasive diagnostics and precision interventions; improves targeting of pest and disease control; supports monitoring of agroecosystem heterogeneity and habitat complexity[106]. High capital and maintenance costs; need for skilled operators and data-processing capacity; risk of exclusion of smallholders without cooperative or service-based access[101]. Mid-tier (farm-level precision) In-field IoT sensor networks (soil moisture, temperature, pH, nutrient probes); localized weather sensors linked to decision-support systems Real-time monitoring of soil–plant–atmosphere interactions; optimization of irrigation, fertilization, and harvest timing to maximize artemisinin yield and compliance with Good Agricultural Practices (GAPs) Improves resource-use efficiency; reduces input waste and environmental impacts; enhances yield predictability and quality consistency[107]. Dependence on connectivity and power supply; sensor maintenance and calibration requirements; moderate upfront and recurring costs[108]. Frugal (low-cost, scalable) SMS-based advisory services; Unstructured Supplementary Service Data (USSD) platforms; AI-enabled smartphone diagnostic apps; 'farm hack' or Do-It-You (DIY) sensor kits Dissemination of timely agronomic advice (weather alerts, pest warnings, planting/harvest windows); rapid field-level pest and disease identification using mobile cameras Affordable and scalable access to decision support; leverages widespread mobile phone ownership; suitable for remote and resource-constrained settings[101]. Limited data resolution; reliance on basic digital literacy and mobile network coverage; reduced precision compared with sensor-based systems[99]. Service-based/cooperative Outsourced drone services; shared data platforms; cooperative-owned machinery and analytics services Provision of precision agriculture services without individual asset ownership; aggregation of data for regional forecasting and supply-chain coordination Reduces capital risk for farmers; enables access to advanced technologies; strengthens collective bargaining power and data-driven planning[100]. Requires strong cooperative governance and trust; potential data ownership and privacy concerns; dependency on service availability[100]. Finally, bridging the implementation gap requires recognizing that the transition to intelligent farming is not solely a technical challenge but a socio-technical process. Successful adoption depends on Public–Private–Farmer Partnerships (PPFPs) that can subsidize initial technology costs and invest in capacity-building for local extension agents. 'Learning by doing' approaches[107] can enhance farmer engagement and digital literacy while safeguarding data sovereignty, ensuring that technology serves as a tool for empowerment rather than surveillance or exclusion[105]. When embedded within such inclusive governance structures, intelligent farming systems position the A. annua value chain as a model for how high-value medicinal plant production can be simultaneously technologically advanced, ecologically resilient, and socially equitable.
Ecological and agrobiodiversity considerations
-
The large-scale expansion of A. annua cultivation carries ecological risks that must be proactively managed to ensure long-term sustainability. These risks are most pronounced in simplified production systems, where monocultures can destabilize biodiversity, soil health, and ecosystem services, ultimately undermining productivity and resilience.
Impact on biodiversity and ecological stability
-
Large-scale monocultures of A. annua create a homogenized landscape that reduces local biodiversity by displacing native vegetation and simplifying habitat structure. Such environments support fewer species of insects, birds, and soil organisms, disrupting pollination services for adjacent crops and increasing vulnerability to pest and disease outbreaks[38,109−113]. This heightened susceptibility is well explained by the resource-concentration hypothesis, which posits that the success and spread of specialist pests and pathogens increase with the spatial and temporal continuity of their host plants[114].
Empirical evidence demonstrates that continuous cultivation of A. annua promotes the accumulation of host-specific pathogens, including Erysiphe artemisiae and Puccinia tanaceti, leading to increased disease incidence over time[115]. These biodiversity losses and pathogen pressures underscore the inherent ecological fragility of monocropping systems.
To mitigate these risks, biodiversity-enhancing practices are essential. The incorporation of hedgerows, wildflower strips, and diversified cropping patterns around and within A. annua fields provide habitats for pollinators and natural enemies of pests, strengthening biological control mechanisms and restoring functional diversity at the field and landscape scales.
Biotic stress and microbial imbalance in monocultures
-
Beyond aboveground biodiversity loss, A. annua monocultures exert profound effects on belowground ecological processes. Continuous monocropping disrupts soil microbial stability through both physical simplification and biochemical feedback. Artemisinin, released into the soil at concentrations reaching up to 11.7 mg·kg−1 under field conditions, exhibits dual biochemical effects[116−118]. While its allelopathic properties successfully suppress weeds, its potent antimicrobial activity simultaneously disrupts the soil ecosystem. Specifically, it reduces microbial biomass carbon and nitrogen while suppressing key nutrient-cycling enzymes, including dehydrogenase and urease[116].
