






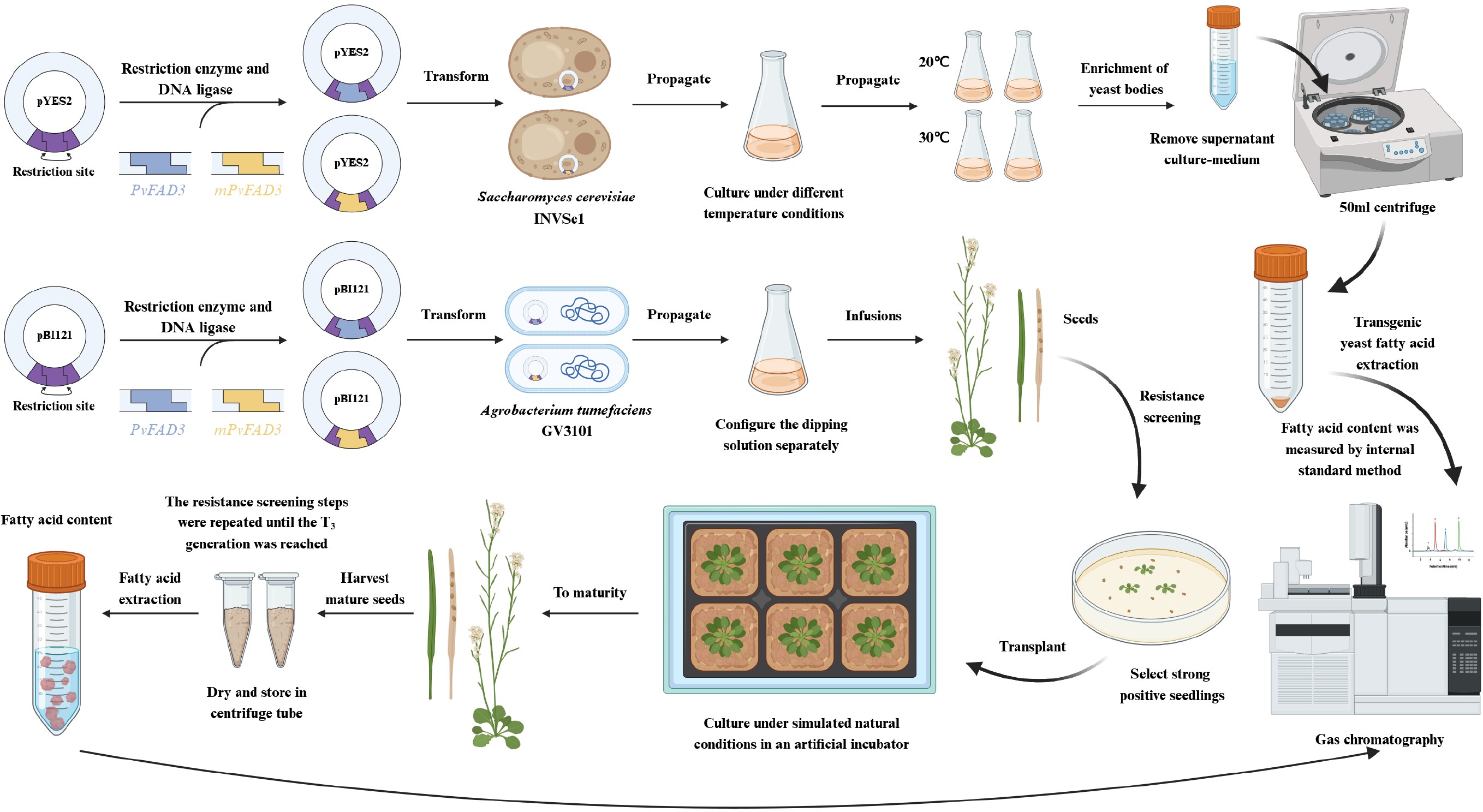



-
Plukenetia volubilis, belonging to the Euphorbiaceae family, is a perennial woody liana renowned for its significant economic value. Cultivated as an oilseed crop in South America[1,2], it yields approximately 1,200 tons of seeds annually in Peru[3]. In China, P. volubilis is recognized as an emerging economic crop, cultivated extensively in subtropical and tropical regions such as Guangdong, Yunnan, and Hainan provinces[4]. Mature P. volubilis seeds (PVS) contain approximately 50% oil by weight[5]. The predominant fatty acids (FAs) are palmitic acid (C16:0), stearic acid (C18:0), oleic acid (C18:1), linoleic acid (C18:2, LA), and linolenic acid (C18:3, ALA)[6]. Notably, the essential FAs of LA and ALA constitute approximately 25% and 50% of total FAs, respectively[7]. As essential nutrients, LA and ALA cannot be endogenously synthesized in humans due to the lack of Δ15-desaturase and must be acquired through dietary sources[8]. As the downstream metabolites of ALA, eicosapentaenoic acid and docosahexaenoic acid exhibit a wide array of biological activities beneficial for human health[9]. These compounds show preventive effects against various disorders, including coronary heart disease[10], hypertension, diabetes, arthritis, high cholesterol, cancer, and autoimmune diseases. Consequently, the ALA content serves as a key quality indicator for edible oils. Developing high-ALA varieties represents a major objective in oil crop breeding.
Fatty acid desaturase 3 (FAD3) is a key enzyme in ALA biosynthesis and functions as the rate-limiting step[11]. FAD3 activity is regulated by its catalytic center structure, enzyme-substrate affinity, and protein conformational stability[12]. FAD3 proteins exhibit conserved structural features and high sequence homology among diverse species. Initially cloned from Arabidopsis thaliana, the FAD3 gene has been subsequently identified in various plants, including P. volubilis, Perilla frutescens, Glycine max, and Carthamus tinctorius[13−16]. Heterologous overexpression of the Jatropha curcas FAD3 (JcFAD3) gene significantly increases ALA accumulation in A. thaliana seeds[17]. The FAD3 expression markedly increases ALA content in the yeast mutant (H1246)[18]. Notably, elevated FAD3 expression in seeds strongly correlates with ALA accumulation, including in P. volubilis[19]. Transcriptome analysis reveals a direct relationship between ALA content and P. volubilis FAD3 (PvFAD3) expression[20]. Furthermore, heterologous PvFAD3 overexpression enhances ALA synthesis in Nicotiana benthamiana seeds[19]. Overall, FAD3 expression directly influences ALA accumulation in plant seeds, underscoring its crucial role in the regulation of ALA synthesis. However, current PvFAD3 research has mainly examined its expression effects on ALA levels and regulatory networks, whereas structural studies on its catalytic mechanism remain limited.
