









-
Perennial ryegrass (Lolium perenne L.) is a high-value cool-season species widely utilized as premium turfgrass and nutritious forage, prized for its high protein content, extended growing season, strong grazing tolerance, and excellent palatability. As the most extensively cultivated perennial forage grass in temperate regions, it plays a critical role in landscaping and ecological management[1,2]. However, climate change has led to increasingly frequent and severe abiotic stresses—including heat waves, drought, soil salinization, and heavy metal contamination. These stresses can significantly disrupt plant hormone homeostasis, while phytohormones like abscisic acid (ABA), auxin, jasmonic acid, etc, in turn, play critical regulatory roles in mediating plant responses and adaptation to such environmental challenges. The adverse effects markedly impair its growth, yield, and persistence, thereby threatening agricultural sustainability and food security[3,4]. In this context, uncovering the genetic basis of stress adaptation is essential for breeding resilient cultivars through molecular approaches[5,6].
Among key regulators of plant stress responses, the Heat Shock Factor (HSF) gene family stands out as a master transcription factor family involved in diverse abiotic and biotic stresses[7,8]. HSFs are characterized by conserved domains such as the N-terminal DNA-binding domain (DBD) and the oligomerization domain (HR-A/B), and are classified into subfamilies A, B, and C based on structural features and functional motifs—including activation domains (AHA) in class A and repressor domains in class B[9]. Functional studies across species have revealed their critical roles: in Arabidopsis, AtHSFA1 acts as a master regulator of heat stress response (HSR)[10], while AtHSFA2 overexpression improves tolerance to heat, salt, and osmotic stress[11]. In maize, ZmHSF20 negatively regulates thermotolerance by modulating cell wall-related genes[12], whereas in lily, LlHSFC2 forms heteromeric complexes with HSFAs to enhance proteostasis under heat stress[13].
Although HSFs have been identified in several forage grasses—including 16 members in Italian ryegrass[14] and 25 in an earlier perennial ryegrass genome[15]—their functional characterization remains fragmentary, largely limited to heat stress. A comprehensive expression atlas capturing their roles across multiple stresses and developmental contexts is still lacking. This study systematically identified 26 LpHSFs from the updated perennial ryegrass genome, classifying them into 16 class A, six class B, and four class C members. A spatio-temporal expression atlas of LpHSFs was further established under diverse abiotic stresses (heat, salt, alkali, heavy metals) and hormone treatments, as well as across tissue types and senescence stages. Beyond confirming their canonical heat-responsive roles, broad-spectrum stress responsiveness and tissue-specific regulation were uncovered, highlighting the functional diversification of LpHSFs within the plant stress adaptation network. This work provides a foundational resource for elucidating HSF-mediated regulatory mechanisms and supports molecular breeding aimed at enhancing multi-stress resilience in perennial ryegrass and related species.
-
The protein sequences of HSF family members from Arabidopsis thaliana, Zea mays, and Oryza sativa were retrieved from the Plant Transcription Factor Database (PlantTFDB;
http://planttfdb.cbi.pku.edu.cn )[16]. HSF protein sequences from perennial ryegrass were acquired from its published genome[17] and used to construct a local protein database. Using BLASTP, proteins containing the HTH (helix-turn-helix) domain were identified by querying with the HSF sequences from A. thaliana, Z. mays, and O. sativa. Candidate genes were further screened using the hidden Markov model (HMM) profile of the HTH domain (PF00447) obtained from the PFAM database (http://pfam.xfam.org ). Redundant sequences were filtered using the online tools HMMER (www.hmmer.org ), and SMART (http://smart.embl-heidelberg.de ). Non-redundant sequences and those predicted with high confidence were selected and designated as LpHSF genes. The coding sequences (CDS) of the LpHSF genes were extracted from the perennial ryegrass genome annotation files. Finally, the physicochemical properties of the LpHSF proteins, including molecular weight (MW) and theoretical isoelectric point (pI), were predicted using ProtParam (https://web.expasy.org/protparam ).Phylogenetic analysis and classification of the LpHSF gene family
-
The HSF proteins of Lolium perenne, Oryza sativa, Triticum aestivum, Zea mays, and Arabidopsis thaliana were downloaded from the perennial ryegrass[17], rice[18], wheat[19], corn[20] genome annotation project, and TAIR11[21], respectively. The HSF protein sequences of Lolium perenne, Oryza sativa, Triticum aestivum, Zea mays, and Arabidopsis thaliana were used for multiple sequence alignment with Muscle[22]. The maximum-likelihood phylogenetic tree was constructed using IQ-tree[23] with the VT+F+R10 model identified as the best substitution model using ModelFinder[24]. Ultrafast bootstrap[25] with 1,000 replications was used to test the branch supports. The LpHSF genes were divided into different subgroups on the basis of the classification of perennial ryegrass, rice, wheat, corn, and Arabidopsis HSF.
Gene structure, cis-acting analysis, and chromosomal locations
-
To visualize the exon-intron structure of LpHSF genes, the gene structure was displayed using TBtools[26], according to the perennial ryegrass genome annotation file[17]. The analysis of cis-acting elements of 2,000 bp upstream of the 26 LpHSF genes was performed by using the PlantCARE software (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) and PlantPAN 4.0 (http://plantpan.itps.ncku.edu.tw/plantpan4/index.html ), and then the illustrations were drawn using the TBtools[26].The chromosomal locations of the LpHSF genes were obtained from the perennial ryegrass genome annotation files and mapped to the corresponding chromosomes using TBtools[26]. In order to beautify the chromosome distribution of LpHSF genes, the online tool MapGene2Chrome (MG2C,
http://mg2c.iask.in/mg2c_v2.1 )[27] was used to make it.Plant materials and growth conditions
-
The perennial ryegrass seeds (cv. 'Buena Vista') were germinated at Sichuan Agricultural University in a growth chamber for 10 d. Germination was carried out in petri dishes lined with filter paper and moistened with sterile water. The seedlings were then transplanted into a hydroponic box containing 1/2 Hoagland (pH = 5.8) for 4 weeks. The Hoagland's solution was changed every 3 d. The growth temperature was 25/22 °C (day/night) with and a 16/8 h light/dark photoperiod.
