









-
Mulberry (Morus alba L.) is a widely cultivated plant species across Asia, Europe, North and South America, and Africa, for its economic significance and ecological restoration capabilities. Globally, Japan, India, and China predominantly grow a total of 30 mulberry species and nine varieties of mulberry[1]. In China alone, there are approximately 3,000 preserved germplasm resources of mulberry, comprising 15 species and four varieties[2]. The primary use of mulberry in China has been for silkworm rearing (Bombyx mori L.), contributing to local farming and textile industries. Moreover, mulberry products such as foods, drinks, and medicines have gained popularity due to their perceived health benefits including the reduction of risks associated with high blood glucose, coronary heart disease, and atherosclerosis[3−5]. Furthermore, mulberry's fast growth rate and adaptability under changing global environmental conditions make it a promising candidate for phytoremediation and bio-energy production[6,7]. However, mulberry planting areas often face challenges such as soil barrenness, drought, salinity waterlogging, and heavy metal ions exposure, which limit the healthy growth and high yield potential of mulberries.
Arbuscular mycorrhizal fungi (AMF) are crucial and ubiquitous components of soil microbial communities, forming mutually beneficial symbiotic associations with the roots of over 80% of terrestrial plants[8,9], including cash crops like mulberry[10], coffee[11], and tea[12]. This association significantly enhances crop productivity, nutrient cycling, and ecosystem sustainability. AM fungi are well-known for providing host plants with soil phosphorus (P) and nitrogen (N), as well as other benefits such as protection against pathogens, in exchange for plant-assimilated carbon[8,13]. Moreover, AMF improves soil structure through the production of the glomalin glycoprotein and the formation of an extensive mycelium network in the soil, which directly promotes the absorption of soil water and nutrients from the rhizosphere[8,14]. Glomalin related soil proteins (GRSP) have been reported as a stable and persistent protein produced in substantial quantities by mycorrhizal fungi, potentially constituting a significant reservoir of organic N in soil[15]. Studies have shown that AMF promotes root N accumulation and uptake through GRSP-regulated soil nitrogen[16]. Inoculation with AMF improves mulberry's resistance against drought, salinity, alkalinity, diseases, and heavy metal toxicity by increasing plant nutrient uptake from soil and enhancing photosynthesis capacity[17−21]. Also, AMF inoculation has been found to improve the survival rate of mulberry seedlings while reducing fertilizer application requirements, ultimately leading to higher yields and improved quality of mulberry leaves[22−24]. Different AMF species within the same genus or different genera play unique roles in mulberry growth and leaf quality[24]. Given the long history of coevolution between AMF and plants in various ecosystems, resulting in their adaptation to specific areas, identifying AMF species and exploring their community composition is crucial for evaluating their beneficial functional potentials to the host plant.
Over the past two decades, extensive research has been conducted on the association between crop plants and AMF across various agricultural conditions. Various agricultural management practices such as fertilization[25], land-use changes[26], tillage[27], host crop[28], geography[29,30], cropping systems[31,32], and soil type[33], have been found to impact the abundance or communities of AMF in soil. For instance, AMF colonization in rice roots occurs only under dry conditions, rather than in the conventional wet paddy fields[34]. Differences in soil available P, soil organic carbon, fertilization, and soil layer depth influence AMF communities in maize and wheat[35]. Specifically, increased concentrations of soil P fertilization or inorganic N application have been shown to reduce AMF root colonization, as well as AMF species richness and diversity[16,36]. AMF spore densities vary significantly with soil depth, being lower at 50–70 cm than at 20–50 cm and at 20–50 cm than at 0–20 cm in vineyards, grasslands, and maize fields in extensively and intensively managed agroecosystems[37]. However, there are conflicting reports regarding the impact of host plant identity on AMF communities. Corredor et al.[38] found that the identity of seven Salix cultivars had a greater influence on AMF communities than the health status of the host plants. In contrast, Bainard et al.[39] demonstrated that soil phosphate flux and climatic variables significantly impacted the soil AMF community structure in three different crops (pea, lentil, and wheat), with little effect from plant host identity. Similarly, studies on the rhizosphere AMF community of navel orange found that cultivars did not significantly influence the diversity and richness of the AMF community, but there was a noticeable variation in the composition of the rhizosphere AMF community across different habitats[40]. Therefore, understanding how AMF communities vary with soil depth and what factors affect their community assembly is essential for developing suitable topsoil and subsoil management strategies to maintain soil health and crop productivity.