Certain conserved amino acid positions, at or near the catalytic center, can exert profound influences on enzyme stability, function, and activity[12]. For example, the E6V mutation in hemoglobin's β-subunit causes sickle cell anemia[21]. Site-directed mutagenesis of TesA thioesterase significantly enhanced octanoic acid (C8:0) production via the Type II fatty acid synthesis pathway[22]. Contemporary methods, including functional domain exchange, site-specific mutation, and bioinformatics, are employed to explore the intricate relationship between enzyme structure and function[23,24]. In plants, membrane-bound FAD3 exhibits greater structural complexity than soluble proteins[25]. Thus, this complexity impedes FAD3 purification, limiting applications of cryo-electron microscopy and nuclear magnetic resonance techniques. Currently, limited research on the crystal structure of Δ9 fatty acid desaturase (belonging to the same family as FAD3) in humans and mammals has indicated the presence of four hydrophobic transmembrane structures[26,27]. This finding serves as a valuable reference for studying the structure and function of FAD3. Plant FAD3 contains 3–4 transmembrane domains (TM1–TM4) and two amphipathic helices (AH1−AH2)[25]. Importantly, three conserved histidine clusters (HX3H in the His-boxI region, HX2HH in the His-boxII, and His-boxIII regions) coordinate non-heme Fe2+, essential for FAD3 activity[25]. Despite these insights, limited reports explore the relationship between the structure of the FAD3 enzyme and its catalytic function. Additional studies are needed to elucidate this mechanism.
In this investigation, a comprehensive approach involving homology modelling, model evaluation, conserved domain prediction, and homology sequence comparison analysis was employed to predict the pivotal sites influencing the activity of the PvFAD3 protein. Using site-directed mutagenesis via the bypass method, a mutant PvFAD3 (mPvFAD3) variant was generated, and key residues in PvFAD3 were substituted. To assess the impact of this substitution on enzyme activity, both PvFAD3 and mPvFAD3 were overexpressed in Saccharomyces cerevisiae, and replacement experiments were conducted in A. thaliana FAD3 mutants (AtFAD3m). Gas chromatography-mass spectrometry (GC-MS) revealed the influence of key sites on PvFAD3 activity. Identifying these critical PvFAD3 residues advances our understanding of fatty acid desaturase mechanisms and enables novel oil crop improvement strategies.
-
Mature PVS samples were sourced from the plantation base of Hainan University, located in Danzhou, Hainan, China (Fig. 1; longitude and latitude: 109.503179, 19.542727)[20]. Post-collection, fruits were meticulously peeled, and the seeds were extracted, promptly frozen in liquid nitrogen, and stored at −80 °C for future use.

Figure 1.
Phenotypes of Plukenetia volubilis and its fruits.
The T-DNA insertion line FAD3 silencing mutant A. thaliana (SALK_071996C) was procured from AraShare (
www.arashare.cn ). Subsequent to germination, the seeds were cultivated in a controlled environment within an artificial climate incubator, main-taining conditions of 23 °C, 60% humidity, light intensity ranging from 100−150 μmol/m2/s, and a light period of 16 h, followed by 8 h of darkness.Homology modelling and molecular docking
-
The PvFAD3 gene sequence was extracted from previous transcriptome data of P. volubilis[20]. The amino acid sequence of PvFAD3 was obtained using SnapGene software. The tertiary structure of PvFAD3 was predicted using the Phyre2 online prediction tool (
www.sbg.bio.ic.ac.uk/phyre2 ). To validate the reliability of the predicted model, the SAVES online server (https://saves.mbi.ucla.edu/ ) was employed. The three-dimensional data structure of the substrate LA was retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/ ). AutoDock Tools (ADT) was utilized for processing, resulting in the generation of a ligand structure file. The PvFAD3 simulated protein was dehydrated and hydrogenated using ADT software to maintain the original charge, designating it as the receptor structure. Further, ADT was employed to conduct rigid docking of small ligand molecules to the receptor. The grid box size were set to 30 Å × 30 Å × 30 Å, with a typical grid spacing of 0.375 Å. Ultimately, the tertiary structure of PvFAD3 was analyzed using SPDBV software.Homologous sequence comparison
-
To assess the homology of FAD3 proteins, a search was conducted using the NCBI database (
www.ncbi.nlm.nih.gov/ ). A total of 26 protein sequences homologous to PvFAD3 were selected (Supplementary Table S1). The conserved domain within the full-length amino acid sequence of FAD3 was predicted using the MEME online tool (http://meme-suite.org/ ). The full-length amino acid sequences of 27 FAD3, including PvFAD3, were compared and analyzed using DNAMAN software.Topological structure analysis of PvFAD3
-
Protein hydrophilicity and hydrophobicity of PvFAD3 was analyzed using DNAMAN software. The TMHMM online program (
www.cbs.dtu.dk/services/TMHMM/ ) was utilized to predict the PvFAD3 topology information across the membrane structure. Based on the aforementioned information, the topological model for PvFAD3 proteases was constructed.Objective gene acquisition and construction of an expression vector
-
Based on the gene sequences of PvFAD3, specific primers (Supplementary Table S2) were designed to obtain the full-length gene of PvFAD3. The gene sequence of mPvFAD3 was obtained using a 'bypass method' (Supplementary Fig. S1). By introducing the gene sequences of PvFAD3 and mPvFAD3 into the pYES2 vector treated with EcoRI and HindIII endonucleases, the recombinant vectors pYES2-PvFAD3 and pYES2-mPvFAD3 were generated (Supplementary Fig. S2). Similarly, by incorporating the gene sequences of PvFAD3 and mPvFAD3 into the pBI121 vector treated with XbaI and SacI endonucleases, the recombinant vectors pBI121-PvFAD3 and pBI121-mPvFAD3 were obtained. The resulting recombinant vector was sent to Hainan Nanshan Biotechnology Company (Hainan, China) for sequencing, and stored at −20 °C for future use.
Transgenic yeast culture
-
Yeast transformation was conducted employing thermal stimulation, with transgenic S. cerevisiae strains harboring the empty pYES2 vector as the negative control. Transgenic S. cerevisiae strains carrying the recombinant vectors pYES2-PvFAD3 and pYES2-mPvFAD3 underwent inoculation in SD-Ura liquid medium for activation culture. Under aseptic conditions, bacteria were collected through centrifugation and then inoculated into SG/-Ura liquid medium containing 1% raffinose at a ratio of 1:100. Subsequently, 50 μL of LA with a final concentration of 300 μg/mL was added, and 1% surfactant IGE-PAL CA-630 was included in the medium to emulsify LA and facilitate its cellular entry. The yeast was cultured at 28 °C and 180 rpm in a shaker, and at 5 °C and 180 rpm in a separate shaker for 36 to 72 h until reaching a stable stage. Yeast cells were then collected by centrifugation at 5,000 rpm, washed three times with deionized water, dried using a vacuum freeze dryer, and stored at −80 °C for future use. Three parallel samples were prepared for the experiment.
Arabidopsis mutant replacement
-
The AtFAD3m seeds were sown and nurtured for robust growth. The three-primer method was used to identify mutant homozygotes (Supplementary Table S2). Homozygous AtFAD3m seeds were subsequently cultivated, and the PvFAD3 and mPvFAD3 genes were integrated into the Arabidopsis genome using Agrobacterium-mediated transformation. Transgenic positive seedlings were chosen on a 1/2MS plate supplemented with 50 μg/mL Kanamycin (Kana+), and DNA extracted from these seedlings underwent PCR verification using specified primers (Supplementary Table S2). After screening until the T3 generation, 20 mg of seeds were collected and weighed for subsequent FA extraction and detection.
