









-
Platycodon grandiflorus (Jacq.) A. DC., commonly referred to as the balloon flower, is a perennial herb belonging to the family Campanulaceae and is indigenous to East Asia[1]. It is widely valued, not only as an ornamental plant, but also as a medicinal herb. Its ornamental appeal lies in its distinctive balloon-shaped buds that open into star-shaped flowers, available in various colours such as blue, purple, pink, and white. With a long blooming period from June through early autumn, it is popularly used in gardens, rockeries, and containers[2,3]. Meanwhile, the root of P. grandiflorus contains bioactive saponins with demonstrated efficacy against respiratory and inflammatory diseases[2]. In recent years, especially during the COVID-19 pandemic, its medicinal applications have attracted increasing attention, leading to growing demand[3].
Saponins, as the primary active components of P. grandiflorus, are important indicators for evaluating its medicinal quality[4]. The saponin skeleton of triterpenoid saponins is primarily synthesized through two pathways: the MVA and MEP pathways[5−7]. Several cytochrome P450 monooxygenases (CYP450) and glycosyltransferases (GT) catalyze the hydroxylation, oxidation, and glycosylation of the saponin skeleton to produce saponin monomers, including platycodin D, platycoside E, and platycodin D3[8,9]. Notably, saponin accumulation is subject to transcriptional regulation by phytohormonal elicitors. Among these, MeJA has emerged as a potent inducer of triterpenoid saponin biosynthesis in diverse medicinal species. Exogenous MeJA application can activate biosynthetic pathways by upregulating key genes involved in both early skeleton formation and late-stage modification steps, including CYP450s[10]. This established role makes MeJA induction a valuable experimental approach for probing the regulatory mechanisms underlying saponin production, and for identifying candidate biosynthetic genes. Cytochrome P450 is one of the classes of heme-containing B-type cytochrome superfamily proteins[11]. Through evolutionary expansion, it is widely distributed in cells of animals, plants, bacteria, and fungi[12]. Currently, more than 300,000 CYP450 genes, and over 50,000 CYP450 proteins have been discovered. They are divided into 11 clans, including four multi-family clans (CYP71, CYP72, CYP85, and CYP86), and seven single-family clans (CYP51, CYP74, CYP97, CYP710, CYP711, CYP727, and CYP746)[13].
CYP71 is a multi-family clan and contributes to approximately 50% of members in the plant CYP450 family[14]. It comprises several families that are heavily involved in the production of triterpenes, monoterpenes, and sesquiterpenes, including CYP71, CYP81-84, and CYP92[15,16]. The CYP71 gene family is the most abundant and functionally diverse family in the CYP71 clan[17]. Several members of the CYP71A, CYP71D, CYP71AR, and CYP76B subfamilies are involved in monoterpene metabolism[18]. Meanwhile, CYP71AV, CYP71BA, CYP71BL, CYP71D, and CYP71Z play a crucial role in sesquiterpene oxidation[19]. In addition to their involvement in the biosynthesis of monoterpenes and sesquiterpenes, genes from the CYP71 family participate in the biosynthesis of triterpene saponins. In Sapindus mukorossi, combined analysis of metabolites and transcriptome, and molecular biology experiments revealed that SmbHLH2, SmTCP4, and SmWRKY27 directly regulate the transcription of SmCYP71D-3 to affect the saponin content[20]. However, it is still unclear whether CYP71 genes in P. grandiflorus is involved in saponin biosynthesis.
The secondary metabolites of traditional Chinese medicinal plants are difficult to obtain through artificial synthesis due to their complex structures[21]. Inducing hairy roots using Agrobacterium rhizogenes-mediated transformation is considered a sustainable and effective method to study the biosynthesis of secondary metabolites in plants[22]. In addition, this method is proven to be effective in validating gene function, particularly in the case of regulatory key genes that play a role in specific biosynthetic pathways of secondary metabolites[23]. To date, this method has been successfully employed in various plants, including Panax ginseng, Salvia miltiorrhiza, and Astragalus membranaceus[24,25]. However, it requires tissue culture, increasing the difficulty and cost of operation. To solve this problem, the cut-dip-budding (CBD) delivery system is established, which can generate transformed roots and transformed buds under non-sterile conditions, and without the need for tissue culture. This approach has been successfully applied to various plants, including Aralia elata and Coronilla varia[26]. However, there have been no studies using this method to obtain transgenic roots in P. grandiflorus.
Although CYP450 genes have been identified and functionally verified in various plant materials, the identification of the CYP450 gene family in P. grandiflorus has not been conducted. Nine clans were formed from the identification of 175 PgCYP450s genes in P. grandiflorus. The expression patterns of 175 PgCYP450s genes under MeJA treatment were then investigated, and a hairy root transformation system was used to confirm PgCYP71BE217 gene's function. This study offers valuable insights into the expression dynamics of the CYP450 gene family in response to MeJA induction, therefore establishing a foundation for future investigations into the roles of CYP450 genes in saponin biosynthesis.
-
The National Genomics Data Center (
https://ngdc.cncb.ac.cn/gwh ) provided the genome data for P. grandiflorus[27]. First, potential PgCYP450s in the genome of P. grandiflorus were found using the hidden Markov model profile of the p450 domain (PF00067) as a reference file. Second, the members of the CYP450 gene family in P. grandiflorus were identified using a BLASTP search with query sequences derived from Arabidopsis thaliana CYP450 genes. Ultimately, the P450 nomenclature committee (http://drnelson.uthsc.edu/CytochromeP450.html ) received the acquired amino acid sequences of the PgCYP450s to assign nomenclature. For a thorough understanding, the ExPasy website (https://web.expasy.org/protparam/ ) was used to study the properties of the PgCYP450 sequences, including their amino acid length, molecular weight (MW), isoelectric point (pI), and other pertinent data. The phylogenetic tree with 1,000 bootstrap repetitions was built using the neighbor-joining (NJ) method in MEGA7.0 software. Based on the genomic annotation of the P. grandiflorus genome, the chromosomal position map was plotted using TBtools software[28].Analysis of phylogenetic tree, cis-acting elements, and conserved domains of the PgCYP71s
-
The motif patterns of the PgCYP71s protein sequences were examined using the MEME website (
https://meme-suite.org/tools/meme ). The nmotif parameter was set to 10; the minw parameter was set to 6, and the maxw parameter was set to 50. The ggtree platform was used to illustrate the conserved domains and phylogenetic tree of PgCYP71s. TBtools was employed for the extraction of a 2,000 bp sequence upstream of the start codon for every PgCYP450 member. The PlantCARE website (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) was used to identify the cis-acting elements in the promoter region.Treatment of P. grandiflorus roots with MeJA
-
P. grandiflorus seedlings were cultured uniformly on 1/2 MS medium (pH 5.8) supplemented with 100 μM MeJA and 30 g/L sucrose, and 5 g/L agar[5]. Control groups received equivalent volumes of dimethyl sulfoxide. MeJA-treated samples were harvested at 0, 12, 24, and 48 h post-exposure.