Stress treatments and several hormone treatment
-
To assess the expression patterns in response to various treatments, one-month-old perennial ryegrass seedlings were subjected to drought stress (20% PEG6000), salt stress (250 mM NaCl), heat stress (38/33 °C, day/night), alkaline stress (200 mM, pH = 9.4), heavy metal stress (1.33 mM CdCl2)[28,29], and phytohormones treatment[30] such as gibberellins (GA, 100 μM), auxin (IAA, 20 μM), abscisic acid (ABA, 50 μM), salicylic acid (SA, 100 μM), methyl jasmonate (MeJA, 20 μM), ethephon (ETH, 200 μM), N-(Phenylmethyl)-9H-purin-6-amine (6-BA, 25μM). After 0, 1, 4, 12, and 72 h of different treatments, samples were collected, rapidly frozen in liquid nitrogen, and stored at −80 °C before RNA extraction.
RNA extraction, cDNA synthesis, and RT-qPCR analysis
-
Total RNA was isolated using the HiPure Plant RNA Mini Kit (Magen Biotech Co., Ltd., China), according to the manufacturer's protocol. MonScript™ RTIII All-in-One Mix with dsDNase (Monad Biotech Co., Ltd., China) was used for the synthesis of cDNA. RT-qPCR analyses were performed using the Tag SYBR® Green qPCR Premix x (BestEnzymes Biotech Co., Ltd., Canada), in accordance with the manufacturer's protocol; the reactions were run using the CXF Connect™ Real-Time System (Bio-Rad). The EUKARYOTIC INITIATION FACTOR 4 ALPHA (EIF4A) was selected as the internal reference gene. Relative expression was calculated using the 2−ΔΔCᴛ method[31]. The primers used in this study are listed in Supplementary Table S1.
Statistical analysis
-
All statistical analyses were conducted in JMP software (Version 10, SAS Institute Inc., USA). Results are reported as mean values ± SEM derived from three independent biological replicates. To determine the statistical significance of differences in LpHSFs transcript abundance among different organs and in response to various abiotic stresses and phytohormone treatments, Fisher's protected least significant difference (LSD) test was applied, with a probability level of 0.05 considered statistically significant.
-
Based on the newly released genome version of perennial ryegrass[17], a total of 26 HSF genes (LpHSFs) containing the characteristic HTH domain were identified and systematically classified into three classes (A, B, and C) with 16, six, and four members, respectively, and named accordingly from LpHSFA1B to LpHSFC1.4 (Fig. 1a). The coding sequence lengths of these LpHSFs ranged from 369 bp (A7A) to 2,067 bp (A1B), while the molecular weights of the encoded proteins varied from 1,2581.2 Da (A7A) to 74,431.97 Da (A1B), with isoelectric points spanning 4.74 (A2.5) to 10.69 (C1.1).
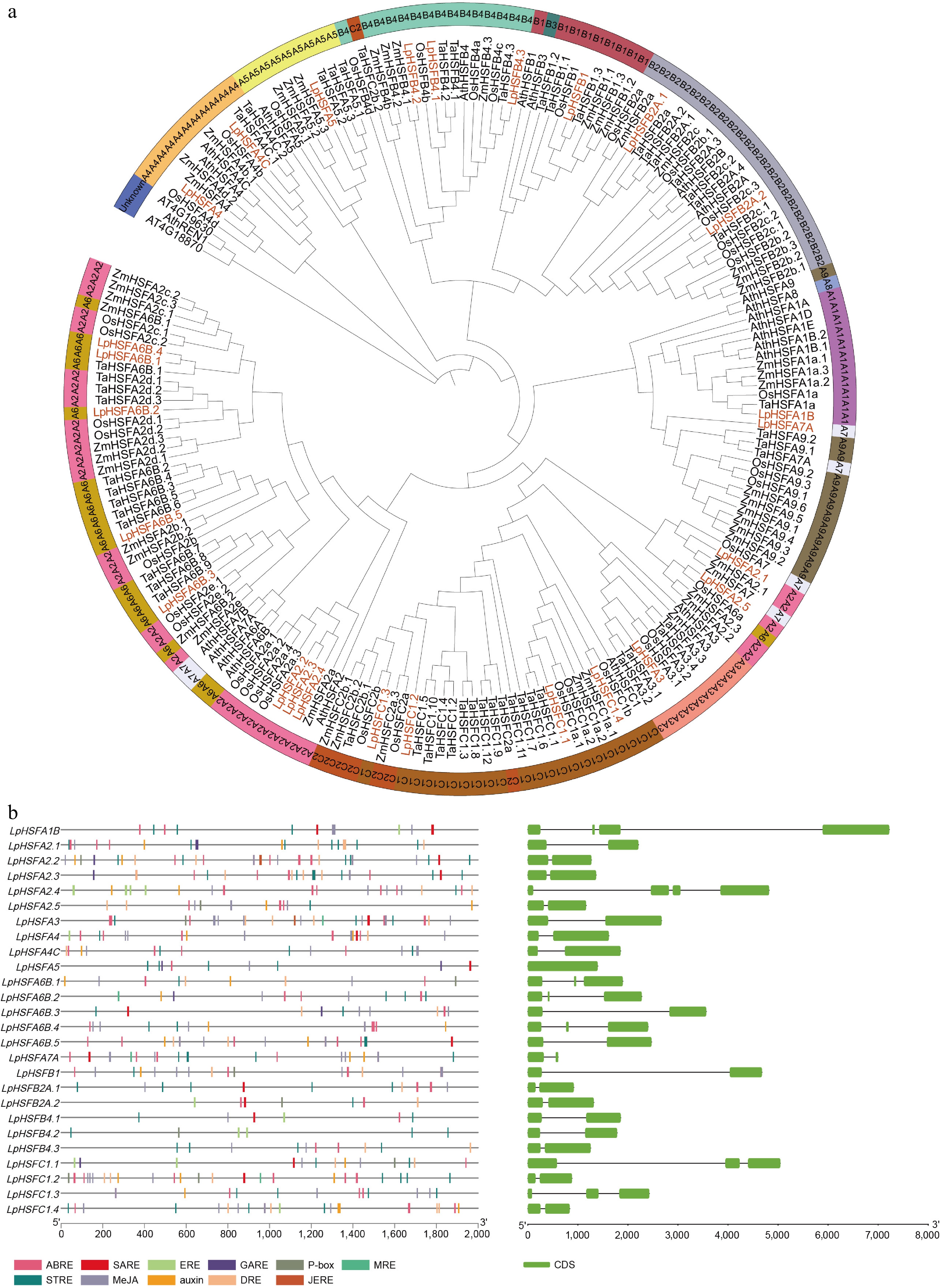
Figure 1.