RT-qPCR validation
-
Total RNA was isolated using the RNeasy Plant Mini Kit (Qiagen, Germany), followed by cDNA synthesis with the GoScript Reverse Transcription System (Promega, USA). AtACT2 (AT3G18780) served as the endogenous control, as previously established[28]. RT-qPCR was performed using ChamQ Universal SYBR qPCR Master Mix (Vazyme, China), on a LightCycler 96 system (Roche, Switzerland). Primer sequences are provided in Supplementary Table S3. Three technical replicates were analyzed, with relative expression levels calculated using the 2−ΔΔCᴛ method.
FA extraction and detection
-
Heptadecanoic methyl ester (ANPEL, Shanghai, China), and GLC Mixture (ZZSRM, Shanghai, China) were used as the internal and external standards, respectively. An Agilent 6,890 gas chromatograph with a flame ion chromatograph detector was used for analysis. Column box operation procedure: 50 °C for 1 min, 10 °C/min rising to 150 °C for 1 min, and then 4 °C/min rising to 250 °C for 3 min. The sample was loaded with an automatic injector with a loading volume of 1 µL and a split ratio of 50:1. Helium was used as a carrier gas with a flow rate of 1 mL/min.
Using GC-MS data analysis software, fatty acid methyl ester (FAME) standard samples (ANPEL, Shanghai, China) were employed as references to compare the retention times and peak areas of a mixed FAME solution and heptadecane methyl. The percentage composition of fatty acid components was then calculated using the area normalization method. The FA methyl ester samples of transgenic yeast and A. thaliana seeds were analyzed qualitatively and quantitatively. Statistical software GraphPad Prism version 8.0.2.263 was used to analyze the significance of differences. The statistical method employed was one-way ANOVA, and a p-value < 0.05 indicated statistically significant differences among the groups.
-
The tertiary structure of PvFAD3 was generated using the Phyre2 online server. The prediction showed 99.8% confidence for template c4zyoA as the PvFAD3 structural model (Fig. 2a). Model quality was assessed through the SAVES online server. The pull diagram results indicated that 97% of the amino acid residues in the c4zyoA model conformed to stereochemical rules within the allowed region (Fig. 2b), confirming its suitability for subsequent analyses. The three-dimensional structure of the substrate LA was obtained from the PubChem database (Fig. 2c). Molecular docking was performed using ADT software after structure preparation. Rigid docking of LA with the PvFAD3 model simulated their binding interaction (Fig. 2d). Docking results suggested mutual induction, deformation, and adaptation between the LA ligand and PvFAD3 mimic protein (Fig. 2d–f). Interestingly, the LA ligand formed a hydrogen bond with the aspartate at position 104 (ASP-104) of the PvFAD3 mimic protein (Fig. 2e, f). Furthermore, hydrogen bond analysis between amino acid residues indicated a bond between ASP-104 and asparagine at position 109 (ASN-109) of the PvFAD3 mimic protein (Fig. 2g). These findings imply that ASP-104 may play a crucial role in protein stability, substrate binding, and potentially have a significant impact on enzyme function.
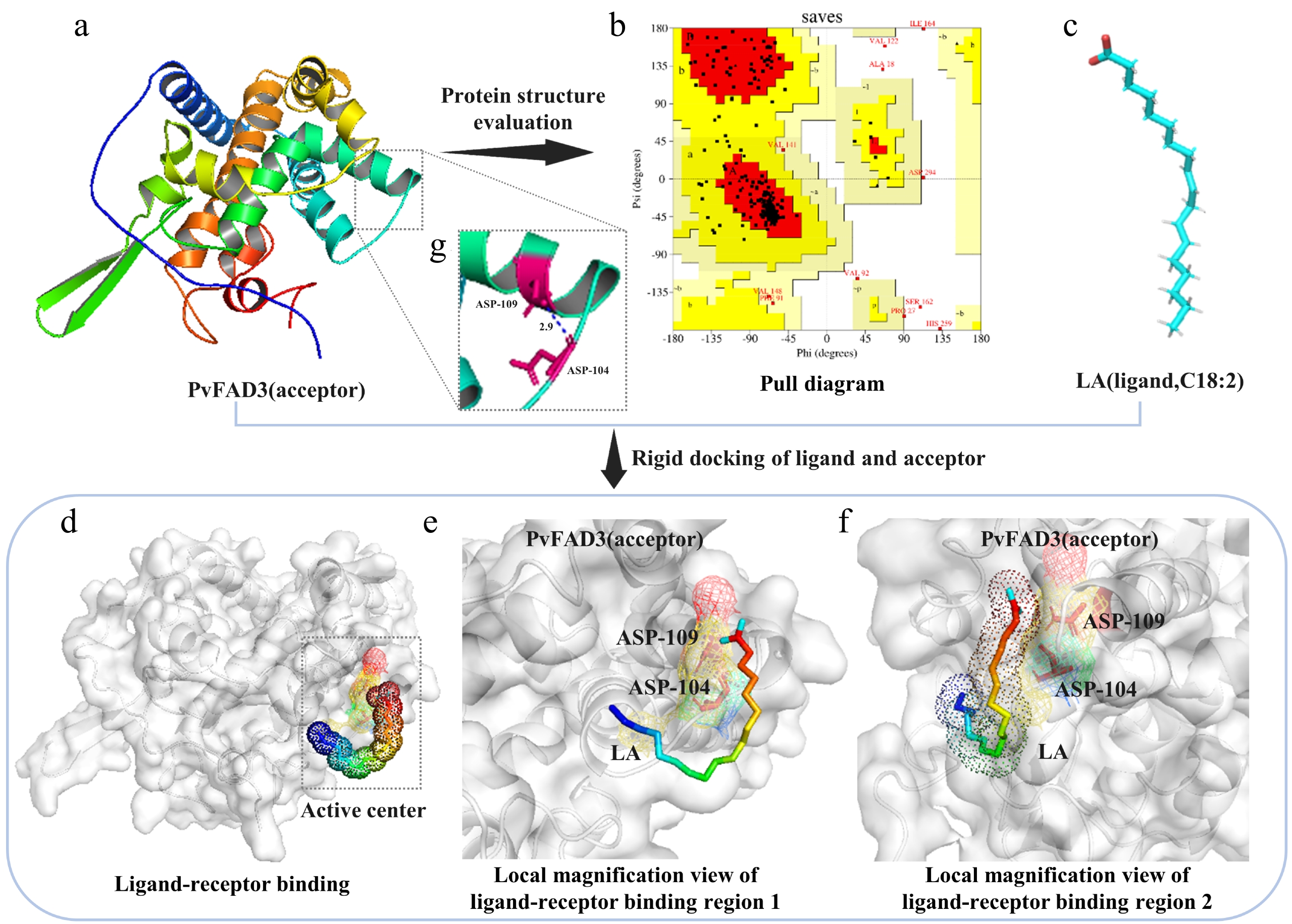
Figure 2.