qRT-PCR assay
-
Total RNA was isolated from P. grandiflorus tissues and MeJA-treated roots using TransZol UP (TransGen Biotech, Beijing, China). cDNA synthesis employed the SuperMix kit for qRT-PCR (TransGen Biotech). Gene expression quantification utilized the BIO-RAD CFX96 Touch system (Hercules, CA, USA), with 18S rRNA as the endogenous reference (Supplementary Table S1).
Establishment of a genetic transformation system for hairy roots
-
A hairy root system was developed for P. grandiflorus to speed up the evaluation of gene function. The TY solid medium (50 mg/L kanamycin) was used to cultivate Agrobacterium rhizogenes K599 transformed with the empty vector pBI121 for 36–48 h. A 5 mL of TY liquid medium was used to incubate a few selected single colonies for the entire night. A 100 μL culture was then added to 50 mL of TY liquid that had been treated with 200 μmol/L AS and 50 mg/L kanamycin. Centrifugation (for 10 min at 4,000 rpm) was used to pellet the cells once they reached an OD600 ≈ 0.8. They were then resuspended in activation buffer (200 μmol/L AS, 10 mmol/L MES, and 10 mmol/L MgCl2), and dark-incubated for two hours at room temperature. The activated solution was used to infect root-free sterile 30-day-old seedlings via vacuum infiltration[5]. After 60 d, transformed plants were moved to sterile soil, and positive transformants were found for target gene expression and saponin measurement.
Subcellular localization and self-activation of PgCYP71BE217 in yeast
-
Cloning the PgCYP71BE217 coding gene without a termination codon into the pCAMBIA1300-GFP BamHI/KpnI sites. In tobacco epidermal cells, the recombinant construct (pCAMBIA1300-PgCYP71BE217-GFP), and the empty vector were both simultaneously expressed. Subcellular localization was analyzed using confocal microscopy (Olympus FV3000; 488 nm excitation). For yeast two-hybrid assays, PgCYP71BE217 CDS was fused to pGBKT7 and transformed into the Y2H Gold strain. Self-activation testing followed established protocols[29].
Analysis of the function of PgCYP71BE217 in the hairy root of P. grandiflorus
-
The recombinant pCAMBIA1300-PgCYP71BE217-GFP construct was introduced into Agrobacterium rhizogenes K599 for P. grandiflorus transformation. Transgenic hairy roots were initially screened via GFP fluorescence detection using a chemiluminescence imaging system (Tanon 5200, Shanghai, China). Subsequently, genomic DNA from positive lines was amplified with GFP-specific primers. Lastly, the transcript levels of PgCYP71BE217 and related pathway genes were measured using RT-qPCR assay (Supplementary Table S1), while HPLC assessed platycodin D, platycoside E, and platycodin D3 contents as described previously[30].
-
In total, 175 PgCYP450s genes with conserved p450 domains were identified in P. grandiflorus. The P450 nomenclature committee received the protein sequences of 175 CYP450s to provide nomenclature. Their characteristics, including amino acid length, MW, and pI, are listed in Supplementary Table S2. The amino acid number varied from 219 (PgCYP716S4) to 940 (PgCYP71AT281); MW varied from 24.89 kDa (PgCYP716S4) to 107.32 kDa (PgCYP71AT281), and pI varied from 5.79 (PgCYP71AT281) to 9.41 (PgCYP72A1005). Phylogenetic analysis divided PgCYP450s into nine clans (Fig. 1), including five single-family, and four multi-family clans. Among the nine clans, the clan with the largest number of members was the CYP71 clan, which consisted of 78 members, with the CYP71 family containing the highest members (18), followed by the CYP85 (30), CYP72 (29), and CYP86 (17) clans. The remaining clans exclusively consisted of single-family clans, with the CYP710 clan having the highest number of members.
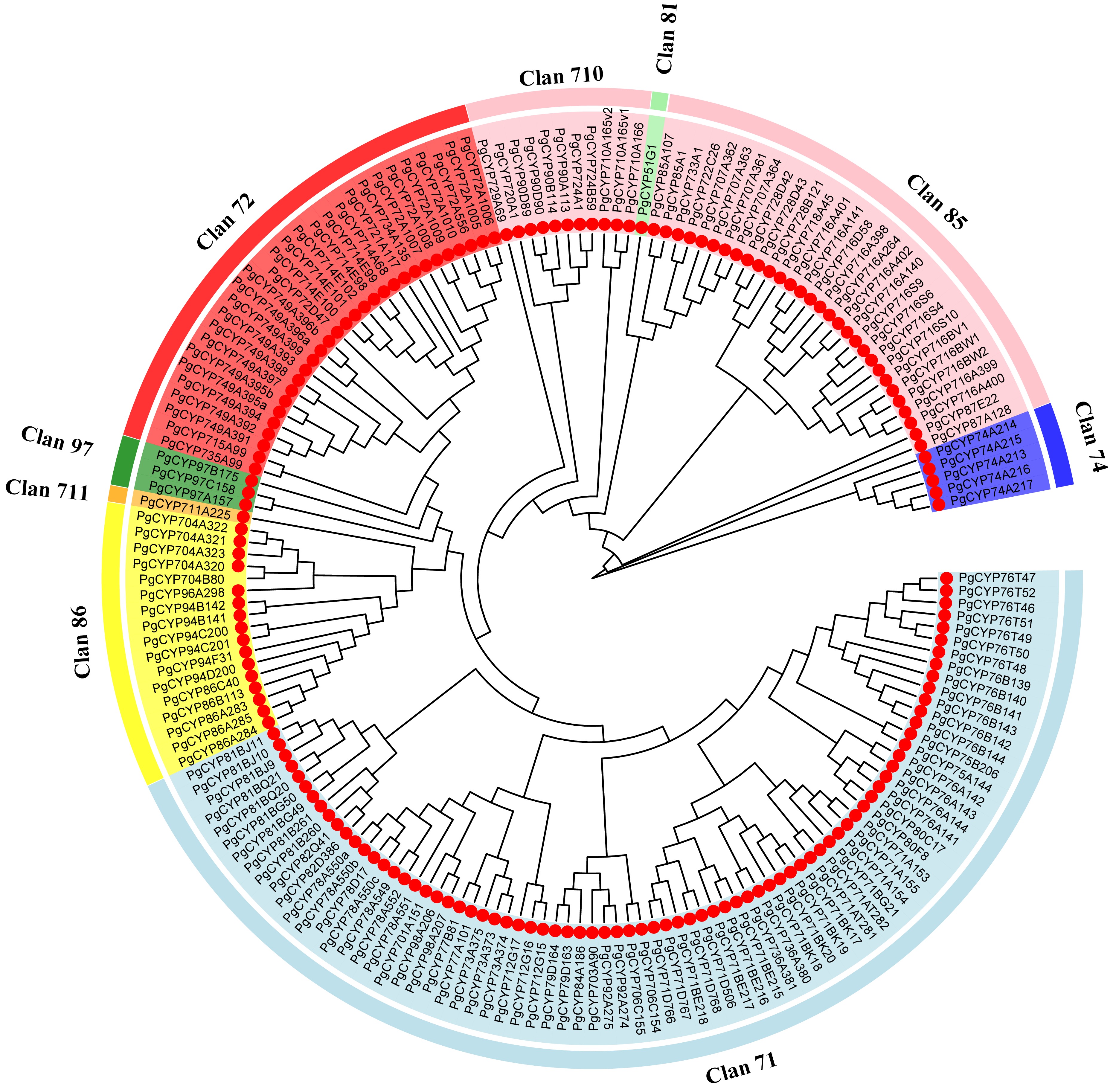
Figure 1.