(a) Unrooted phylogenetic tree representing relationships among the HSF proteins of Lolium perenne, Oryza sativa, Triticum aestivum, Zea mays, and Arabidopsis thaliana. The genes in Lolium perenne are marked in orange, while those in Oryza sativa, Triticum aestivum, Zea mays, and Arabidopsis thaliana are marked in black. For the identification of subgroup A, the following color scheme was applied: A1 (mediumorchid), A2 (hotpink), A3 (lightsalmon), A4 (lightorange), A5 (khaki), A6 (goldenrod), A7 (aliceblue), A8 (skyblue), and A9 (olivedrab). In subgroup B, B1, B2, B3, and B4 were assigned the colors indianred, bluegray, slategray, and aquamarine, respectively. Within subgroup C, C1 was colored dark_goldenrod, and C2 was designated chocolate. Non-grouped members were uniformly colored slate_blue. (b) The cis-acting elements of the LpHSFs promoter region from perennial ryegrass, and gene structures of LpHSFs from perennial ryegrass in which the exons region is indicated by green boxes.
To elucidate their evolutionary relationships, a phylogenetic tree was constructed using HSF protein sequences from Lolium perenne, Oryza sativa, Triticum aestivum, Zea mays, and Arabidopsis thaliana. The classification was further supported by distinct functional domains: Class A members possess an activation domain (AHA), Class B contains a repressor domain (RD), and Class C comprises the remaining members. Phylogenetic analysis revealed that LpHSFs cluster closely with other Gramineae homologs, indicating a common evolutionary origin. Notably, this study identifies 26 LpHSF members, differing from the previous report[15] by the inclusion of additional members such as A6B.4, A7A, and A2.4, a discrepancy likely attributable to the improved genome assembly used here. Detailed information on all identified LpHSF genes is provided in Supplementary Table S2.
Genomic distribution and putative functional analysis of LpHSF genes
-
Chromosomal localization analysis revealed that the 26 identified LpHSF genes are distributed across all seven chromosomes. As illustrated in Supplementary Fig. S1, chromosome 4 harbors the highest number, with eight genes (A1B, A6B.3, A2.2, A2.3, A2.4, B4.2, A7A, and A6B.2). Five genes (A6B.4, A5, B4.3, B1, and B2A.2) are located on chromosome 5, while four ones (A2.1, C1.1, A4C, and C1.4) reside on chromosome three. Chromosome 2 contains three HSFs (B4.1, A6B.5, and B2A.1), and chromosomes 1, 6, and 7 each contain only two genes.