The resulting decline in microbial richness and evenness can favor the over-proliferation of specific bacterial taxa, increasing the likelihood of soil-borne disease outbreaks and contributing to the phenomenon commonly described as 'soil sickness'[116]. Artemisinin has also been shown to significantly inhibit the growth and nutrient uptake (nitrogen, phosphorus, and potassium) of ectomycorrhizal fungi, which play a critical role in nutrient mobilization and the health of adjacent forest and agroecosystems[87]. Collectively, these biotic stresses illustrate why monoculture-based A. annua production systems are ecologically risky over the long term.
Impacts on ecosystem services
-
The ecological consequences of A. annua monocultures extend to broader ecosystem services. Annual monocropping systems are associated with increased soil erosion and nutrient leaching, particularly under intensive management[119]. Artemisinin is relatively persistent in soil and potentially leachable, posing risks to aquatic ecosystems where it exhibits toxic effects comparable to commercial herbicides such as atrazine[117]. Moreover, artemisinin acts as a repellent to key soil fauna, including earthworms (Eisenia fetida), whose activity is essential for maintaining soil structure, aggregation, and overall soil quality[117].
Restoring stability through ecological buffering
-
The integrated framework addresses these ecological risks through the principle of ecological buffering, defined as the strategic use of agrobiodiversity to interrupt pest and disease cycles, stabilize soil microbial communities, and restore ecosystem services. Intercropping A. annua with functionally contrasting species, such as soybean (Glycine max), acts as a 'dilution effect' that reduces host visibility and resource continuity for specialist herbivores[120]. In diversified plant communities, volatile organic compounds emitted by non-host species can further enhance indirect plant defenses by attracting predators and parasitoids that suppress pest populations below economic thresholds[114,120].
Belowground, ecological buffering is achieved through crop rotations with species such as Sesbania and the application of organic amendments, including vermicompost. These practices have been shown to restore soil microbial diversity, increase operational taxonomic unit (OTU) richness and evenness, and recover enzymatic activities critical for nutrient cycling, thereby counteracting the microbial degradation associated with continuous A. annua monocropping[121] (Table 6).
Table 6. Ecosystem disservices of A. annua monocultures versus buffering benefits of diversified systems.
Ecosystem parameter Monoculture impact (disservice) Polyculture solution (buffering) Evidence basis Pest vulnerability Rapid spread of specialist pests (e.g., Artemisia aphids) Attraction of natural enemies via diversified plant volatiles [114] Pathogen load Accumulation of E. artemisiae and P. tanaceti Disruption of disease cycles through rotation and intercropping [115] Soil microbiome Reduced microbial richness; dominance of pathogenic taxa Restoration of OTU richness and evenness via organic amendments and crop diversity [116,121] Nutrient cycling Suppressed urease and dehydrogenase activity Recovery of enzymatic activity and nitrogen availability [116,121] Soil fauna Repellence of earthworms and degradation of soil structure Improved habitat quality for beneficial soil fauna and arthropods [117,120] Implications for soil health and nutrient cycling
-
The buffering mechanisms outlined above form the foundation for maintaining long-term soil health in A. annua production systems. By stabilizing microbial communities, enhancing soil fauna activity, and restoring enzymatic processes, agrobiodiversity acts as the primary mechanism through which nutrient cycling, soil structure, and organic matter dynamics are preserved. These processes directly support the soil health outcomes, reinforcing the central role of biodiversity-based buffering in sustaining productive and resilient A. annua agroecosystems[121]. Table 7 below analyzes and compares major cropping systems, evaluating their operational characteristics, primary strategic trade-offs, and potential for enhancing agrobiodiversity. The relevance of each system is specifically assessed for its applicability to the cultivation of A. annua, to balance high artemisinin yield with ecological stewardship and farmer prosperity.
Table 7. A comparative framework of cropping systems for sustainable A. annua cultivation.