Phylogenetic relationships of the Platycodon grandiflorus CYP450 proteins. A neighbor-joining phylogenetic tree was generated based on the alignment of full-length amino acid sequences of the 175 PgCYP450 proteins in P. grandiflorus.
Phylogenetic analysis and analyses of conserved domains and cis-acting elements of PgCYP71 genes
-
MeJA has been demonstrated to enhance the accumulation of triterpenoid saponins[5]. MeJA significantly increased the expression level of the PgCYP71s gene in the PgCYP450 family, according to an analysis of transcriptome data from previously published studies (Supplementary Fig. S1)[5]. This finding suggested that PgCYP71s may play a role in the MeJA-mediated biosynthetic of triterpenoid saponins. Moreover, 18 PgCYP71s were determined in P. grandiflorus, which were divided into four subfamilies, namely, CYP71As, CYP71Ats, CYP71Ds, and CYP71BEs. Analysis of the conserved domains of PgCYP71 genes revealed that most genes had 9–10 motifs, except the PgCYP71BG21 gene, exhibiting five motifs (Fig. 2). Promoter cis-acting element analysis revealed that PgCYP71 genes are mainly involved in growth and development, light response, plant hormone signaling, and stress response. Among these genes, PgCYP71BE217 contained the highest number of plant hormone signaling elements (Fig. 3). These results revealed that MeJA appears to regulate saponin accumulation by inducing PgCYP71s expression.
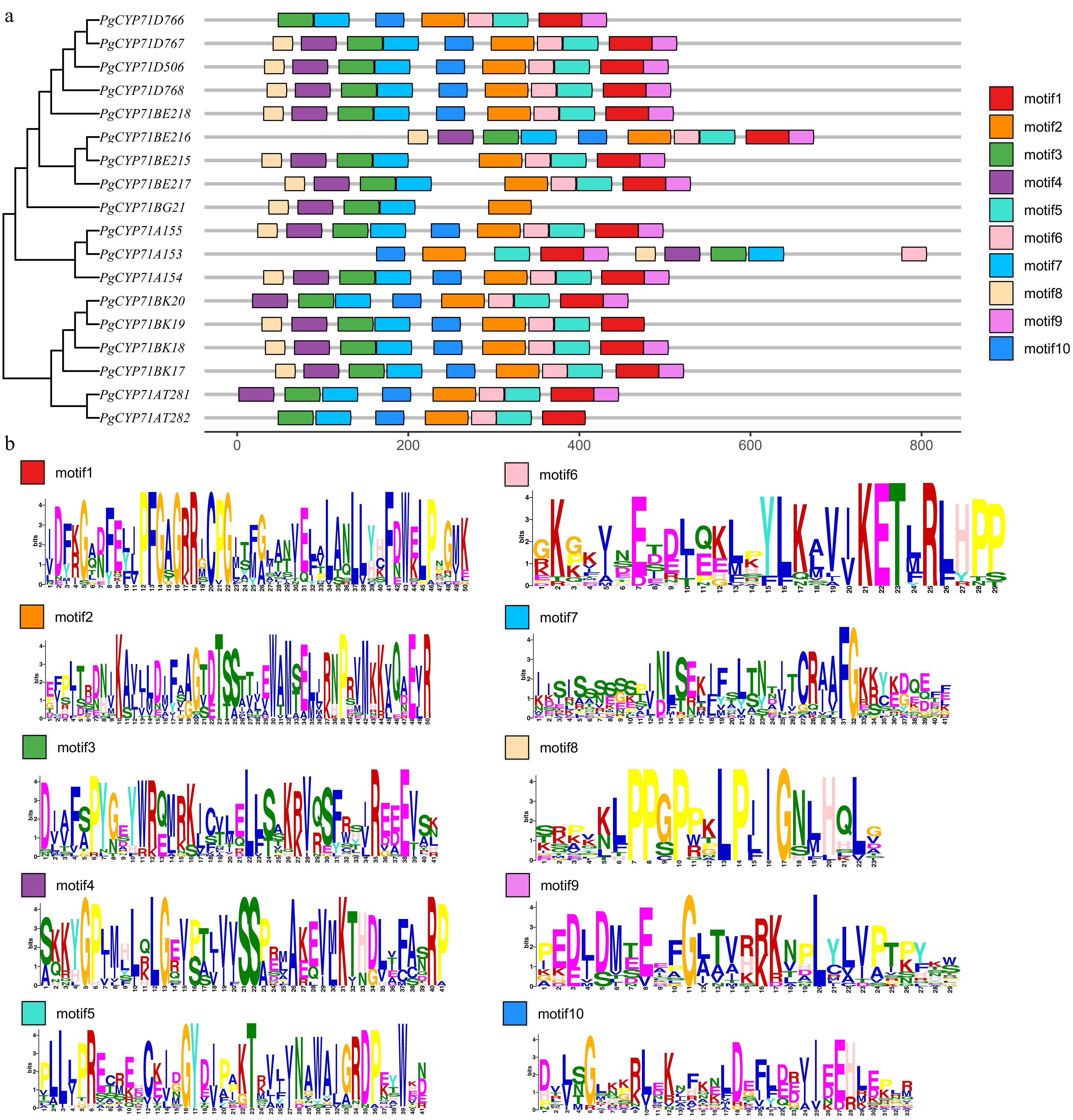
Figure 2.