To gain functional insights, the cis-elements in the promoters of these genes (pLpHSFs) were analyzed using PlantCARE. The results indicate that most pLpHSFs contain a variety of hormone-responsive and mRNA recognition elements (Fig. 1b; Supplementary Table S3), suggesting potential roles in abiotic stress responses and hormone signaling pathways. Furthermore, gene structure analysis showed diversity in intron-exon organization (Fig. 1b). Specifically, A5 is intronless, A1B and A2.4 contain three introns, five genes (A6B.1, A6B.2, A6B.4, C1.1, and C1.3) have two introns, and the remaining LpHSF genes possess a single intron.
Expression profiling of LpHSF genes across tissues
-
The tissue-specific expression profiles of LpHSF genes were investigated to elucidate their potential roles in development and leaf senescence (Fig. 2). The analysis revealed that 23 out of 26 LpHSFs were highly expressed in leaves at different developmental stages. Strikingly, 15 A-type, three B-type, and two C-type LpHSFs displayed peak expression in the older, lower-positioned leaves (4th or 5th). Notably, six genes (A2.4, A4, A4C, A6B.2, A6B.3, B2A.1) exhibited leaf-specific expression.
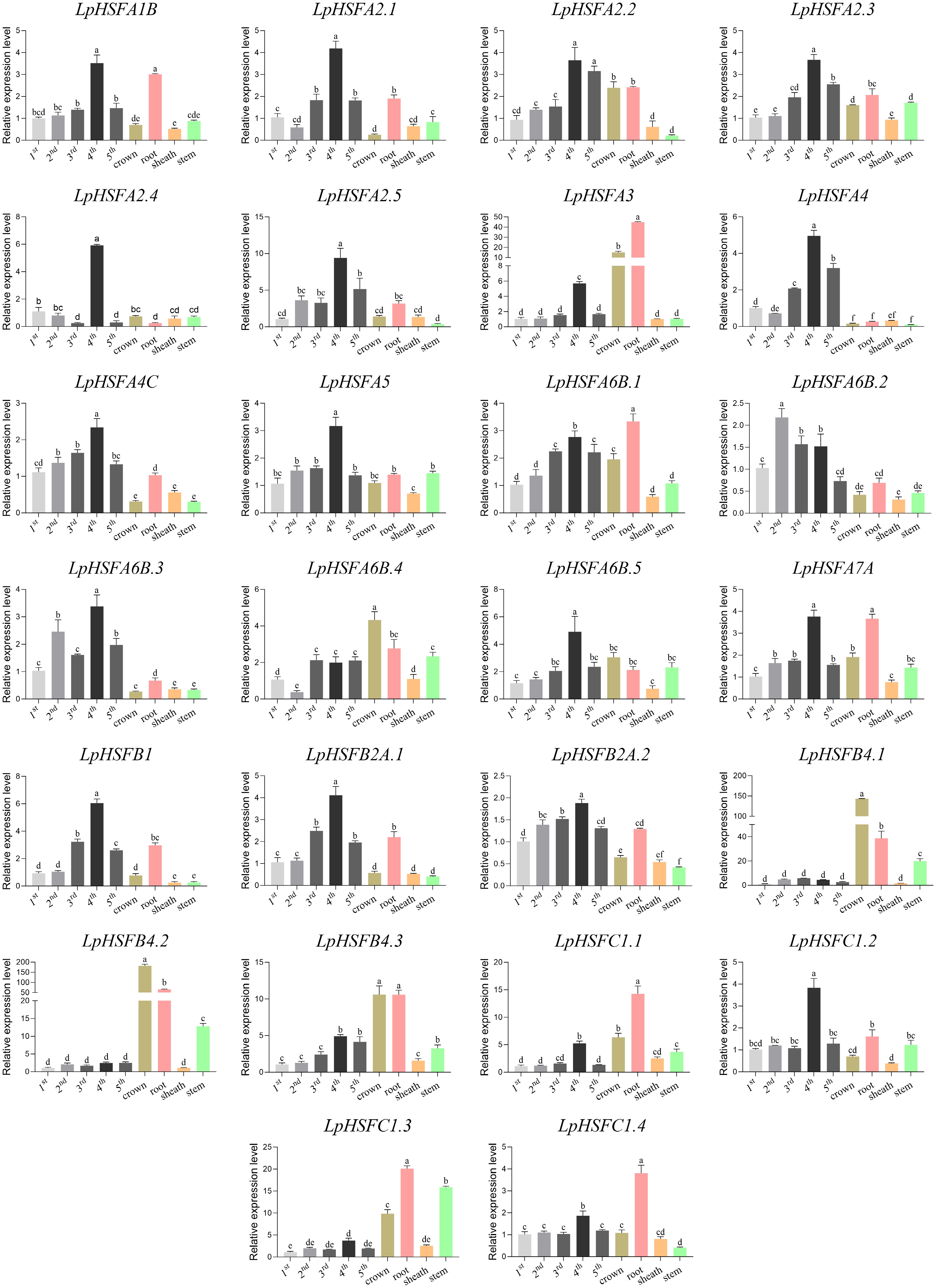
Figure 2.
Expression patterns of LpHSF in different tissues. The statistical significance was determined using Fisher's protected least significant difference (LSD) test (p < 0.05).
In roots, 21 LpHSFs showed relatively high expression, with all B- and C-type members included. The C-type genes C1.1, C1.3, and C1.4 were particularly prominent, showing their highest transcript abundance in root tissue.
Within the crown tissue, 13 LpHSFs were highly expressed. Among these, A6B.4, B4.1, B4.2, and B4.3 achieved their maximum expression levels in the crown, with B4.1 and B4.2 exhibiting expression approximately 3.5-fold and 3-fold higher, respectively, than in their next highest-expressing tissue (root).
Only ten LpHSFs were highly expressed in the leaf sheath, and for all of them, the sheath was not their primary site of expression.
Expression profiles of LpHSF response to phytohormones
-
Although the presence of abundant hormone-responsive cis-regulatory elements in pLpHSFs suggested their potential regulatory roles, expression profiling of all 26 LpHSFs was conducted under phytohormone treatments using RT-qPCR. As shown in Fig. 3, the expression of most LpHSFs was unstable under control conditions.
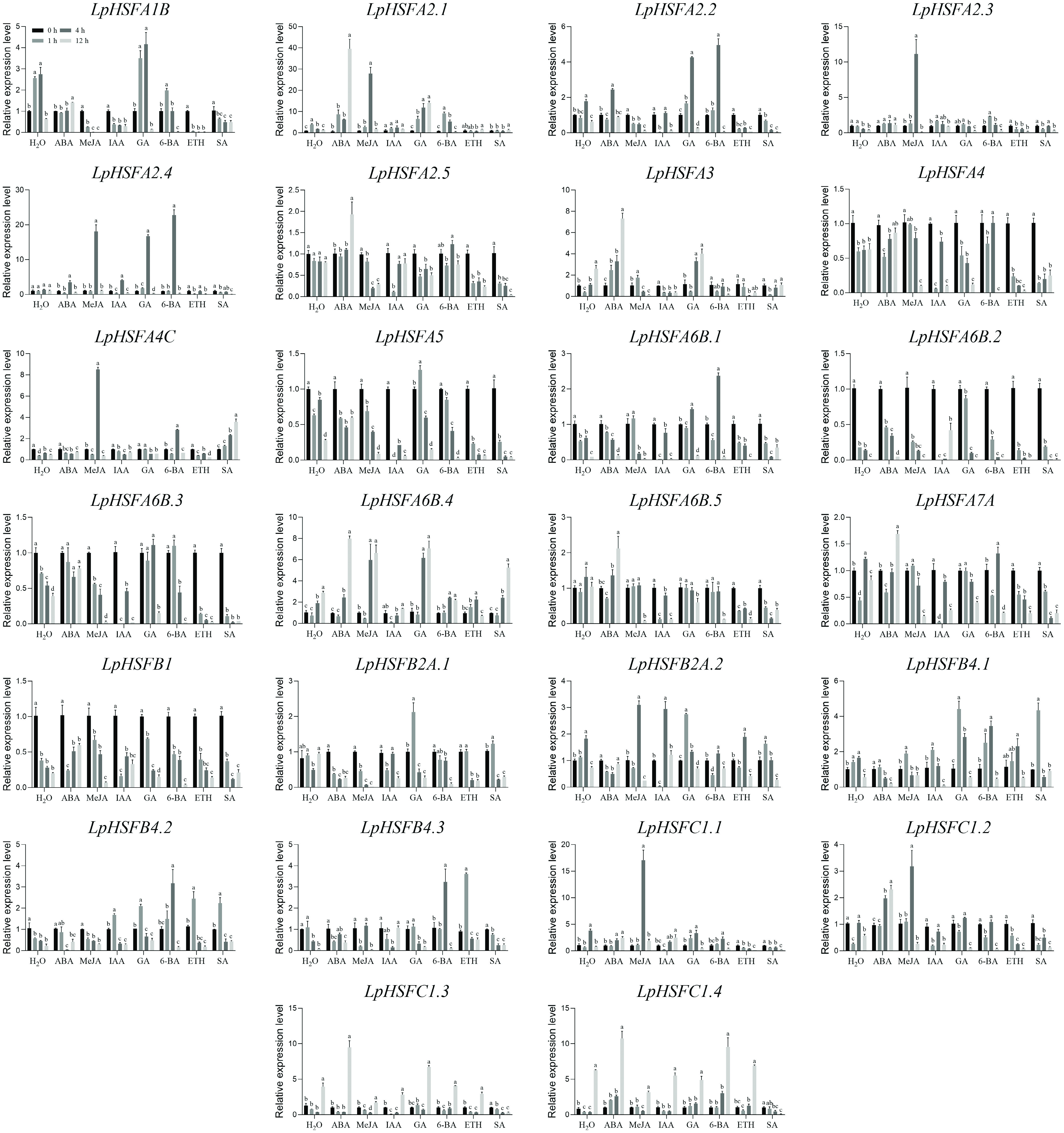
Figure 3.