Cropping system Description Primary trade-off Biodiversity potential Relevance for Artemisia annua L. cultivation I. Monoculture systems Monocropping
(sole cropping)Cultivation of a single crop species on the same land over successive seasons. Efficiency and scale vs ecological risk − The current industrial standard for maximizing short-term yield, but it degrades soil health and agrobiodiversity, creating long-term vulnerabilities. Ratooning Cultivating a new crop from the regrowth of the previous crop's stumps or roots. Low establishment cost vs declining yield and pest accumulation − Not a viable system. A. annua is an annual plant harvested for its aerial biomass and does not regrow effectively from stumps. II. Polyculture (multi-cropping) systems Sequential cropping Growing two or more crops in succession on the same land within one year. Land use intensity vs climatic dependence + Highly relevant. Allows A. annua to be rotated with a staple food crop (e.g., legumes, short-season cereals), improving food security and breaking pest cycles. Intercropping (simultaneous cropping) Growing two or more crop species concurrently in the same field. Resource synergy vs management complexity ++ A core strategy. Companion planting with legumes (e.g., cowpea) fixes nitrogen, while aromatic herbs (e.g., basil) can provide pest deterrence. Mixed intercropping Crops are grown together without a distinct row arrangement. Maximum resilience vs inability to mechanize +++ Best suited for smallholder, low-input subsistence farms where A. annua is part of a diverse garden of medicinal plants and vegetables. Row intercropping Crops are grown in distinct, alternating rows. Management control vs potential interspecies competition ++ An excellent, practical model. A. annua can be grown in rows alternating with low-growing cash crops like groundnut or beans. Strip Intercropping Crops are grown in wide, multi-row strips, allowing for separate mechanical access. Mechanization vs reduced ecological interaction + A scalable commercial model. Allows farms to balance the efficiencies of mechanization with the soil health benefits of crop rotation and diversity. Relay cropping Sowing a second crop into an established primary crop before it is harvested. Season maximization vs risk of harvest interference ++ A promising intensification strategy. Legumes can be under-sown into a mature A. annua stand to ensure continuous land cover and provide a 'green manure.' Agroforestry Integrating woody perennials (trees, shrubs) with crops and/or livestock. Long-term resilience vs high initial investment +++ A long-term vision. Trees can serve as windbreaks, improve the microclimate, enhance water retention, and provide farmers with diversified, high-value income streams. Alley cropping (hedgerow intercropping) Crops are grown in 'alleys' between rows of trees or shrubs. Nutrient cycling vs labor for hedgerow management +++ A structured agroforestry approach. Planting hedgerows of nitrogen-fixing trees (e.g., Gliricidia sepium) can provide natural fertilizer and mulch for the A. annua grown in the alleys. Biodiversity potential is rated on a scale from negative (−) to highly positive (+++), reflecting the system's capacity to support species richness and ecosystem services. Genetic diversity as a complementary safeguard
-
Finally, ecological buffering at the field and soil levels must be complemented by strategies that protect genetic diversity within A. annua populations. Over-reliance on a narrow set of high-yielding hybrids accelerates genetic erosion, increasing vulnerability to emerging diseases and climate variability[122−128]. A multi-pronged approach—combining community seed banks, in-situ conservation incentives, participatory breeding programs, and strengthened linkages with national and international gene banks—ensures that genetic diversity remains a dynamic resource for adaptation rather than a static archive.
Together, these ecological, soil, and genetic dimensions position agrobiodiversity not as a constraint on intensification, but as the central organizing principle for sustainable A. annua cultivation systems.
Socio-economic enablers and policy recommendations
-
The successful implementation of this integrated framework depends not only on technical feasibility but also on a supportive socio-economic and policy environment. Transitioning from conventional A. annua monoculture toward diversified, agroecological systems requires mechanisms that reduce risk, correct power imbalances, and create durable incentives for farmer participation.
Innovative partnership models are central to this transition, with Public–Private–Farmer Partnerships (PPFPs) providing a structural framework for coordinated action (Fig. 3). Within this model, public institutions support research, extension services, and enabling policies; private-sector actors contribute market access, processing capacity, and long-term purchasing commitments; and farmer cooperatives strengthen collective bargaining power, knowledge exchange, and access to productive resources. Together, these interactions create the institutional conditions for a stable, profitable, and environmentally sustainable artemisinin supply chain. However, while PPFPs establish the architecture for collaboration, their internal governance mechanisms must be intentionally designed to avoid reproducing historical power imbalances and inequities characteristic of global commodity chains.