Phylogenetic relationships and conserved motifs of P. grandiflorus PgCYP71 proteins. 18 PgCYP71s in P. grandiflorus have their full-length amino acid sequences aligned to generate a neighbor-joining phylogenetic tree. (a) Phylogenetic tree and onserved motifs distribution of PgCYP71 proteins. (b) Composition of the conserved motifs in PgCYP71 proteins.
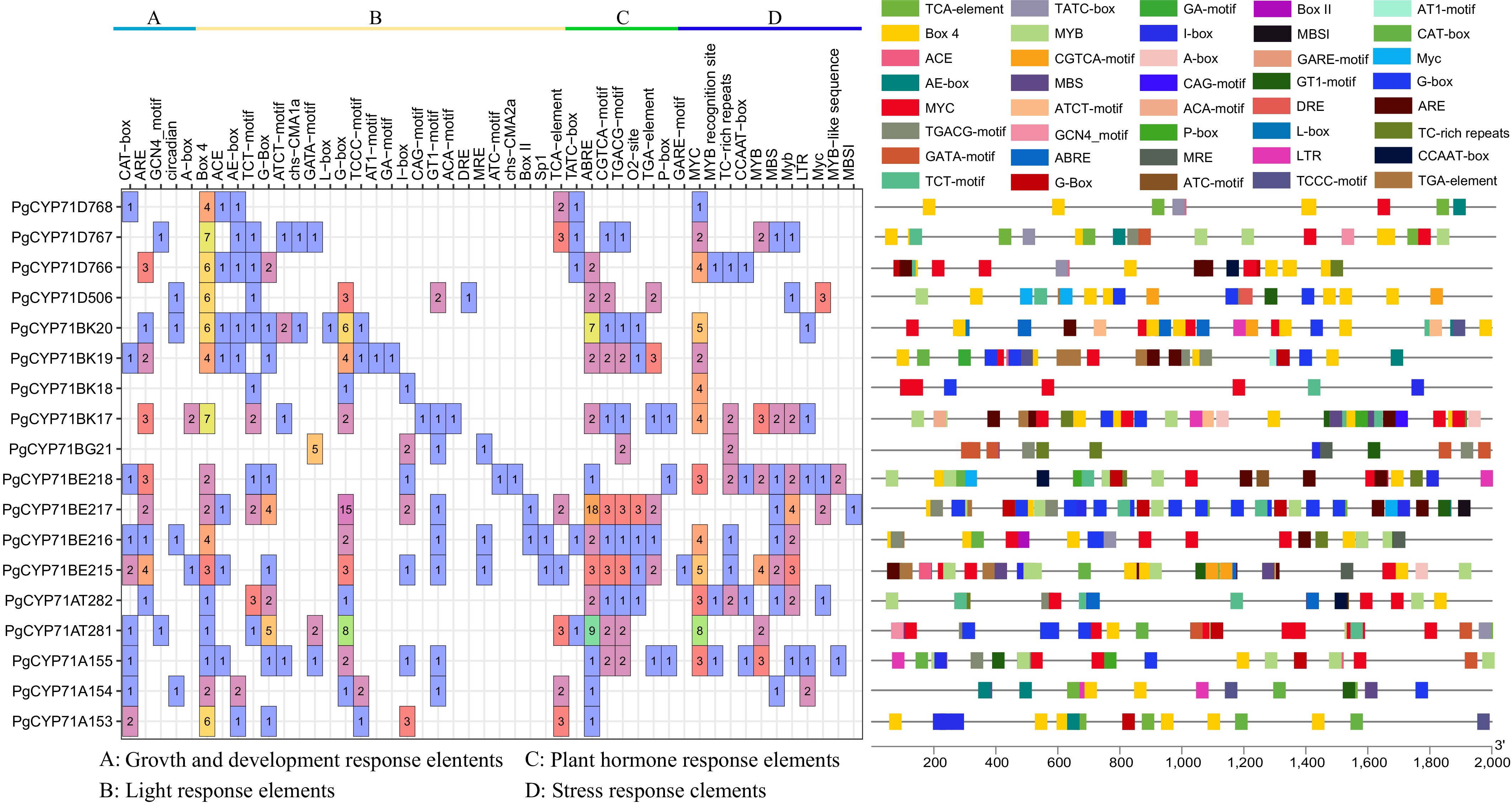
Figure 3.
Cis-acting elements in the promoter regions of PgCYP71 genes. Left - Distribution and functional classification of cis-acting elements identified in each PgCYP71 gene. Right - Visualization of major cis-acting elements, with each category represented by a distinct color.
qRT-PCR analysis of the expression patterns of PgCYP71s under MeJA treatment
-
MeJA treatment could increase the total saponin and monomeric saponin contents in the root of P. grandiflorus[5,31]. The expression pattern of the PgCYP71 gene family was analyzed using RT-qPCR. It can be loosely classified into three different classes on the basis of expression trend. MeJA upregulated PgCYP71BK20 and PgCYP71BE217, which were included in the first category. The second category included PgCYP71A153 and PgCYP71D506, which were inhibited by MeJA. PgCYP71AT281 and PgCYP71BE218 were included in the third category; their expression levels first decreased, then increased, and finally decreased (Fig. 4). Compared with the control group, PgCYP71BE217 exhibited upregulation by more than 60-fold under MeJA treatment at 12 h, implicating it as a pivotal MeJA-responsive regulator of saponin biosynthesis.