Model prediction and molecular docking. (a) PvFAD3 tertiary structure prediction. (b) The reliability of the predicted model was evaluated. The Ramachandran plot results showed that 84.1% of the amino acids in the PvFAD3 protein model were located in the most favored region (red region), 11.6% in other allowed regions (yellow region), 1.3% in generally allowed regions (light brown region), and 3% in disallowed regions (white region). If the proportion of amino acid residues falling within the allowed region of the protein model is higher than 90%, the conformation of the model can be considered to conform to the principles of stereochemistry. (c) The tertiary structure of ligand LA. (d) Substrate protein docking prediction. The dashed rectangular region represents the rigid docking active site. (e) Top view of ligand LA small molecule binding to PvFAD3 mimicking protein. (f) Side-view magnification of ligand LA small molecule binding to PvFAD3 mimicking protein. The LA ligand molecule forms hydrogen bonds with the side chain R group of ASP-104 in the simulated PvFAD3 protein. (g) The intramolecular structure of the PvFAD3 protein model was analyzed. The amino acid residues at positions 104 (ASP-104) and 109 (ASN-109) were connected by hydrogen bonds.
Prediction analysis of conserved domains
-
To further evaluate the impact of ASP-104 on FAD3 function, 27 FAD3 homologs were analyzed across diverse species. Three conserved motifs were identified: motif 1 (positioned between 18–67), motif 2 (positioned between 96–145), and motif 3 (positioned between 262–310) (Supplementary Fig. S3). Notably, Asp104 and Asn109 localize within conserved motif 2, comprising the critical region for PvFAD3 protease activity (Fig. 3a). Multiple sequence alignment highlighted the conservation of ASN-109 across all species, while ASP-104 exhibited relative conservation (Fig. 3b). Intriguingly, these species with higher ALA content (such as P. volubilis, Perilla frutescens, Paeonia suffruticosa, etc.) possess aspartate at position 104, whereas these species with lower ALA content (such as Arachis hypogaea, Juglans regia, Prunus armeniaca, etc.) have asparagine or glutamate at position 104 (Fig. 3b). This correlation establishes Asp-104 as a determinant of FAD3 catalytic efficiency.
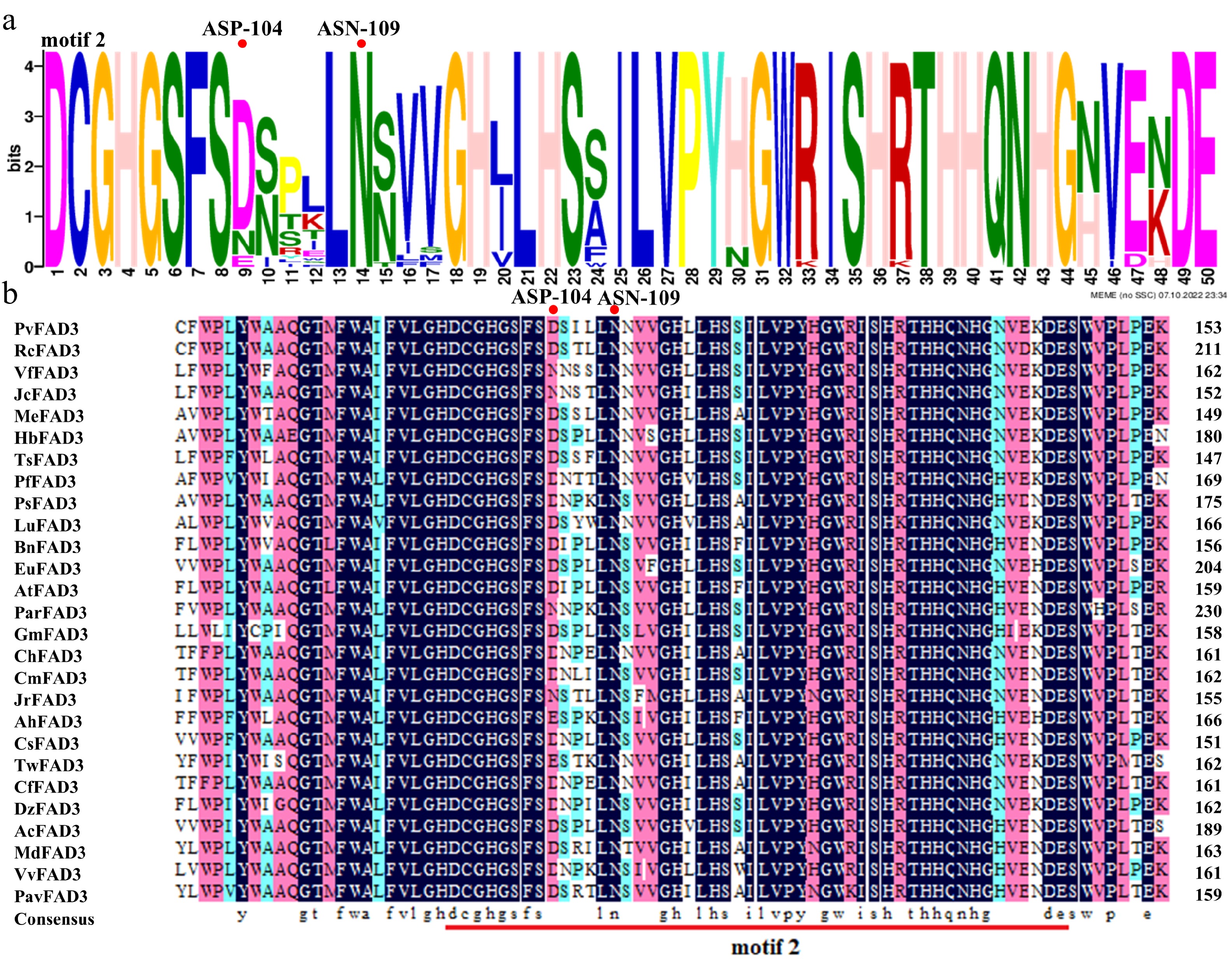
Figure 3.
Conserved domains are predicted by homologous sequence alignment. (a) ASP-104 and ASN-109 are located in the PvFAD3 protein conserved domain motif 2. The larger the letter font, the more conserved the site. (b) The comparison of FAD3 proteins in 27 different species. There were significant differences in ASP-104 among different species, while ASN-109 was absolutely conserved. Dark blue: amino acid homology = 100%, Pink: amino acid homology ≥ 75%, Light blue: amino acid homology ≥ 50%. RcFAD3, EEF36775.1; VfFAD3, AAC98967.2; JcFAD3, NP_001292929.1; MeFAD3, XP_021610352.1; MeFAD3, XP_021610352.1; HbFAD3, XP_021666224.1; TsFAD3, ABM68629.1; PfFAD3, AQZ42316.1; PsFAD3, AVZ47050.1; LuFAD3, BAG70950.1; BnFAD3, AAT09135.1; EuFAD3, ARQ20744.1; AtFAD3, NP_180559.1; ParFAD3, CAB4284757.1; GmFAD3, NP_001236114.1; ChFAD3, AEF80000.1; CmFAD3, KAF3952997.1; JrFAD3, XP_018835457.1; AhFAD3, QBI71562.1; CsFAD3, XP_028086292.1; TwFAD3, XP_038705526.1; CfFAD3, KAE8039281.1; DzFAD3, XP_022756628.1; AcFAD3, PSS09495.1; MdFAD3, XP_008384790.2; VvFAD3, XP_002277573.1; PavFAD3, XP_021816263.1.