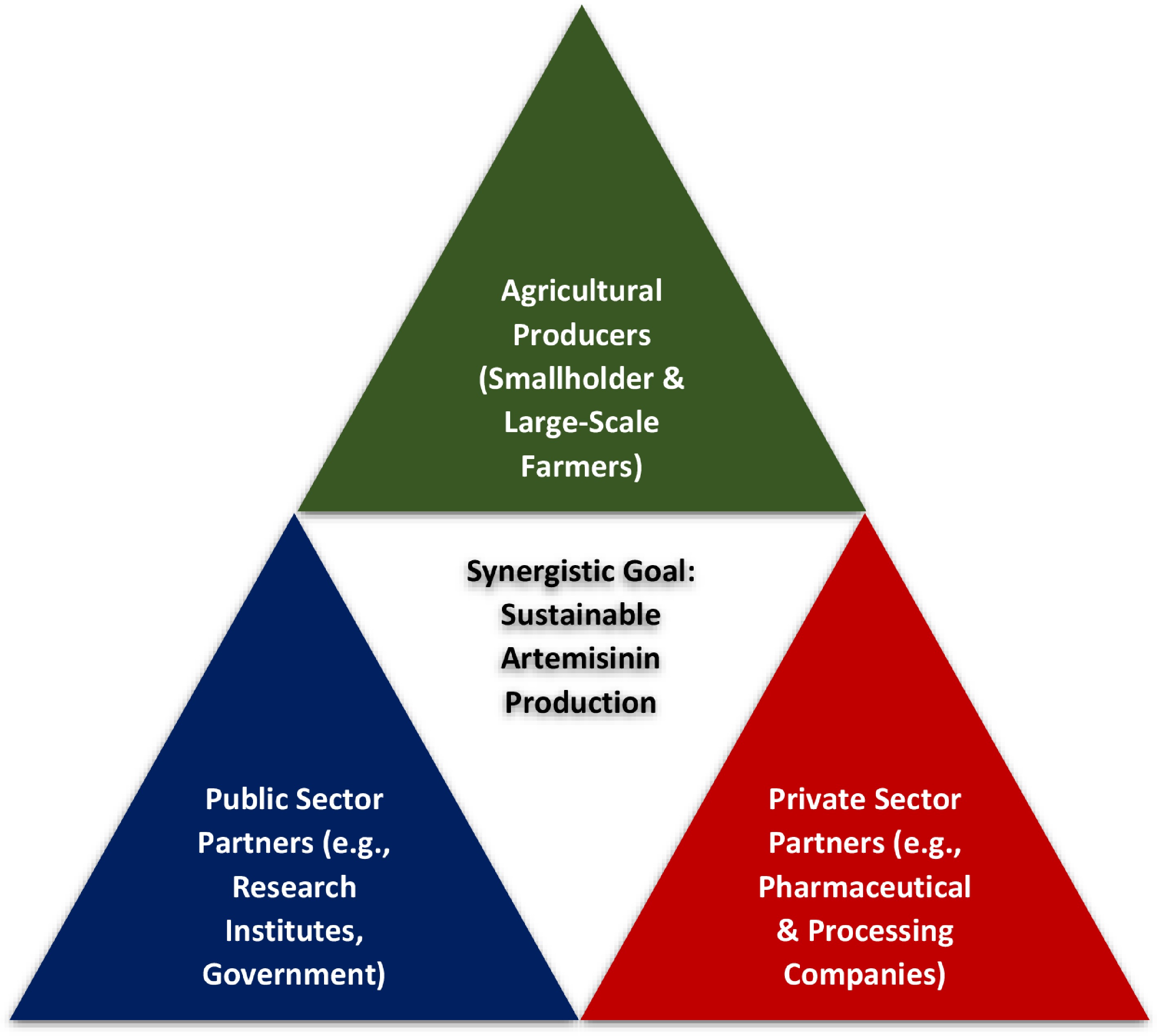
Figure 3.
Proposed Public-Private-Farmer Partnership (PPFP) model for a sustainable artemisinin value chain.
To move beyond a purely technical framework, the expansion of A. annua cultivation must explicitly address structural asymmetries in value distribution, where primary producers typically capture only a small fraction of final pharmaceutical value[129]. In global value chains, economic rents are often concentrated in high–value-added stages such as extraction, formulation, and retailing, leaving smallholders exposed to price volatility and extractive forms of 'data colonialism'[105,130]. Within PPFPs, this framework therefore integrates three interlinked pillars to promote social equity and fair value distribution: price transparency, contractual safeguards, and inclusive collective governance, with gender-transformative design as a cross-cutting principle.