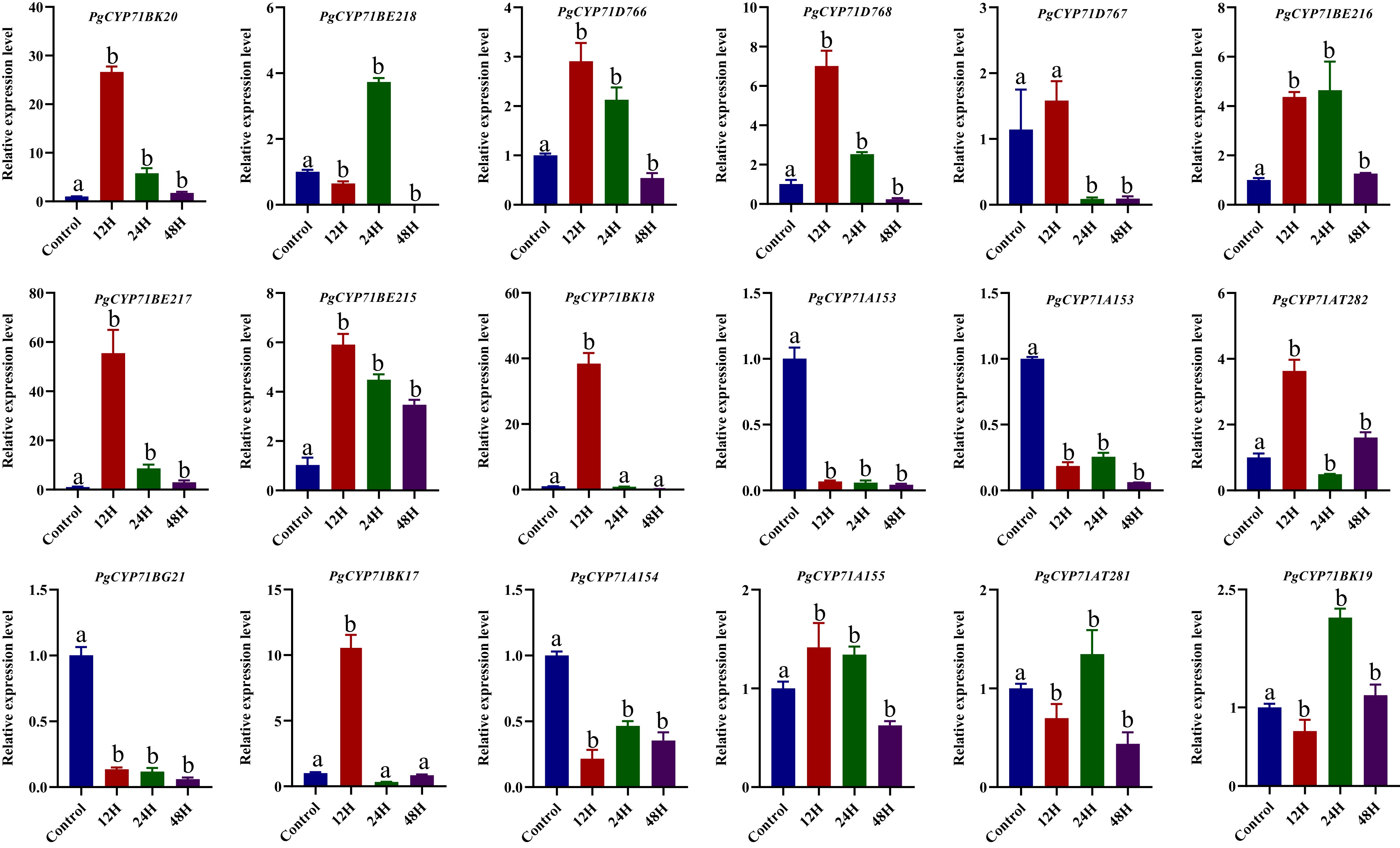
Figure 4.
The qRT-PCR has shown the expression pattern of the PgCYP71 gene family at 0 (control), 12, 24, and 48 h after 100 μmol MeJA treatment. Different letters indicate significant differences (p < 0.05).
Expression analysis and self-activation of PgCYP71BE217 in yeast
-
The 504 amino acid-encoding CDS section of the PgCYP71BE217 gene was cloned. The presence of p450 domains in this gene was discovered through multiple sequence alignment (Fig. 5a). Subcellular localization analysis in tobacco leaf epidermal cells revealed that PgCYP71BE217-GFP fluorescence was predominantly observed at the plasma membrane and in the cytoplasm (Fig. 5b). PgCYP71BE217 did not exhibit self-activation activity (Fig. 5c). Moreover, tissue-specific analysis revealed that the PgCYP71BE217 gene was highly expressed specifically in the roots (Fig. 5d, e).
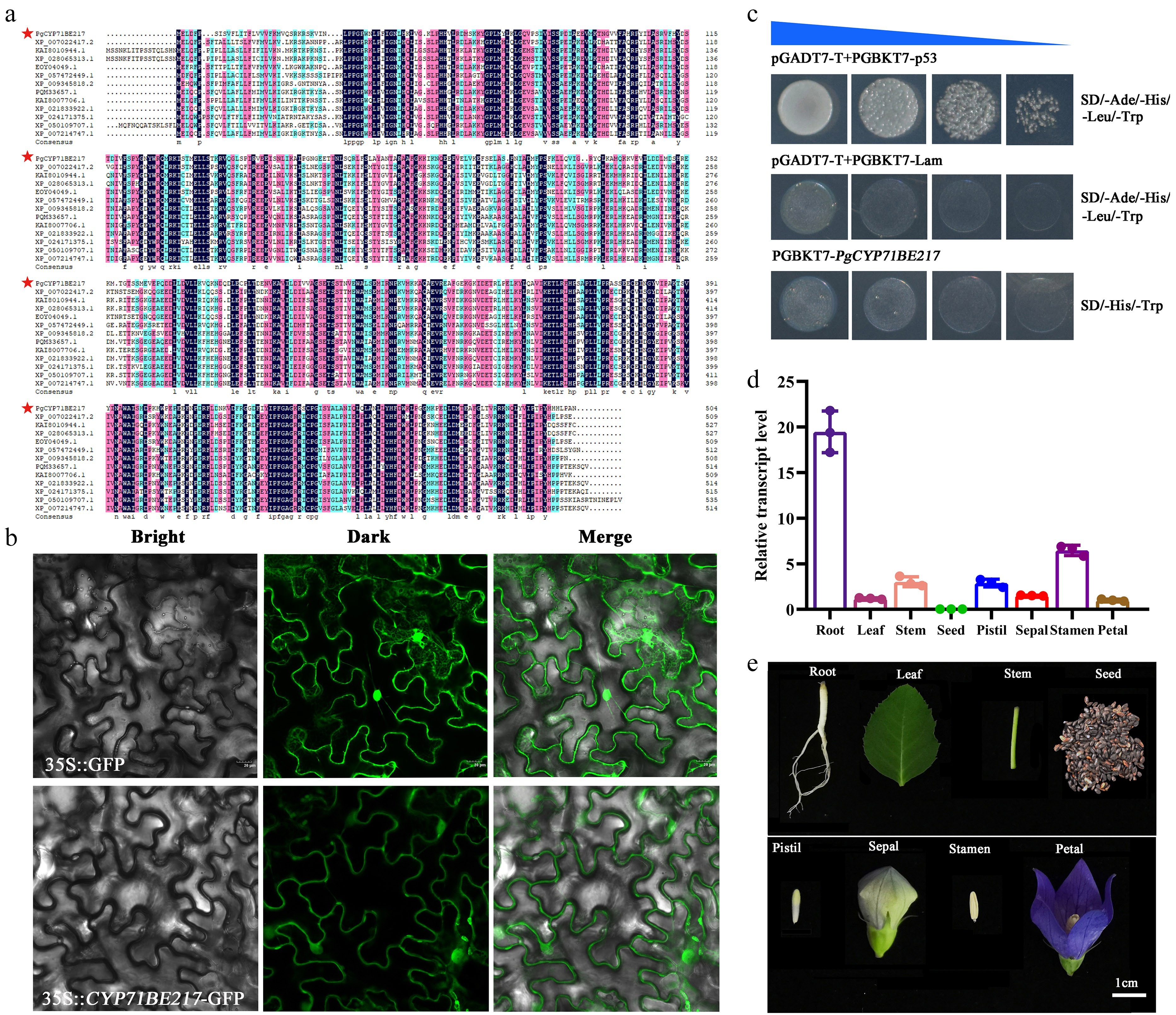
Figure 5.