Expression profiles of 26 LpHSF genes in response to ABA, MeJA, IAA, GA, 6-BA, ETH, and SA treatments. The vertical bars indicate standard error. The statistical significance was determined using Fisher's protected least significant difference (LSD) test (p < 0.05).
In response to ABA, half of the subfamily A members exhibited significant expression changes, including seven upregulated genes (A2.1, A2.4, A2.5, A3, A6B.4, A6B.5, A7A) and one downregulated gene (A1B). In contrast, only three subfamily B members (B4.1, B2A.1, B2A.2) were repressed, and three subfamily C members (C1.2, C1.3, C1.4) were upregulated.
Under MeJA treatment, 11 out of 16 subfamily A members responded, with six being upregulated (A2.1, A2.3, A2.4, A4C, A6B.1, A6B.4) and five downregulated (A1B, A2.2, A2.5, A6B.5, A7A). In subfamily B, B2A.2 was induced, and B2A.1 was suppressed. Among subfamily C members, C1.1 and C1.2 were upregulated, while C1.3 was downregulated.
Following IAA treatment, A2.3, A2.4, and A6B.2 were upregulated in subfamily A, whereas A1B, A3, A5, and A6B.4 were downregulated. In subfamilies B and C, only B2A.2 and B4.2 were induced.
Under GA treatment, ten subfamily A members (A1B, A2.1, A2.2, A2.4, A3, A5, A6B.1, A6B.2, A6B.3, A6B.4) were upregulated, while A4, B2A.1, B2A.2, B4.1, B4.2, C1.3, and C1.4 were downregulated.
In response to 6-BA, nine subfamily A members (A2.1, A2.2, A2.4, A2.5, A4, A4C, A6B.1, A6B.3), four subfamily B members (B4.1, B4.2, B4.3), and one subfamily C member (C1.4) were upregulated, whereas A1B, A3, and B2A.2 were downregulated.
Under ETH treatment, only B4.2 and B4.3 were upregulated, while 12 subfamily A members (A1B, A2.1, A2.2, A2.4, A2.5, A4, A5, A6B.2, A6B.3, A6B.4, A6B.5, A7A) and one subfamily C member (C1.1) were downregulated.
After SA treatment, two subfamily A members (A4C, A6B.4) and two subfamily B members (B4.1, B4.2) were upregulated, while nine subfamily A members (A1B, A2.2, A2.5, A3, A4, A5, A6B.3, A6B.5, A7A) and three subfamily C members (C1.1, C1.2, C1.3) were downregulated.
Notably, A6B.2 showed marked downregulation across multiple treatments. Within class B, all genes except B1 exhibited no hormone responsiveness. Additionally, A2.3, A4C, C1.1, and C1.3 were specifically induced by MeJA.
Comprehensive expression profiling of LpHSF genes under abiotic stresses
-
The expression patterns of all 26 LpHSF genes were comprehensively profiled under various abiotic stresses, including heat, salt, cadmium (Cd), drought, and alkali treatments (Fig. 4). The qRT-PCR analysis under heat stress across four time points corroborated the transcriptomic data, establishing the reliability of the dataset. Beyond heat stress, the LpHSF family exhibited extensive transcriptional responsiveness to all other tested conditions.
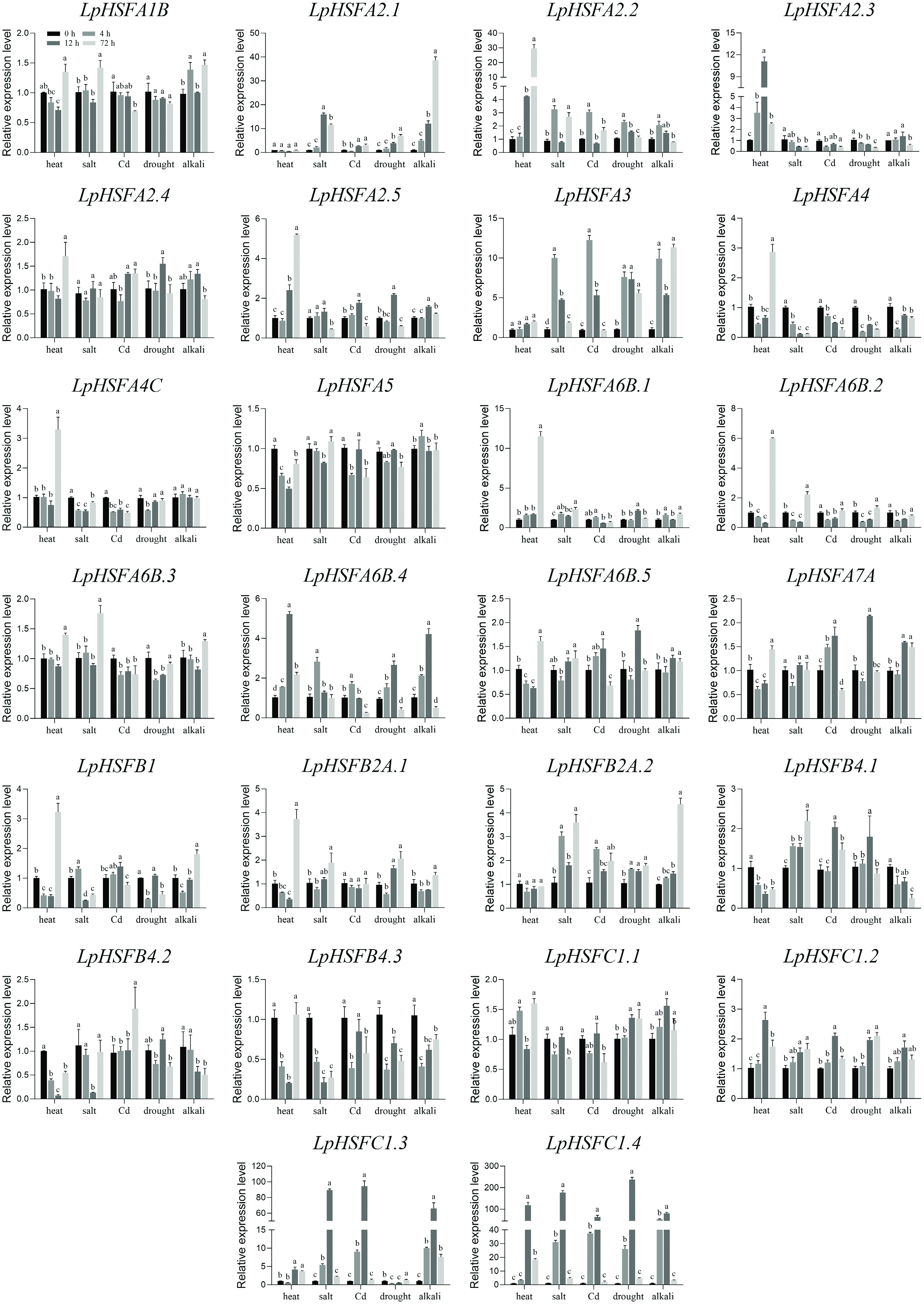
Figure 4.