First, price transparency enabled by digital traceability is essential for correcting information asymmetries that allow intermediaries to suppress farm-gate prices[131]. Mobile-ledger systems and blockchain-enabled traceability can create conditions of 'hyper-transparency,' providing farmers with real-time access to market prices, transaction records, and quality verification[105]. Such systems reconfigure power relations by shifting farmers from passive 'ICT tenants,' who merely supply data, to active 'data governors,' who understand their data rights and leverage information to strengthen their bargaining position[130].
Second, contractual safeguards based on value chain profit sharing are required to buffer producers against artemisinin price crashes. Rather than relying on rigid fixed-price contracts, the framework recommends a 'Value Chain Profit Sharing' model that distributes payments in multiple instalments[132]. An initial, conservative payment at harvest ensures immediate liquidity and covers production costs[133], followed by a secondary payment once downstream sales are realized, reflecting actual pharmaceutical profits. This structure mitigates the 'double marginalization' effect, allowing farmers to participate in upside market gains while being protected from downside volatility[132,134].
Third, collective governance and upward vertical integration strengthen farmers' structural position within the value chain. Individual smallholders often lack the leverage to negotiate with dominant buyers or processors[135]. Farmer-led cooperatives enable collective action and partial downstream ownership—such as primary drying, grading, and leaf–stem separation—thereby allowing producers to retain a greater share of value added[133,136]. Beyond economic gains, collective governance builds social capital, reinforcing community resilience, self-determination, and long-term adaptive capacity[129,135].
Across all these mechanisms, gender inclusion and targeted capacity building are indispensable. Women constitute approximately 43% to over 50% of the agricultural workforce in low-income countries and play a central role in labor-intensive nursery management and post-harvest handling of A. annua[129,130]. Yet a persistent 'gender data gap' often renders their labor invisible and unpaid. Equitable PPFP design, therefore, requires gender-transformative co-design processes that ensure women's representation in cooperative leadership and provide targeted training in quality-enhancing and income-generating practices[130,133].
Table 8 below compares complementary organizational mechanisms designed to address structural inequities in the A. annua value chain. Rather than acting in isolation, these models are most effective when embedded within Public–Private–Farmer Partnerships (PPFPs), where digital transparency, contractual innovation, and collective governance jointly counteract price volatility, information asymmetry, and unequal value capture. Together, they enable a transition from extractive commodity relationships toward inclusive, agroecologically grounded value chains that align farmer livelihoods with pharmaceutical supply security. Together, these organizational and contractual interventions—summarized in Table 6—demonstrate how PPFPs can be operationalized to redistribute value, enhance transparency, and embed social equity within the A. annua value chain.
Table 8. Comparison of organizational models for smallholder equity in the A. annua value chain.
Organizational mechanism Core institutional innovation How value redistribution is achieved Equity outcomes for smallholders Key limitations and governance risks Evidence base Blockchain-enabled digital ledgers Decentralized, tamper-resistant transaction and traceability systems Creates real-time visibility of prices, volumes, quality grades, and payments across the value chain; reduces information asymmetry between farmers and buyers Improves farm-gate price transparency; lowers search and transaction costs; strengthens farmers' bargaining power and data sovereignty Requires digital literacy and connectivity; risk of elite capture if data governance rules are unclear; potential exclusion of farmers without access to digital tools [105,
130,131]Value chain profit-sharing contracts Multi-installment payment structures linked to downstream sales rather than fixed farm-gate prices Distributes risk and reward across the chain by aligning farmer payments with realized pharmaceutical profits Stabilizes income; buffers farmers against artemisinin price crashes; allows participation in upside market peaks while limiting downside exposure Requires high trust and transparent accounting; dependent on enforceable contracts and buyer compliance [132−134] Farmer-led vertical integration Cooperative ownership of early processing stages (drying, grading, leaf–stem separation) Retains value addition at the producer level by capturing margins typically absorbed downstream Increases share of final value retained locally; reduces dependency on intermediaries; enhances income resilience Capital and managerial requirements; risk of cooperative mismanagement without strong governance structures [133,136] Collective governance (cooperatives and producer organizations) Democratic decision-making structures and pooled negotiation power Aggregates production volume and voice to improve negotiating leverage with buyers and service providers Enhances bargaining power; builds social capital; supports collective risk management and long-term resilience Effectiveness contingent on trust, inclusivity, and internal accountability; risk of elite domination [129,135] Gender-transformative inclusion models Explicit integration of women into leadership, data systems, and training pathways Makes women's labor visible and remunerated; ensures equitable access to skills, information, and decision-making Improves household income equity; strengthens quality control in labor-intensive nursery and post-harvest stages Cultural norms may limit participation; requires sustained institutional commitment and monitoring [129,
130,133]These socio-economic enablers also provide the necessary foundation for market-based incentives. Certification schemes for 'Agroecologically-Grown' or 'Biodiversity-Friendly' artemisinin can only function credibly if transparent digital ledgers and traceability systems are already in place. Such certification would allow pharmaceutical companies, donors, and global health programs to preferentially source raw materials that meet high environmental and social standards, thereby translating ecological stewardship and fair governance into tangible price premiums for farmers.