Topological structure analysis of PvFAD3
-
To construct a topological model of PvFAD3 protein, the analysis of hydrophilic regions, hydrophobic regions, and transmembrane structure was performed. Combined hydrophilicity/hydrophobicity (Supplementary Fig. S4), and transmembrane domain (Fig. 4a) predictions yielded the PvFAD3 topological model (Fig. 4b). The PvFAD3 protein displayed three non-signal peptide transmembrane structures (TM1, TM2, and TM3), along with two inlay membrane structures (AH1 and AH2), and three conserved histidine regions (1 HX3H and 2 HX2HH, known as His-boxI, II, and III) (Fig. 4b). The catalytic His-box motifs localize to the cytoplasmic side of the endoplasmic reticulum membrane (Fig. 4b). The ASP-104 site of the PvFAD3 protein was identified within the AH1 membrane structure, situated between the active regions of His-boxI and His-boxII.
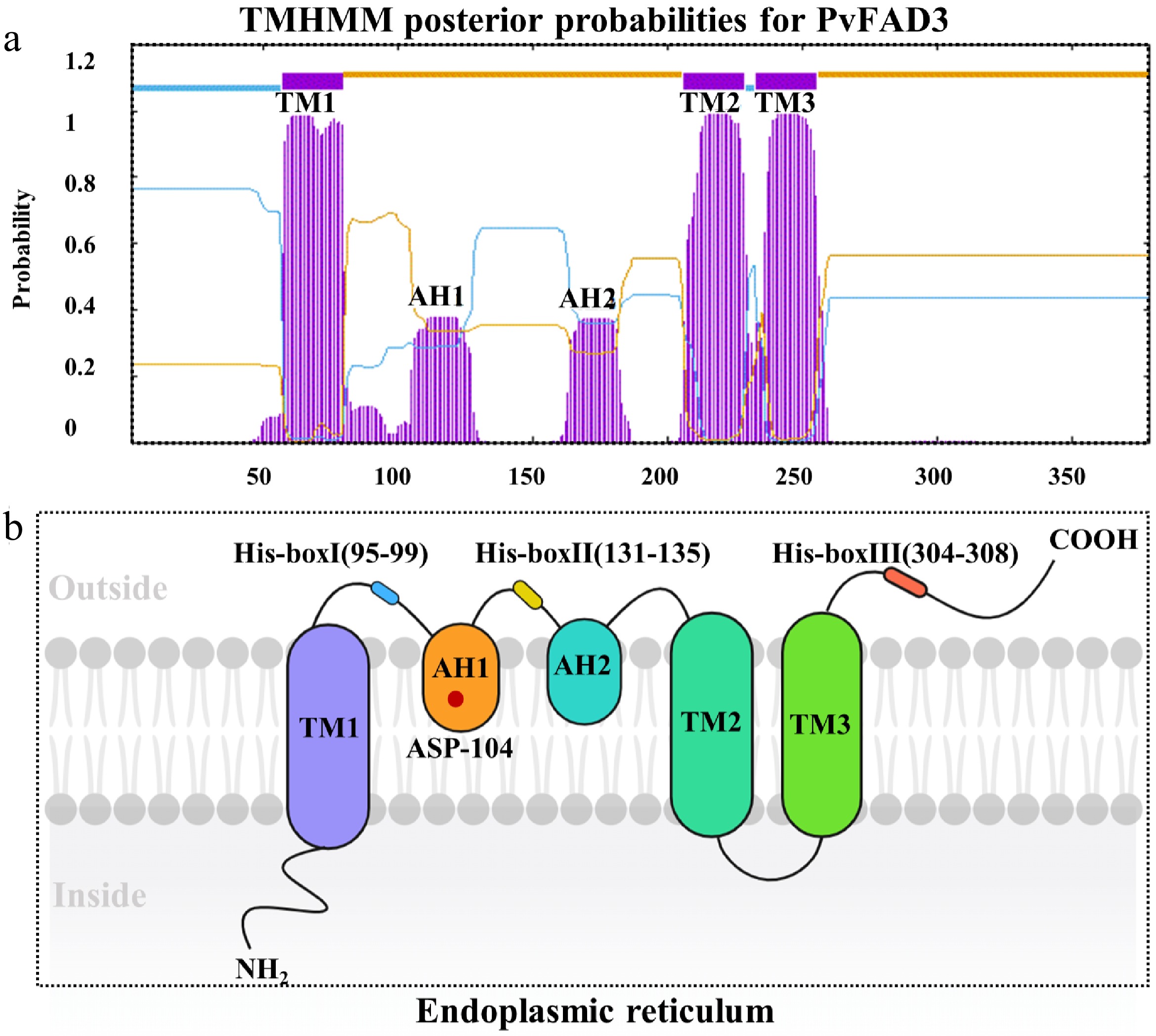
Figure 4.
The topological model of PvFAD3. (a) PvFAD3 protein cross membrane structure prediction. The purple structure represents a cross-membrane or inlaid protein structure. (b) Topological model prediction of PvFAD3 proteins. TM1, TM2, and TM3: Protein transmembrane structure. AH1 and AH2: Protein mosaic structure. His-boxI, His-boxII, and His-boxIII: Histidine active region. Red Star: ASP-104.
In vitro verification of ASP-104 for PvFAD3 activity
-
To investigate the impact of ASP-104 on PvFAD3 activity, aspartate was substituted with asparagine at position 104. PvFAD3 and mPvFAD3 were both cloned into a pYES2 vector and transformed into S. cerevisiae (Supplementary Figs S5–S10). In the control group, the main FA components were monounsaturated FAs (Table 1). Compared with PvFAD3 transgenic yeasts, mPvFAD3-transformed yeasts had a similar FA profile with the control group (Table 1). For example, there was a slight difference in C14:0, C16:0, C16:1, C18:0, and C18:1 contents between mPvFAD3-transformed yeasts and the control group (Table 1). Interestingly, the expression of PvFAD3 resulted in a significant several-fold accumulation of saturated FAs, such as C14:0 and C16:0 (Table 1).
Table 1. Fatty acid compositions in different yeasts.