PgCYP71BE217 expression patterns in P. grandiflorus. (a) PgCYP71BE217 multiple sequence alignment. (b) Subcellular localization of PgCYP71BE217. PgCYP71BE217 was primarily seen at the plasma membrane and in the cytoplasm; (Top) subcellular localization of the empty pCAMBIA1300-GFP vector used as the positive control; (Bottom) Bright field, dark field, and merged field are represented by the left, center, and right panels, respectively. (c) Evaluation of self-activation of PgCYP71BE217. (d) The expression level of PgCYP71BE217 in eight different tissues as revealed by qRT-PCR. (e) The eight different tissues of P. grandiflorus subjected to qRT-PCR of (d).
PgCYP71BE217 overexpression enhanced P. grandiflorus's saponin content
-
P. grandiflorus was effectively used to develop a hairy root transformation system, and positive transgenic hairy roots with an 82% transformation efficiency were produced (Fig. 6). PgCYP71BE217 was successfully introduced into P. grandiflorus (Fig. 7a, b). PgCYP71BE217 expression levels in OE1, OE7, and OE12 were substantially higher than those in the wild type (Fig. 7c). Moreover, the expression levels of PgHMGS, PgbAS1, and PgSS (responsible for the biosynthsis of saponin) were significantly elevated compared with wild type (Fig. 7d). According to HPLC examination, compared to the roots of wild-type plants, the roots overexpressing PgCYP71BE217 had significantly larger concentrations of platycoside E, platycodin D3, and platycodin D (Fig. 7e). These results clearly demonstrated that PgCYP71BE217 participated in saponin biosynthesis.
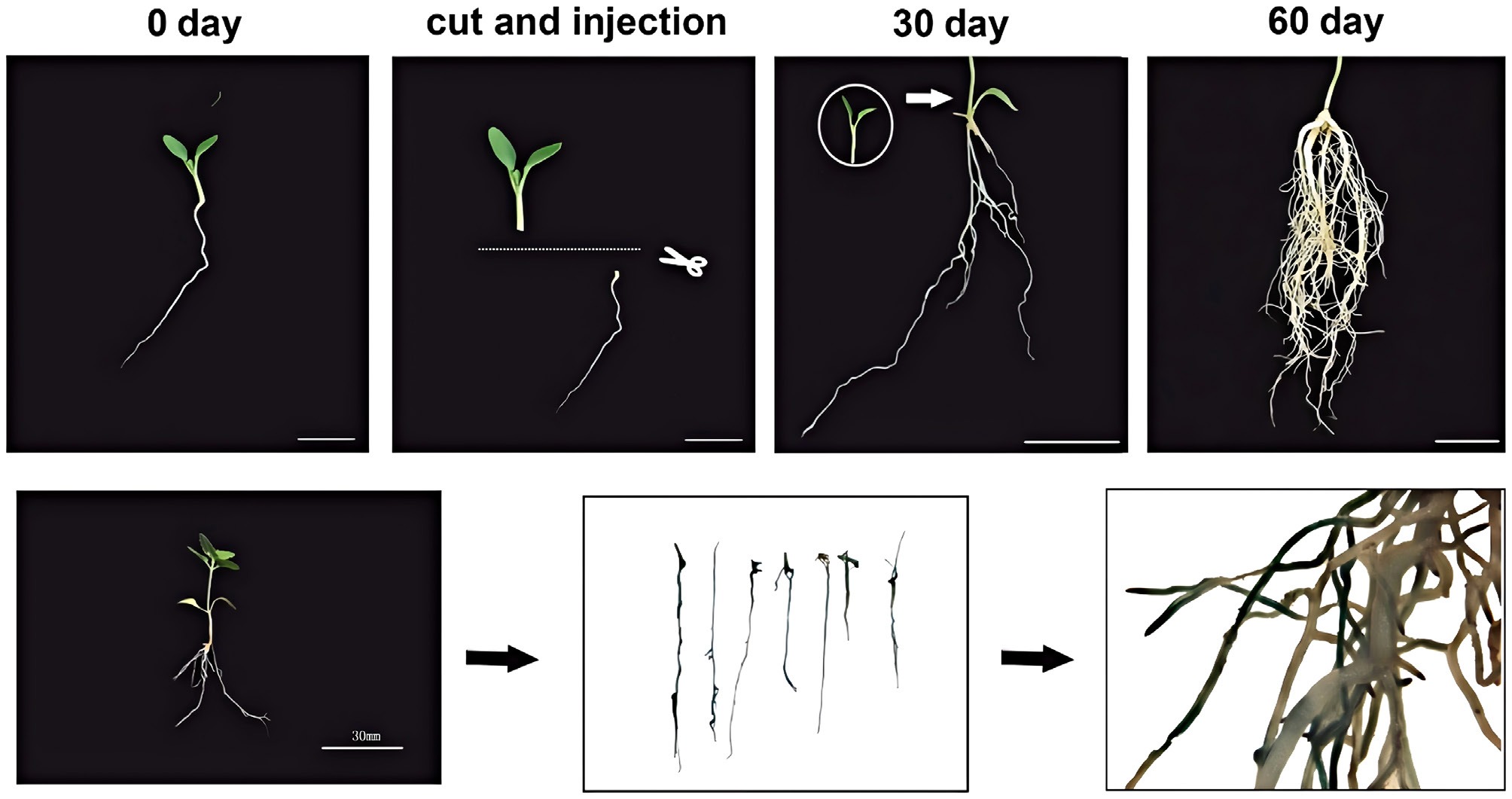
Figure 6.