Complemented by supportive government policies—such as transition subsidies, agroforestry establishment grants, and crop insurance—and by strengthened extension systems focused on soil health, integrated pest management, and biodiversity monitoring, these measures align social equity with ecological sustainability. In doing so, they reinforce the central premise of this framework: securing a stable global supply of artemisinin for malaria control must proceed in parallel with protecting ecosystem integrity and ensuring dignified, resilient livelihoods for the farmers at the foundation of the value chain.
This literature-based review proposes an integrated framework that moves beyond a narrow focus on yield maximization toward a holistic strategy grounded in the principles of agrobiodiversity. By synergizing advanced agronomic practices, climate-adaptive breeding, intelligent farming technologies, and robust ecological governance, A. annua cultivation can be expanded in a manner that is both productive and environmentally sustainable. These coordinated elements align with the key milestones required to establish a resilient artemisinin supply chain, as summarized in Fig. 4.
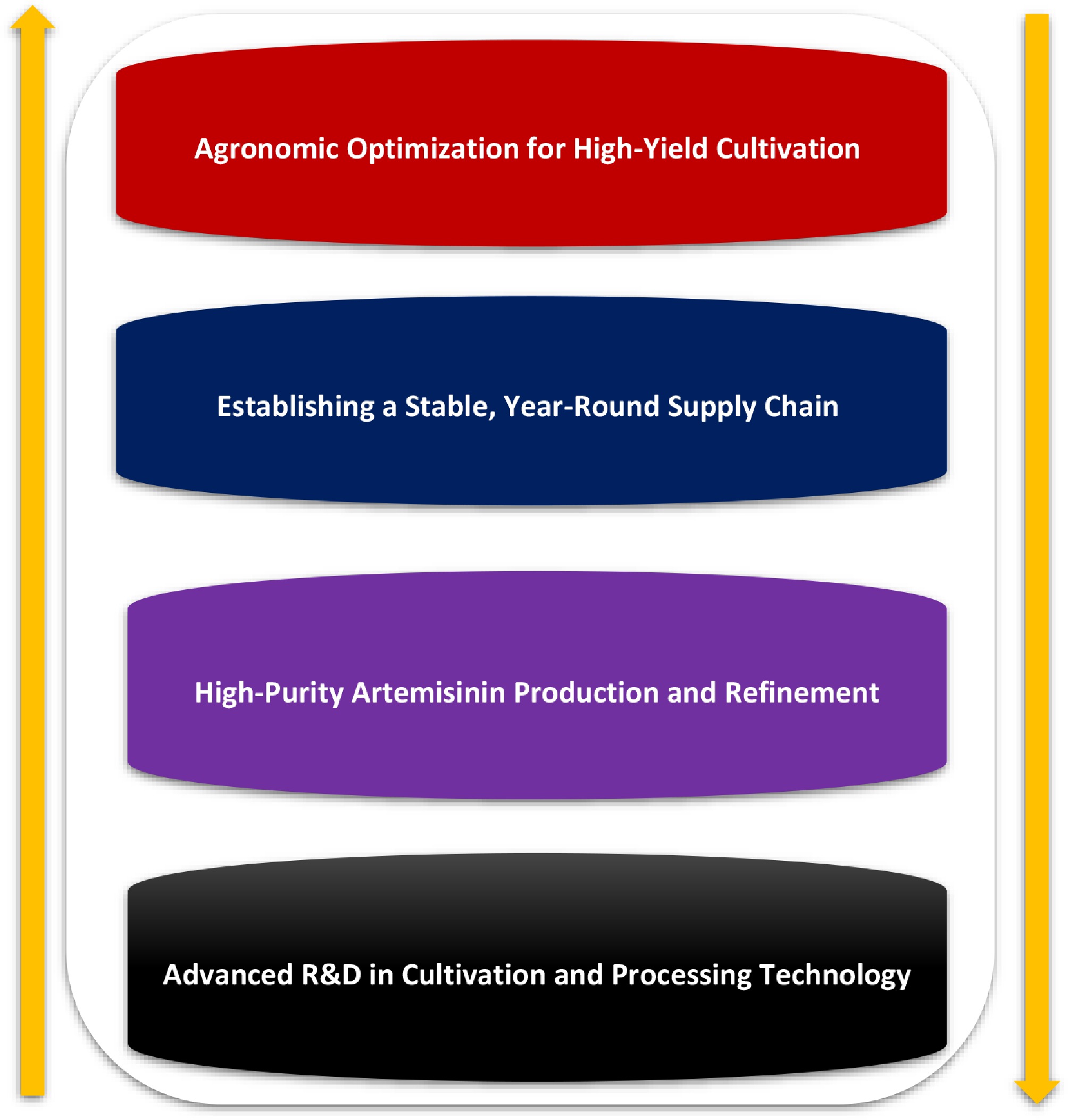
Figure 4.