Culture condition Group FA components (%) C14:0 C16:0 C16:1 C18:0 C18:1 C18:2 C18:3 30 °C pYES2 1.28 ± 0.34c 12.16 ± 0.76e 43.12 ± 0.61a 2.82 ± 0.06c 21.36 ± 0.65a 19.26 ± 0.40c 0.00 ± 0.00e pYES2-PvFAD3 12.63 ± 0.45a 44.03 ± 0.54b 6.24 ± 0.08c 1.59 ± 0.04d 9.87 ± 0.55c 17.14 ± 0.64d 8.49 ± 0.09b pYES2-mPvFAD3 1.67 ± 0.07c 15.27 ± 0.53d 42.23 ± 0.49a 4.15 ± 0.06b 22.77 ± 0.55a 7.35 ± 0.39e 6.56 ± 0.17c 20 °C pYES2 0.00 ± 0.00d 15.98 ± 0.30d 29.01 ± 0.42b 4.53 ± 0.22a 14.09 ± 0.76b 36.39 ± 0.71a 0.00 ± 0.00e pYES2-PvFAD3 10.99 ± 0.23b 62.04 ± 0.34a 1.42 ± 0.15d 1.33 ± 0.07d 6.78 ± 0.43d 1.92 ± 0.28f 15.52 ± 0.32a pYES2-mPvFAD3 1.94 ± 0.22c 18.50 ± 0.54c 28.09 ± 0.43b 4.66 ± 0.05a 19.64 ± 0.68a 22.10 ± 0.44b 5.06 ± 0.47d Different letters indicate significant differences (p < 0.05). The primary focus lies in the alterations in ALA content. Under cultivation at 30 °C, the ALA (C18:3) content in PvFAD3- and mPvFAD3-transformed yeasts increased by 8.49% and 6.56%, respectively, compared with the control group (Table 1). Under 20 °C cultivation conditions, PvFAD3- and mPvFAD3-transformed yeasts exhibited increases of 15.52% and 5.06%, respectively (Table 1). Although both PvFAD3 and mPvFAD3 expressions increased ALA production, wild-type PvFAD3 showed significantly greater enhancement (Table 1). Notably, when cultivated at 20 °C, the ALA content in PvFAD3-transformed yeasts was three times higher than that in mPvFAD3-transformed yeasts (Table 1). These findings illustrate the ability for PvFAD3 to effectively enhance ALA accumulation at 20 °C.
In vivo verification of ASP-104 for PvFAD3 activity
-
Homozygous Atfad3 mutants were genotyped using a three-primer PCR strategy (Fig. 5a). PvFAD3 and mPvFAD3 were cloned into the pBI121 vector and transformed into homozygous AtFAD3m. Through resistance selection and PCR analysis, complementation lines for PvFAD3 and mPvFAD3 were screened (Fig. 5b–e). The RT-qPCR demonstrated that there is no significant difference in the expression levels of PvFAD3 and mPvFAD3 in their respective corresponding A. thaliana strains (Fig. 5f). It was obvious that PvFAD3 and mPvFAD3 complementation lines exhibit more robust growth phenotypes compared to AtFAD3m (Fig. 5f, g). Compared with AtFAD3m seeds (16.18%), total oil contents were significantly increased in PvFAD3 (22.60%) and mPvFAD3 (20.53%) complementation lines (Table 2). The FA composition analysis showed that PvFAD3 complementation seeds had a significant increase in the average content of C18:2, C18:3, C20:0, and C20:1, with improvements of 39.80%, 63.27%, 38.89%, and 42.90%, respectively (Table 2). The mPvFAD3 complementation seeds exhibited a significant increase in the average content of C18:2 and C20:0, with improvements of 25.71% and 20.24%, respectively (Table 2). Importantly, when compared with AtFAD3m lines, the PvFAD3 complementation lines demonstrated a significant increase in C18:3 content, while mPvFAD3 complementation lines displayed no significant change (Table 2).
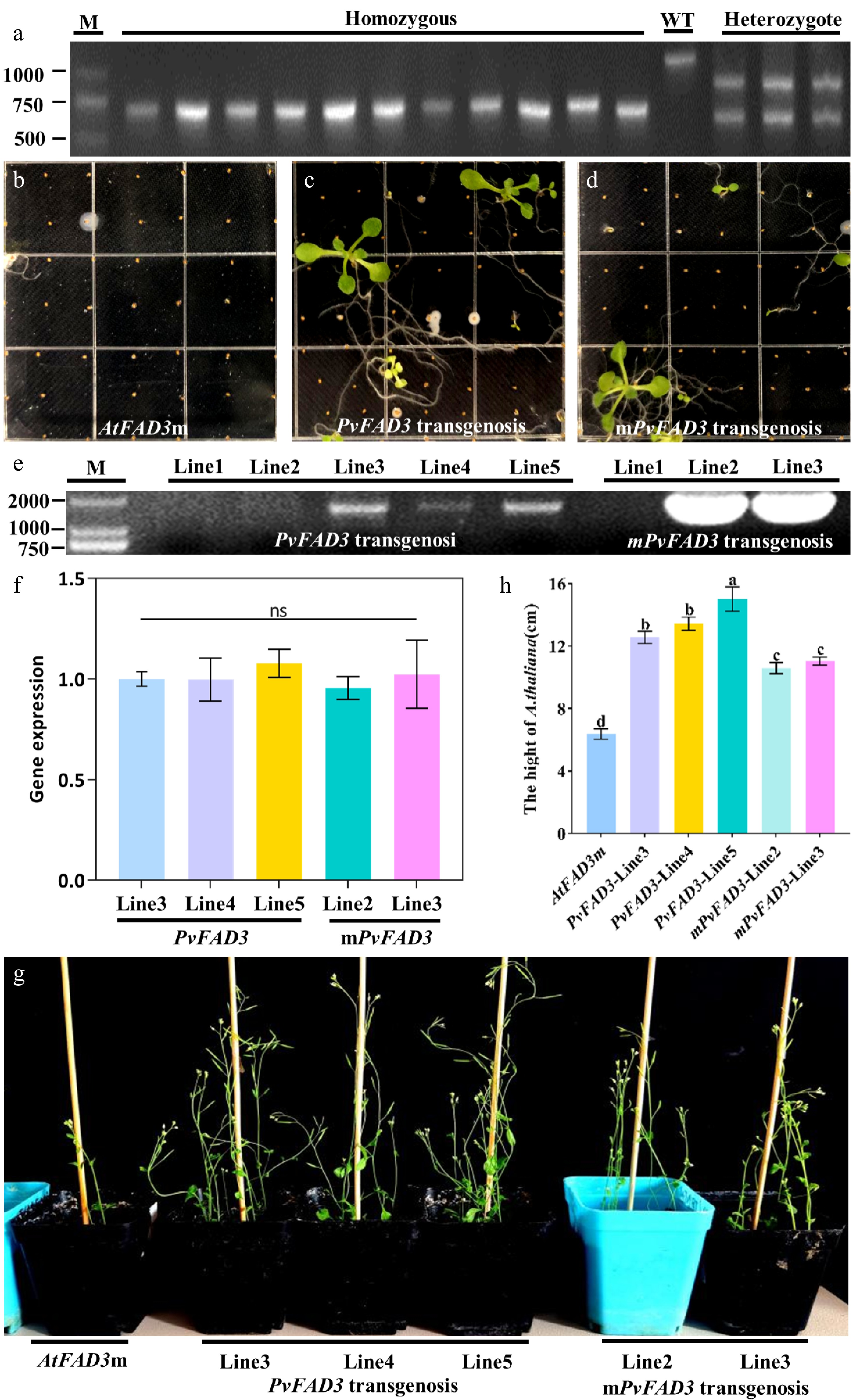
Figure 5.