Expression profiles of 26 LpHSF genes under heat, salt, Cd, drought, and alkali stress. The vertical bars indicate standard error. The statistical significance was determined using Fisher's protected least significant difference (LSD) test (p < 0.05).
Under salt stress, 15 LpHSFs were upregulated, comprising nine from subfamily A (A1B, A2.1, A2.2, A3, A6B.1-A6B.5), three from B (B2A.1, B2A.2, B4.1), and three from C (C1.2–C1.4). In contrast, five genes (A2.3, A4, A4C, B4.3, C1.1) were downregulated. Notable induction was observed for A2.1, A3, B2A.2, C1.3, and C1.4, which exhibited upregulation ranging from 3- to 200-fold, whereas A4 and B4.3 were the most significantly repressed.
Exposure to heavy metal (Cd) stress resulted in the upregulation of 13 LpHSFs, including A2.1, A2.2, A2.5, A3, and A6B.5 from subfamily A; B1, B2A.2, B4.1, and B4.2 from subfamily B; and C1.2, C1.3, and C1.4 from subfamily C. Conversely, ten genes were downregulated, including A1B, A2.3, A4, A4C, A5, A6B.1-A6B.3, B4.3, and C1.1. Marked induction was observed for A3, C1.3, and C1.4, with expression peaks elevated by 6- to 100-fold, while A4 was the most suppressed.
Under drought conditions, 15 genes were upregulated, including nine A-subfamily members (A2.1, A2.2, A2.4, A2.5, A3, A6B.1, A6B.4, A6B.5, A7A), three from the B-subfamily (B2A.1, B2A.2, B4.1), and three from the C-subfamily (C1.1, C1.2, C1.4). Eight genes were downregulated, including A2.3, A4, A4C, A5, A6B.2, A6B.3, B1, and B4.3. The expression of A2.1 and A3 was steadily induced, while C1.4 showed the most dramatic upregulation, and A2.3 was the most strongly repressed.
A distinct expression profile emerged under alkali stress, which triggered the most widespread upregulation, affecting 18 LpHSFs. These included 12 A-subfamily members (A1B, A2.1, A2.2, A2.3, A2.5, A3, A5, A6B.1, A6B.3, A6B.4, A6B.5, A7A), two from B-subfamily (B1, B2A.2), and all four C-subfamily members. Only five genes (A4, A6B.2, B4.1, B4.2, B4.3) were downregulated. The induction patterns of A2.1, A3, and C1.4 under alkali stress were notably consistent with those observed under drought.
-
Perennial ryegrass is a widely cultivated cool-season species valued both as forage and turfgrass. However, its productivity and survival are increasingly threatened by climate change-induced abiotic stresses, including extreme temperatures, salinity, alkalinity, and heavy metal toxicity. The heat shock transcription factor (HSF) family has been widely reported to play critical roles in plant responses to such stresses, positioning it as a key target for molecular breeding aimed at enhancing stress resilience[12,13,32]. Beyond their canonical role in thermotolerance, this study provides a comprehensive expression atlas of the HSF gene family in perennial ryegrass, revealing their functional diversity and tissue-specific regulation under multiple abiotic stresses, thereby underscoring their versatile roles in stress adaptation.
In this study, 26 LpHSF genes were identified from the perennial ryegrass genome. Promoter analysis revealed an abundance of hormone-responsive cis-elements, including those associated with abscisic acid (ABA), methyl jasmonate (MeJA), salicylic acid, auxin, and gibberellin. This composition is consistent with findings in Dianthus caryophyllus[33], Phaseolus vulgaris[34], and Triticum aestivum[35], suggesting an evolutionarily conserved role of HSFs in hormonal and abiotic stress signaling across plant species. The genome-wide identification and expression profiling further demonstrated that specific LpHSFs, particularly members of the LpHSFA2 subclass, LpHSFA3, LpHSFC1.3, and LpHSFC1.4, exhibit broad-spectrum stress responsiveness beyond heat stress. For instance, LpHSFA2.1 and LpHSFC1.3 were significantly upregulated under salt stress, mirroring the function of GmHSFB2b in soybean, which acts as a positive regulator of salt tolerance[36]. Similarly, the induction of multiple LpHSFs under cadmium stress aligns with the role of PvHSF16 in switchgrass as a positive regulator of cadmium tolerance[37]. Under heat stress, several Class A LpHSFs, including LpHSFA2.2, LpHSFA2.3, and LpHSFA4, were markedly upregulated, consistent with their established role as master regulators of the heat stress response. This functional parallel is observed in other species, such as TaHSFA2h in wheat[38] and OsHSFA4d in rice, which enhances thermotolerance by activating HSP101[39]. Thus, these heat-induced Class A LpHSFs likely serve as key positive regulators within the thermotolerance network of perennial ryegrass.