Strategic milestones for optimizing the A. annua to artemisinin production pipeline.
Figure 4 provides a schematic overview of the critical objectives linking agricultural production to pharmaceutical supply. The framework begins with (1) agronomic optimization to ensure high germination rates and maximize field-level yields. This agricultural foundation enables (2) a stable and year-round supply of biomass, thereby reducing vulnerability to seasonal fluctuations. Downstream processing is subsequently refined to (3) generate high-purity artemisinin suitable for pharmaceutical applications. The entire pipeline is continuously strengthened through (4) ongoing research and development, in which advances in tissue culture, process optimization, and continuous extraction technologies drive further gains in yield, purity, and system-level sustainability.
Box 2: Proposed roadmap for empirical validation of the integrated framework
To transition the proposed framework from a conceptual synthesis to an empirically validated model, a phased, multi-year experimental program is recommended. This roadmap is designed to generate robust agronomic, ecological, and socio-economic evidence under real-world smallholder conditions, consistent with the standards of large-scale sustainability research.
Experimental design overview (three-year, multi-location trial)
A coordinated field experiment should be implemented across three distinct agro-ecological zones representing contrasting climatic and socio-economic contexts (e.g., subtropical humid, tropical sub-humid, and semi-arid systems). Within each zone, trials would be conducted using a randomized complete block design with farmer-managed replication.
Treatment structure
Three production systems are proposed for side-by-side comparison:
1. Standard monoculture (control): Conventional A. annua monoculture using locally prevalent cultivars and baseline agronomic practices, representing current production norms.
2. Biodiversity-integrated agronomic system: A. annua cultivated in diversified systems (e.g., intercropping with nitrogen-fixing legumes or rotational integration with staple crops), combined with organic soil amendments and reduced synthetic inputs.
3. Fully integrated framework: Biodiversity-integrated agronomy combined with low-cost intelligent farming tools (e.g., shared IoT soil-moisture sensors, SMS-based advisory systems, or cooperative drone services) and embedded within a Public–Private–Farmer Partnership (PPFP) governance model.
Core metrics for evaluation
To enable cross-scale comparison, standardized indicators should be measured annually across all treatments:
1. Productivity metrics: Artemisinin yield per hectare (kg·ha−1), total leaf biomass, and harvest stability across seasons.
2. Ecological metrics: Soil microbial diversity indices (e.g., Shannon or Simpson indices derived from 16S/ITS profiling), soil organic carbon, and incidence of pest and disease outbreaks.
3. Resource-use efficiency: Water-use efficiency and nutrient-use efficiency, particularly in intelligent-farming-enabled plots.
4. Socio-economic outcomes: Net farmer income (USD ha−1), income variability over time, and equity metrics capturing benefit distribution within PPFP arrangements.
5. Governance and adoption indicators: Farmer participation rates, technology uptake, and gender-disaggregated labor and income outcomes.
Validation pathway and scaling logic
Data generated during the first two years would support iterative optimization of agronomic and technological components, while the third year would focus on system stabilization and comparative performance analysis. Cross-site synthesis would enable identification of context-specific versus generalizable effects, providing the empirical basis for scaling recommendations and policy engagement.
By explicitly linking field-level agronomic performance, ecosystem responses, and socio-economic outcomes, this roadmap establishes a clear validation pathway through which the conceptual framework can be tested, refined, and translated into evidence-based deployment strategies.