Establishment of hairy root transformation system and GUS validation. Top - Schematic diagram illustrating the procedure for hairy root transformation. Bottom - GUS assay results confirming transgenic hairy roots.
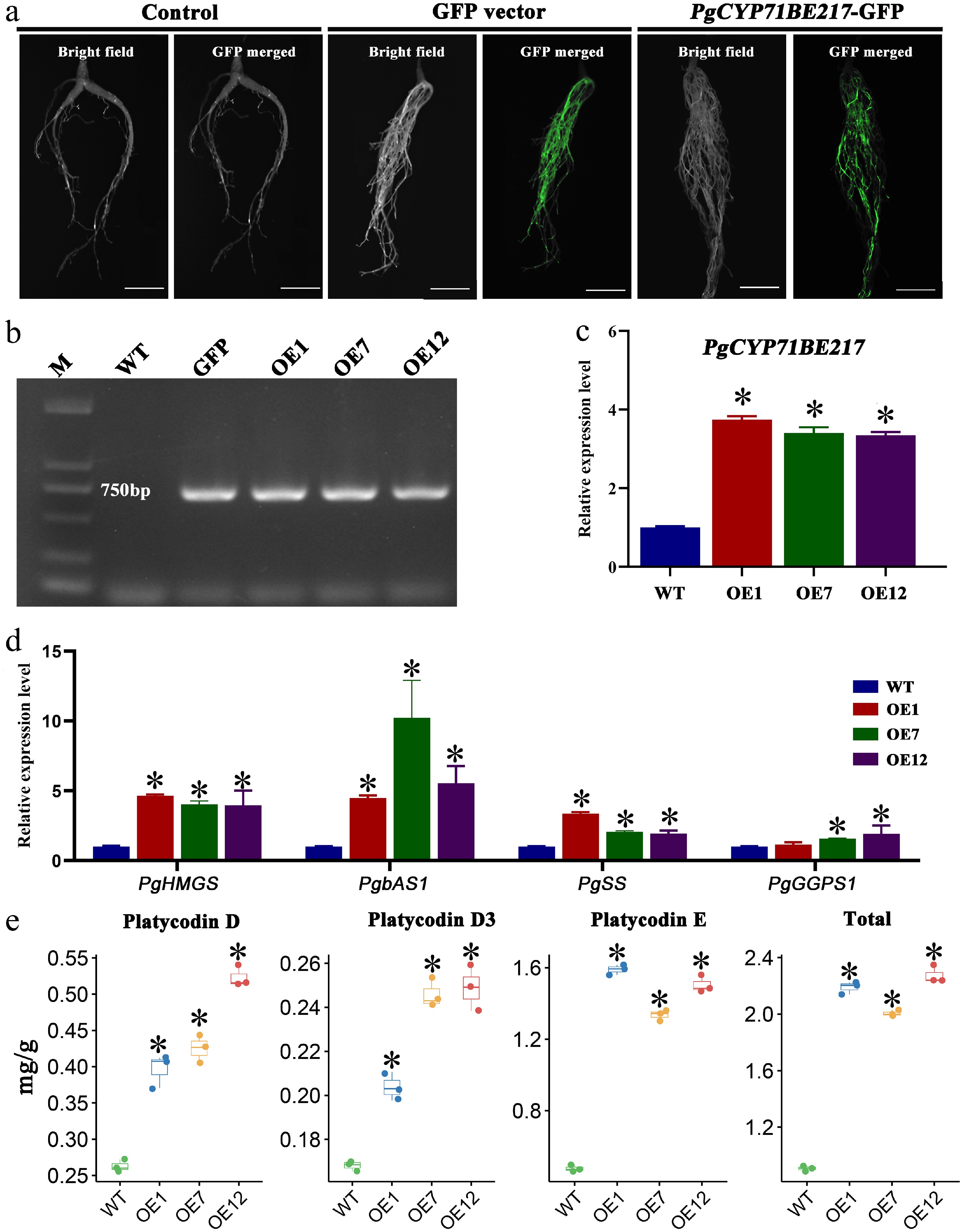
Figure 7.
Functional verification of PgCYP71BE217. (a) Phenotypic characterization of PgCYP71BE217-OE transgenic hairy roots using GFP detection. (b) DNA verification of PgCYP71BE217-OE vector. (c) qRT-PCR analysis of PgCYP71BE217 expression in transgenic and wild-type hairy roots. (d) The expression levels of PgHMGS, PgbAS1, and PgSS were significantly increased in the transgenic hairy roots of plants with PgCYP71BE217 overexpression. (e) Changes in saponin content after PgCYP71BE217 overexpression. The asterisk denotes a statistically significant difference compared to the WT (p < 0.05).
-
In recent years, with the development of genome sequencing technology and bioinformatics, researchers could identify CYP450 genes from 66 plant species, including A. thaliana, Solanum lycopersicum, and Zea mays[32]. The model plant A. thaliana has 286 CYP450 genes, which can be categorized into nine clans consisting of 45 families, and 72 subfamilies[33]. A total of 175 PgCYP450s genes were found in this investigation. According to evolutionary analysis, the PgCYP450 genes could be divided into nine clans: five single gene family clans (CYP51, CYP74, CYP97, CYP710, and CYP711), and four multiple gene family clans (CYP71, CYP72, CYP85, and CYP86). The CYP727 and CYP746 clans were not present. This is consistent with the majority of dicotyledonous plants[16].