Homozygous screening and genetic transformation of AtFAD3m. (a) AtFAD3m homozygote, WT and AtFAD3m heterozygote. (b) AtFAD3m seeding. (c) The transgenic PvFAD3 positive seedlings of T1 generation were screened for Kana+ resistance. (d) The transgenic mPvFAD3 positive seedlings of T1 generation were screened for Kana+ resistance. (e) PCR screening of transgenic plants. (f) RT-qPCR validation of transgenic plants. (g) When T3 positive seedlings were transplanted into soil for 5 weeks, the growth state of transformed PvFAD3 and mPvFAD3 was significantly better than that of AtFAD3m. (h) Plant height statistics of AtFAD3m, PvFAD3, and mPvFAD3.
Table 2. Fatty acid compositions in Arabidopsis seeds.
Group FA components (%) C16:0 C18:0 C18:1 C18:2 C18:3 C20:0 C20:1 Total AtFAD3m AtFAD3m - line1 1.62 ± 0.02d 0.78 ± 0.04c 2.79 ± 0.06de 6.94 ± 0.06g 0.21 ± 0.02b 0.42 ± 0.02d 3.03 ± 0.05f 15.78 ± 0.08f AtFAD3m - line2 1.65 ± 0.06d 0.80 ± 0.04c 2.93 ± 0.06d 7.51 ± 0.08e 0.20 ± 0.03b 0.44 ± 0.03cd 3.15 ± 0.04e 16.68 ± 0.16e AtFAD3m - line3 1.64 ± 0.03d 0.80 ± 0.06c 2.74 ± 0.04e 7.25 ± 0.15f 0.19 ± 0.01b 0.40 ± 0.02d 3.07 ± 0.05ef 16.09 ± 0.26f PvFAD3 transfer PvFAD3 - line3 1.77 ± 0.06c 0.79 ± 0.04c 3.59 ± 0.19c 9.78 ± 0.03b 0.29 ± 0.02a 0.55 ± 0.04b 4.13 ± 0.07c 20.90 ± 0.33c PvFAD3 - line4 1.75 ± 0.02c 0.83 ± 0.06c 3.62 ± 0.06c 9.53 ± 0.06c 0.30 ± 0.02a 0.57 ± 0.02b 4.12 ± 0.05c 21.73 ± 0.12c PvFAD3 - line5 2.35 ± 0.06a 1.10 ± 0.03a 4.67 ± 0.13a 10.96 ± 0.05a 0.29 ± 0.03a 0.65 ± 0.02a 5.14 ± 0.05a 25.17 ± 0.06a mPvFAD3 transfer mPvFAD3 - line2 1.76 ± 0.08c 0.85 ± 0.03bc 3.68 ± 0.09c 8.49 ± 0.08d 0.19 ± 0.01b 0.47 ± 0.02c 3.94 ± 0.08d 19.40 ± 0.16d mPvFAD3 - line3 1.92 ± 0.07b 0.91 ± 0.02b 4.08 ± 0.05b 9.67 ± 0.10bc 0.19 ± 0.02b 0.53 ± 0.03b 4.36 ± 0.06b 21.66 ± 0.04b Different letters indicate significant differences (p < 0.05). -
As an essential ω-3 fatty acid obtained from plant oils, ALA plays vital physiological roles in humans. ALA not only contributes to lowering hypertension and hypertriglyceridemia, but also undergoes conversion into eicosapentaenoic acid and 3-series prostaglandins within the body[29,30]. The consumption of vegetable oils rich in ALA has been linked to significant cardiovascular benefits[29]. ALA-enriched oils are increasingly incorporated into human diets globally. Recognized as a perennial woody oil plant, P. volubilis yields a high content of polyunsaturated FAs, particularly rich in ALA[31]. Although the ALA content of PVS oil is slightly lower than that of linseed oil, it significantly surpasses that of commonly used edible oils, including peanut oil, maize oil, rapeseed oil, and olive oil[7]. Remarkably, P. volubilis, being a perennial plant, allows for multiple harvests with a single planting and does not require arable land. Consequently, P. volubilis has emerged as a novel and economically valuable oil crop in southern China. Research has consistently demonstrated that FAD3 serves as the principal rate-limiting enzyme in ALA biosynthesis[11]. Previous investigations into omega-3 FAD functionality have underscored the pivotal role of enzyme affinity and stability in determining enzymatic activity[15]. The affinity of enzyme molecules mainly refers to their ability to bind to substrates, while the stability of enzyme molecules is closely related to the spatial structure of their active regions[32]. PvFAD3, being a transmembrane protein[19], has posed challenges in in vitro purification, hindering the elucidation of its three-dimensional spatial structure through conventional crystallization methods. Predicting the protein's three-dimensional structure has also proven elusive through traditional means. In recent years, as protein and enzyme sequence data have burgeoned, the focus of functional analysis has shifted from conventional molecular biology experiments toward a hybrid approach that integrates experimental work with computer technologies. Such integration provides unprecedented insights into enzymatic mechanisms[24]. In this study, various software tools were employed to characterize the structural features, key binding sites with substrates, and hydrophobic transmembrane structures. The prediction results notably highlight the significant impact of ASP-104 on the affinity and stability of the PvFAD3 protein.
The genetic background and metabolic pathways of S. cerevisiae are well understood, and it is known for its rapid growth, easy mass production, and GRAS certification[33]. Previous studies have successfully achieved heterologous expressions of various recombinant proteins in S. cerevisiae, including Δ6 and Δ12 FADs[34,35]. Utilizing free linoleic acid as a substrate, the heterologous expression of linoleic acid isomerase has been shown to yield multiple conjugated linoleic acids[36]. Hence, exploring the catalytic activity differences between PvFAD3 and mPvFAD3 through heterologous expression in S. cerevisiae is a rational and effective approach. Through transgenic S. cerevisiae and GC-MS analysis, both PvFAD3 and mPvFAD3 expressions increased ALA contents, yet the former exhibited a significantly more substantial increase (Table 1). This result strongly supports the notion that ASP-104 positively influences PvFAD3 activity. This conclusion is further substantiated in transgenic A. thaliana. While both PvFAD3 and mPvFAD3 over-expressions in AtFAD3m enhanced ALA content in mature seeds, the impact of mPvFAD3 expression was not as pronounced as that of PvFAD3 expression (Table 2). Considering the outcomes of homologous modeling and molecular docking (Fig. 2), there is compelling evidence to speculate that ASP-104, as a key site in the PvFAD3 protein, positively influences the enzyme's activity.
Previous reports have proposed that the FAD3 protein is a low-temperature-active enzyme, suggesting that a lower culture temperature can enhance polyunsaturated FA content[37]. Indeed, the ALA content of PvFAD3 transgenic yeast nearly doubled when the cultivation temperature was reduced from 30 °C to 20 °C (Table 1). However, the ALA content of mPvFAD3 transgenic yeast did not exhibit similar responsiveness to the lower culture temperature. Conversely, a decrease in culture temperature for mPvFAD3 transgenic yeast from 30 °C to 20 °C resulted in a 22.87% decline in ALA content (Table 1). Under the same temperature conditions, the ALA content in the PvFAD3 was significantly higher than that in the mPvFAD3, and reducing the temperature further enhanced the statistical significance of this difference (Table 1). The radio tracer experiments have indicated that desaturase activity in Brassica napus comprises both temperature-independent basal desaturase activity, and low-temperature-induced desaturase activity[38]. It can therefore be assumed that ASP-104 may be involved in low-temperature-induced desaturase activity of PvFAD3 protein. To date, the molecular mechanisms underlying temperature modulation of FA profiles remain elusive.