Class B HSFs have traditionally been regarded as transcriptional repressors[40], a view supported by their expression patterns under heat stress in this study. However, emerging evidence indicates functional divergence within this class. For example, ZmHSF21 in maize acts as a positive regulator of cold tolerance[41], suggesting that Class B HSFs may exert both positive and negative regulatory effects depending on the species and stress type. Among Class C members, LpHSFC1.4 responded to multiple stresses, including heat, salt, and alkali. In wheat, TaHSFC2a functions as a co-activator for Class A HSFs[38], whereas OsHSFC1a in rice acts by repressing a negative regulator[42]. This implies that stress-responsive Class C HSFs in ryegrass may operate either as transcriptional repressors or through the formation of heteromeric complexes with other HSFs. Collectively, the broad upregulation of LpHSFs under diverse stresses—such as salt, drought, alkali, and heavy metal exposure—aligns with recent studies in major crops including rice, maize, and wheat, underscoring the evolutionarily conserved role of HSFs as central regulators in plant abiotic stress networks.
Tissue-specific expression analysis revealed that most Class A LpHSFs were highly expressed in leaves, indicating a potential role in leaf development. In contrast, Class B and C members showed preferential expression in crowns and roots, suggesting specialized functions in below-ground organ development. Notably, many LpHSFs were also highly expressed in leaves across different developmental stages, implying a role in regulating leaf senescence. This observation is consistent with reports in rye (Secale cereale L.), where nearly half of the HSF members exhibit stage-specific expression[43]. In late-senescing barley genotypes, HSFs were significantly upregulated during late senescence stages, whereas no such enrichment was observed in early-senescing genotypes, indicating a positive correlation between HSF expression and leaf longevity[44]. Similarly, in bermudagrass, six HSFs were upregulated during dark-induced senescence, potentially facilitating the refolding of damaged proteins to maintain proteostasis[45]. In tall fescue, distinct functional specializations among HSF classes have been reported: Class A HSFs are rapidly induced under short-term heat stress and activate HSP expression to confer thermotolerance and delay heat-induced senescence; Class B members are also heat-induced but may participate in stress-senescence crosstalk through unclear mechanisms; and Class C HSFs are repressed by senescence, suggesting a role in modulating the antagonism between heat stress and senescence[46].
Previous studies have identified 16 HSF members in Italian ryegrass (Lolium multiflorum L.)[14]and 25 in perennial ryegrass[15]. While these studies primarily emphasized the role of HSFs in heat and drought responses, our time-course transcriptome analysis under heat stress revealed distinct temporal activation patterns among LpHSF members. Moreover, it was found that most LpHSFs are significantly induced by ABA and MeJA, highlighting their integration into hormone-mediated stress signaling pathways. Building on these findings, our promoter cis-element analysis elucidated the transcriptional regulatory basis of this response. The promoters were notably enriched with hormone-responsive motifs, particularly ABRE and MeJA elements. Strikingly, ABRE copy number exhibited a significant positive correlation with the level of heat-induced gene upregulation (r = 0.45, p < 0.05), functionally implicating this element in thermotolerance[47]. Similarly, the frequency of DRE elements correlated with transcriptional induction under both heat (r = 0.49, p < 0.05) and drought stress (Supplementary Table S4), suggesting its role as a signaling integrator[48]. Members such as LpHSFA2.1 and LpHSFA2.2, which harbor multiple ABRE and DRE copies, showed pronounced induction under saline-alkali and heat stress (Fig. 5). This establishes a direct link between specific cis-regulatory architectures and the observed multi-stress expression profiles of key LpHSF genes.
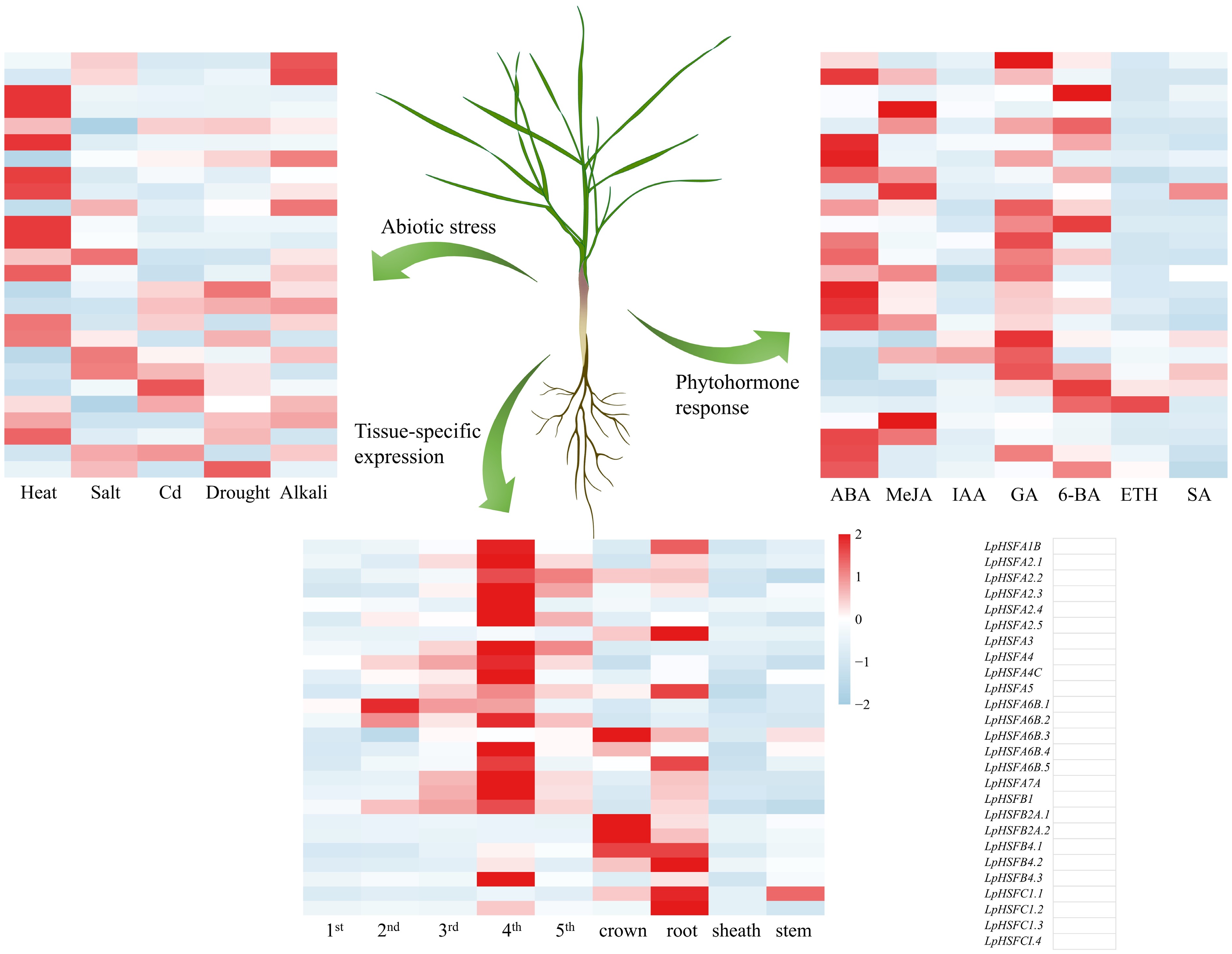
Figure 5.