PgCYP71BE217 controls P. grandiflorus's production of triterpenoid saponins
-
Among the key inducers that control plant growth and secondary metabolite synthesis is MeJA. Furthermore, it is crucial in controlling the production of saponins[34]. RNA-seq analysis revealed that PgCYP71s are induced by MeJA (Supplementary Fig. S1). Several plant hormone-responsive elements, including MeJA cis-acting elements, were present in the promoter of PgCYP71 genes (Fig. 3). PgCYP71BE217 was significantly induced by MeJA as revealed by qRT-PCR (Fig. 4). These data demonstrated that PgCYP71BE217 is involved in regulating the synthesis of plant signaling molecules, which is consistent with the observations in Ginkgo biloba and P. ginseng[35,36]. In addition to the response elements for MeJA, the promoter of this gene contained response elements for other hormones such as abscisic acid and gibberellins, suggesting that PgCYP71BE217 is regulated by other hormones (Fig. 3)[37].
The CYP71 clan, which was often considered the cradle for the synthesis of monoterpenes and sesquiterpenes, participates in the biosynthesis of diterpenes and triterpenes[38,39]. In Lotus japonicus, CYP71D353 belonged to the CYP71 clan, which was significantly upregulated in the roots, and participated in triterpene biosynthesis[38]. In P. grandiflorus, PgCYP71BE217 overexpression in transformed hairy roots improved the content of saponins, indicating that PgCYP71BE217 participated in saponin biosynthesis (Fig. 7). These data demonstrated that the CYP71 clan in P. grandiflorus was involved in saponin biosynthesis. Our subsequent studies will be focused on functional validation of other potential genes in this clan.
Transcription factors specifically bind to DNA sequences to regulate transcription. Through attaching themselves to the promoters of structural genes, they might control the production of triterpenoid saponins. The promoter of PgCYP71BE217 contained numerous transcription factor binding sites, such as AP2/ERF[40], bHLH[41], and WRKY transcription factor families[42]. This indicated that PgCYP71BE217 may be regulated by these transcription factors to regulate saponin accumulation (Fig. 3; Supplementary Table S3).
Rapid analysis of gene function using the hairy root transformation system
-
Transgenic plants are typically obtained using two methods: Agrobacterium-mediated transformation or the gene gun. However, both methods have low transformation efficiency and require tissue culture[26]. Tissue culture is a significant obstacle for obtaining transgenic plants of species that are difficult or impossible to genetically transform. Hence, the development of an efficient method that does not rely on tissue culture is currently a prominent area of research[43].
For medicinal plants, the transformation of hairy roots is a fast and stable method, characterized by genetic stability, rapid growth, and easy cultivation. Related extensive research has been conducted on medicinal plants such as Catharanthus roseus and P. ginseng[44,45]. However, it requires tissue culture, increasing the difficulty and cost of operation. To reduce the complexity of the process and develop a transformation system that does not rely on tissue culture, a tissue-culture-independent hairy root transformation system was established (Fig. 6). The establishment of this method will provide technical support for the rapid analysis of gene function in P. grandiflorus.
-
In this study, 175 PgCYP450 genes in P. grandiflorus were identified. They were divided into nine clans, excluding the CYP727 and CYP746 clans. Saponin accumulated after MeJA treatment, and PgCYP71BE217 was clearly stimulated in roots, suggesting that PgCYP71BE217 may be involved in the production of saponin in roots. The PgCYP71BE217 overexpression may promote saponin accumulation in P. grandiflorus, according to the hairy root transformation. Furthermore, novel technical assistance for quick evaluation of gene function in P. grandiflorus was made possible by the development of a hairy root transformation system that does not require tissue culture.
This work was supported by the Heilongjiang Science and Technology Talent 'Spring Goose' Support Program of Heilongjiang Province of China (Grant No. CYQN24018), and Natural Science Foundation of Heilongjiang Province (Grant No. 2024C012). We also thank MJEditor (www.mjeditor.com) for providing English editing services during the preparation of this manuscript.
-
The authors confirm their contributions to the paper as follows: investigation, data curation, formal analysis, writing − original draft, software: Zhang W, Li P; resources: Dong J; visualization: Che D; software: Song Y; visualization, conceptualization, methodology: Yang T, Zhang J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Wuhua Zhang, Pinghui Li
- Supplementary Table S1 List of all primers.
- Supplementary Table S2 Information on the CYP450 gene family in P. grandiflorus.
- Supplementary Table S3 Prediction of transcription factor binding sites in the promoter of the PgCYP71s genes.
- Supplementary Fig. S1 The expression pattern of PgCYP450s under MeJA treatment based on RNA-seq data.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang W, Li P, Dong J, Song Y, Che D, et al. 2026. Comprehensive genome-wide identification of PgCYP450 genes and functional characterization of PgCYP71BE217 in saponin biosynthesis of Platycodon grandiflorus. Ornamental Plant Research 6: e005 doi: 10.48130/opr-0025-0048 

Comprehensive genome-wide identification of PgCYP450 genes and functional characterization of PgCYP71BE217 in saponin biosynthesis of Platycodon grandiflorus
- Received: 13 August 2025
- Revised: 07 October 2025
- Accepted: 11 October 2025
- Published online: 02 February 2026
Abstract: Platycodon grandiflorus (Jacq.) A. DC., a well-known traditional Chinese medicinal herb, is particularly abundant in saponins. The cytochrome P450 monooxygenase (CYP450) superfamily constitutes the key group of enzymes responsible for regulating saponin biosynthesis. To date, a comprehensive analysis of CYP450 genes associated with saponin biosynthesis in P. grandiflorus has not been conducted. In the present study, 175 PgCYP450 genes were systematically identified in P. grandiflorus. Phylogenetic analysis classified these genes into nine distinct clans. Transcriptome (RNA-seq) data further demonstrated that the majority of genes within the CYP71 family were upregulated in response to methyl jasmonate (MeJA) treatment. qRT-PCR revealed that PgCYP71BE217 had the highest expression level after MeJA induction, which was over 60 times higher than the control, and displayed elevated expression primarily in the roots. Subcellular localization and yeast self-activation experiments indicated that PgCYP71BE217 was localized to both the plasma membrane and cytoplasm, and lacked self-activation activity. Overexpression of PgCYP71BE217 using a hairy root transformation system enhanced the transcript levels of PgHMGS (3-hydroxy-3-methylglutaryl-CoA synthase), PgbAS1 (β-amyrin synthase 1), and PgSS (squalene synthase), while simultaneously elevating the accumulation of saponins such as platycodin D, platycoside E, and platycodin D3. The identification and functional analysis of the CYP450 gene family in this study enhances understanding of saponin synthesis and provides valuable genetic resources for P. grandiflorus breeding.