Following FA biosynthesis, the saturated C16:0-ACP can either be hydrolyzed for C16:1 generation or be further elongated into C18:0-ACP, subsequently desaturated into C18:1, C18:2, and C18:3[20]. The intermediate metabolite, C16:0, generated during this process, exclusively chooses one metabolic pathway. Consistent with expectations, the PvFAD3 transgenic yeast exhibited a significant reduction in C16:1 content (Table 1), suggesting that the PvFAD3 expression could inhibit the desaturation of C16:0 to form C16:1. The inhibitory effect on C16:1 accumulation was more significant at lower temperatures. Because Arabidopsis seeds have very low C16:1 content[39], this inhibitory phenomenon has not been observed in PvFAD3 transgenic plants. In addition, it is worth noting the extreme accumulation of C16:0 in PvFAD3 transgenic yeast (Table 1). This can be explained considering that PvFAD3 inhibits the carbon flux towards C16:1. Interestingly, mPvFAD3 expression did not significantly impact C16:0 and C16:1 content in yeast (Table 1). Whether the ASP-104 site of PvFAD3 protein can be related to the inhibitory role in C16:1 biosynthesis is still unclear and requires further verification. While it might be expected that 18-carbon chain FAs would notably increase in PvFAD3 transgenic yeast, this did not occur, likely due to the limited capacity for C18:0 biosynthesis in yeast[40]. For example, in transgenic PvFAD3 Arabidopsis, despite the absence of significant C16:0 accumulation, there was a general increase in the contents of C18:1, C18:2, C18:3, C20:0, and C20:1.
-
Through the combination of bioinformatics analysis and computer simulation techniques, the ASP-104 of PvFAD3 protein was identified as a key site involved in protein stability and substrate binding. Substituting asparagine for aspartate at position 104 in yeast expression and Arabidopsis mutant complementation experiments unequivocally demonstrated a substantial reduction in enzymatic activity for mPvFAD3, confirming the significance of ASP-104 as a key active site. This investigation enhances our comprehension of FAD3 enzyme activity, offering novel insights for the improvement of oil crop breeding and genetic engineering practices.
This work was supported by the National Natural Science Foundation of China (32401624). We thank BioRender (
https://app.biorender.com/ ) as the schematic diagram in the graphical abstract was created using BioRender.com.-
The authors confirm their contributions to the paper as follows: conceptualization, writing-original draft, methodology, validation, and formal analysis: Fu Y; methodology, validation, and formal analysis: Wang Z; methodology, investigation: Ye L; validation: Deng G; formal analysis: Li K, Zou Y; writing-review and editing: Wang J; conceptualization, writing-review and editing, project administration, funding acquisition: Niu J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0028)
-
Received 26 May 2025; Accepted 24 August 2025; Published online 20 October 2025
-
The key active site residue of the PvFAD3 protein was identified as ASP-104 by molecular docking methods.
Significant reduction of ALA content was found in transgenic mPvFAD3 yeast compared to PvFAD3.
Significant reduction of ALA content was found in transgenic mPvFAD3 Arabidopsis thaliana seeds compared to PvFAD3.
-
# Authors contributed equally: Yijun Fu, Zhaohui Wang
- Supplementary Table S1 FAD3 amino acid sequence information.
- Supplementary Table S2 Primers for PCR, vector construction and genetic transformation.
- Supplementary Table S3 Primers for qRT-PCR.
- Supplementary Fig. S1 Bypass method generates PvFAD3 mutants.
- Supplementary Fig. S2 PCR product gel electrophoresis experiment; 1, 2 pYES2-PvFAD3 and pYES2-mPvFAD3 colony PCR; 3, 4 pBI121-PvFAD3 and pBI121-mPvFAD3 liquid PCR.
- Supplementary Fig. S3 Conservative domain analysis of 27 FAD3 proteins.
- Supplementary Fig. S4 The analysis of the hydrophilic and hydrophobic regions.
- Supplementary Fig. S5 The fatty acid GC of S. cerevisiae with empty vector pYES2 was transformed by LA culture at 30 °C.
- Supplementary Fig. S6 LA was added to transform recombinant vector pYES2-PvFAD3 for fatty acid GC of S. cerevisiae at 30 °C.
- Supplementary Fig. S7 The fatty acid GC of S. cerevisiae with recombinant vector pYES2-mPvFAD3 was transformed by LA culture at 30°C.
- Supplementary Fig. S8 The fatty acid GC of S. cerevisiae with empty vector pYES2 was transformed by LA culture at 20 °C.
- Supplementary Fig. S9 LA was added to transform recombinant vector pYES2-PvFAD3 for fatty acid GC of S. cerevisiae at 20 °C.
- Supplementary Fig. S10 The fatty acid GC of S. cerevisiae with recombinant vector pYES2-mPvFAD3 was transformed by LA culture at 20 °C.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fu Y, Wang Z, Ye L, Deng G, Li K, et al. 2025. Predominance of the ASP-104 site of fatty acid desaturase 3 promoted a linolenic acid (ALA) accumulation in Plukenetia volubilis. Tropical Plants 4: e034 doi: 10.48130/tp-0025-0028 

Predominance of the ASP-104 site of fatty acid desaturase 3 promoted a linolenic acid (ALA) accumulation in Plukenetia volubilis
- Received: 26 May 2025
- Revised: 28 July 2025
- Accepted: 24 August 2025
- Published online: 20 October 2025
Abstract: Plukenetia volubilis is a perennial oilseed plant with high α-linolenic acid (ALA) content. Although fatty acid desaturase 3 (FAD3) is essential for ALA biosynthesis, its structural determinants and functional mechanisms remain unclear. This study employed molecular docking and bioinformatics to identify the aspartate at position 104 (ASP-104), as a pivotal residue in P. volubilis FAD3 (PvFAD3). Through in vitro site-directed mutagenesis, ASP-104 was converted to asparagine, generating the mutant PvFAD3 (mPvFAD3). Compared to yeast expressing PvFAD3, the mPvFAD3 expression showed 32.60% and 77.27% reductions in ALA content at 30 °C and 20 °C, respectively. Complementation assays revealed that heterologous expression of mPvFAD3 resulted in a 65.52% decrease in seed ALA content when compared to Arabidopsis thaliana seeds transformed with PvFAD3. These results conclusively demonstrate that ASP-104 is a critical site positively regulating PvFAD3 activity, with the mutation leading to a significant decline in ALA synthesis. The present work provides mechanistic insights into FAD3 function, enabling targeted genetic strategies for enhancing ALA production in oil crops.
-
Key words:
- Plukenetia volubilis /
- α-linolenic acid /
- PvFAD3 /
- Molecular docking /
- Mutation analysis