The expression profiles of the LpHSF gene family in perennial ryegrass under different abiotic stresses and hormone induction, and different tissues.
In summary, the systematic expression profiling demonstrates that most LpHSF genes are transcriptionally regulated under diverse abiotic stresses and hormone treatments, reflecting the functional diversification of LpHSF proteins. This comprehensive analysis establishes a foundational resource for future investigations into the molecular mechanisms underlying stress adaptation in perennial ryegrass and supports the targeted selection of candidate genes for improving stress resilience through molecular breeding.
-
This study provides the first comprehensive expression atlas of the HSF gene family in perennial ryegrass (Fig. 5), revealing its functional diversification beyond heat stress. The study demonstrates that specific LpHSFs, particularly from classes A and C, are transcriptionally regulated by a broad spectrum of abiotic stresses and hormonal treatments, and exhibit distinct tissue-specific expression patterns suggestive of roles in leaf and root development as well as senescence. These findings highlight the evolutionarily conserved role of LpHSFs as a central regulatory hub in stress adaptation and establish a crucial foundation for future functional studies aimed at molecular breeding for enhanced multi-stress resilience in perennial ryegrass.
This study was supported by the National Natural Science Foundation Regional Innovation and Development (Grant No. U23A20218), the National Key Research and Development Program of China (Grant No. 2023YFF1001400), and the Sichuan Natural Science Foundation project (Grant No. 2023NSFSC0118), and the Sichuan Forage Innovation Team Program (Grant No. SCCXTD-2025-16).
-
The authors confirm their contributions to the paper as follows: study conception and design: Yu G; data collection: Li P, Zhou Y, Wang X; analysis and interpretation of results: Li P, Zhou Y, Wang X; draft manuscript preparation: Yu G, Li P; manuscript revision and review: Xie Z, Yu G, Zhang X; funding acquisition: Yu G, Zhang X. All authors reviewed the results and approved the final version of the manuscript.
-
All the LpHSF sequences were referenced in the NCBI genome (www.ncbi.nlm.nih.gov/datasets/genome/GCF_019359855.2/).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Guohui Yu, Peng Li
- Supplementary Table S1 Sequences of primers used for qRT-PCR analysis.
- Supplementary Table S2 Physicochemical properties and coding sequences of LpHSF proteins.
- Supplementary Table S3 Analysis of cis-elements in LpHSF gene promoters.
- Supplementary Table S4 Correlation analysis between the frequency of cis-elements and abiotic stress responses of LpHSF genes.
- Supplementary Fig. S1 Chromosomal distribution of 26 LpHSF genes.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu G, Li P, Zhou Y, Wang X, Xie Z, et al. 2026. Beyond heat stress: the heat shock factor family orchestrates multifaceted abiotic stress responses in perennial ryegrass. Grass Research 6: e004 doi: 10.48130/grares-0025-0037 

Beyond heat stress: the heat shock factor family orchestrates multifaceted abiotic stress responses in perennial ryegrass
- Received: 19 November 2025
- Revised: 18 December 2025
- Accepted: 23 December 2025
- Published online: 23 January 2026
Abstract: Perennial ryegrass (Lolium perenne L.) is a vital cool-season forage and turfgrass species whose productivity is increasingly threatened by climate change-induced abiotic stresses. The Heat Shock Factor (HSF) family are key transcription factors known to mediate plant stress responses, yet their functional landscape in perennial ryegrass remains poorly characterized, especially beyond heat stress. In this study, genome-wide identification revealed 26 LpHSF genes in perennial ryegrass, harboring abundant stress-responsive cis-elements. Moving beyond a singular focus on thermotolerance, the study established a comprehensive spatio-temporal expression atlas of LpHSFs. The findings demonstrate that specific LpHSFs, particularly members of the LpHSFA2 subclass, LpHSFA3, LpHSFC1.3, and LpHSFC1.4, are significantly upregulated under a broad spectrum of abiotic stresses, including salinity, alkalinity, and heavy metal exposure, indicating their roles as broad-spectrum stress regulators. Furthermore, this study uncovered distinct tissue-specific expression patterns, with Class A genes predominantly expressed in leaves and Classes B and C in roots and crowns, implicating their roles in organ development and stress adaptation. Notably, the expression of numerous LpHSFs during leaf senescence and their induction by hormones such as ABA and MeJA further highlights their functional diversity. This study provides the first integrated expression atlas of the LpHSF family, revealing their versatile roles in coordinating responses to diverse environmental and developmental signals. This work lays a crucial foundation for future functional studies and positions LpHSFs as prime targets for molecular breeding aimed at enhancing multi-stress resilience in perennial ryegrass.
-
Key words:
- Ryegrass /
- HSF /
- Abiotic stress /
- Phytohormone /
- Leaf senescence