-
This review demonstrates that productivity, sustainability, and social equity in A. annua cultivation are not mutually exclusive objectives. On the contrary, practices that enhance agrobiodiversity—such as intercropping, diversified rotations, integrated soil management, and the conservation of genetic resources—also strengthen ecological resilience, reduce agronomic risk, and contribute to more stable artemisinin yields over time. When embedded within inclusive governance structures, these practices further support improved and more equitable livelihood outcomes for smallholder farmers.
The framework presented here is intentionally conceptual and literature-based. Its practical effectiveness will depend on systematic empirical validation through coordinated, multi-location field trials and socio-economic case studies, as outlined in Box 2. Such validation is essential for distinguishing context-specific effects from generalizable principles and for translating the framework into evidence-based policy and investment strategies.
Several limitations and challenges must be acknowledged. Implementation requires sustained political commitment, enabling policy environments, and long-term investment from both public and private actors. Market-based incentives, including certification and price premiums, remain vulnerable to shifts in pharmaceutical procurement policies and donor priorities. Moreover, while diversification can buffer economic shocks, artemisinin price volatility remains an external risk that cannot be fully eliminated at the farm level.
Future progress, therefore, hinges on collaborative action across the artemisinin value chain. Public–Private–Farmer Partnerships (PPFPs), supported by inclusive intelligent farming systems and biodiversity-based agronomic design, offer a viable pathway for aligning pharmaceutical supply security with ecological stewardship and social justice. By harnessing digital technologies not only for efficiency but also for monitoring and enhancing agroecosystem complexity, A. annua cultivation can evolve from a volatile commodity system into a model for sustainable, resilient medicinal plant production. Such a transition is critical for safeguarding both a life-saving medicine and the ecological foundations upon which its supply ultimately depends.
-
The author confirms sole responsibility for the following: study conception and design, data collection, analysis and interpretation of results, manuscript preparation, and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The corresponding author thanks Chief MOSOH Paul Tandong and Chieftess Ateyim Espe MOSOH Ostensia Nkeng of PINYIN (Santa, North-West Region, Cameroon) for their relentless support. The USDA National Institute of Food and Agriculture has supported this study, specifically under the Hatch project 7001563. The author extends sincere gratitude to Professor Wagner A. Vendrame, Horticultural Sciences Department at the University of Florida, for his invaluable support and insightful feedback during the review of the final draft of this manuscript. His expertise and guidance have greatly enhanced the quality of this work. Additionally, the author expresses his sincere appreciation to the reviewer(s) for their diligent review of this work and their insightful comments and suggestions, which have significantly enriched the quality of this publication.
-
The author declares having no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Yunnan Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Mosoh DA. 2026. An integrated agrobiodiversity framework for sustainable Artemisia annua L. cultivation. Agrobiodiversity 3(2): 55−70 doi: 10.48130/abd-0026-0005 

An integrated agrobiodiversity framework for sustainable Artemisia annua L. cultivation
- Received: 05 August 2025
- Revised: 16 February 2026
- Accepted: 15 April 2026
- Published online: 28 May 2026
Abstract: Medicinal plants are foundational to global health systems, yet their increasing integration into modern pharmaceutical supply chains has intensified sustainability challenges associated with climate change, biodiversity loss, and market volatility. Artemisia annua L., the sole commercial source of artemisinin—the cornerstone of artemisinin-based combination therapies for malaria—exemplifies these tensions. Despite decades of agronomic and biotechnological research, global artemisinin supply remains unstable, largely due to yield variability, monoculture-driven ecological risks, and socio-economic vulnerabilities faced by smallholder producers. This literature-based review synthesizes advances across agronomy, climate-adaptive breeding, artificial intelligence–enabled precision agriculture, and socio-economic governance to propose an integrated framework for the sustainable expansion of A. annua cultivation. Central to the framework is the repositioning of agrobiodiversity—from a perceived constraint on productivity to a primary mechanism for enhancing ecological resilience, yield stability, and farmer livelihoods. The framework emphasizes biodiversity-centric agronomic practices, multi-latitude phenological adaptation, inclusive intelligent farming systems, and Public–Private–Farmer Partnerships (PPFPs) designed to promote equitable value distribution. While the framework is conceptual in nature, it is grounded in empirical evidence from diverse disciplinary domains. To facilitate translation into practice, a structured roadmap for empirical validation is proposed, outlining multi-location, multi-year field trials integrating agronomic, ecological, and socio-economic metrics. Collectively, this work provides a systems-level blueprint for stabilizing the global artemisinin supply while aligning medicinal plant production with biodiversity conservation, climate resilience, and social equity objectives.