Based on the previous research by Xing et al.[41], which investigated the composition and diversity of AMF in mulberry plants in rocky desertification areas of Guizhou, China, it is recognized that mycorrhizal colonization and species richness vary across different habitats. Given this variability, we hypothesize that there are significant differences in the diversity and community composition of native AMF populations in cultivated mulberry fields located in different agroecological zones. In light of this, six main mulberry cultivars were selected from two geographically distant locations (Gao and Ningnan counties) in Sichuan Province, a prominent sericulture hub in China. Within each location, three fields featuring diverse mulberry varieties were meticulously selected for comprehensive soil and root sampling. The objectives of the present study were: (1) to characterize the community composition of AMF naturally present in cultivated mulberry fields and identify the specific AMF species involved; (2) to determine whether mulberry varieties recruit specific AMF communities from indigenous AMF populations under field conditions; (3) to assess which factor has a more significant impact on the AMF community: differences in mulberry varieties or variations in soil properties. By addressing these objectives, we aim to gain a deeper understanding of the AMF community dynamics in cultivated mulberry fields and how they are influenced by both plant and soil factors.
-
The study was conducted in mulberry plantation zones of the Ningnan and Gao counties in Sichuan Province (China), representing two distinct ecoregions of mulberry cultivation. To control for geographic confounding factors, all selected orchards shared comparable management histories (5-year-old plantations under standardized fertilization: 300 kg N ha−1, 150 kg P2O5 ha−1, and 200 kg K2O ha−1) and were sampled during the same phenological stage (May 2018).
Ningnan County is located in the southwestern margin of the Sichuan basin (26°50'−27°18' N, 102°27'−102°55' E), with a humid subtropical monsoon climate, an average annual sunshine of 2,257.7 h, an average temperature of 19.3 °C, and an annual rainfall and evaporation of 970.1 and 1,937.9 mm respectively. Three varieties (FengTian 5, HongYa, and YunSang 1) were chosen in Ningnan County (Table 1), and every variety has three fields (20 m × 20 m). Notably, FengTian 5 fields had 10 cm toner cover as a local practice, while other varieties were grown in adjacent fields under identical climatic conditions.
Table 1. Basic information of six mulberry varieties in Sichuan Province, southwest China.
Varieties Location Taxonomic group BaiYou Gao County Morus alba L. ChuanSang 98-1 Gao County Morus alba L. QiangSang 1 Gao County Morus multicaulis Perr. HongYa Ningnan County Morus multicaulis Perr. YunSang 1 Ningnan County Morus multicaulis Perr. FengTian 5 Ningnan County Morus multicaulis Perr. 





Gao County is located on the southern edge of the Sichuan basin (28°11'−28°47' N, 104°21'−104°48' E), which also has a humid subtropical monsoon climate, sunshine, and rainfall is abundant, and the average annual temperature is about 18°C, and mean annual precipitation varies from 1,021 to 1,050 mm precipitation. Gao County has about 30 ha of mulberry trees planted. Three additional varieties (QiangSang 1, BaiYou, and ChuanSang 98-1) were sampled across nine adjacent 20 m × 20 m fields to ensure spatial homogeneity within the County.
Sampling
-
Samples were collected on May 4th and 6th, 2018, in Gao and Ningnan counties, respectively. In each field, a quadrat (2 m × 2 m) was established, and five mulberry trees with consistent growth were selected using the diagonal sampling method. After removing surface debris, soil samples were collected from two depth layers (0−10 and 10−30 cm) within 5−10 cm of mulberry tree trunks using a sterilized soil drill. To specifically target root-zone soil microbiota while minimizing endophytic microbial interference, all root fragments (> 2 mm diameter) and plant debris were surgically removed before DNA extraction, ensuring our analysis focused exclusively on free-living and root-associated (but not intracellular) microbial communities. For each tree, composite samples were created by mixing soil from four directions around each tree crown, then placed into polyethylene bags with appropriate labels and stored in a portable refrigerator. Simultaneously, fine root samples were collected, washed, and preserved in a 60% alcohol solution for subsequent mycorrhizal colonization analysis. The soil samples were divided into two portions: one part was air-dried and sieved through 1 mm and 0.25 mm screens for physicochemical analysis, while the other part was stored at −80 °C for subsequent DNA extraction.
Soil analysis
-
Soil nutrient analysis was conducted according to the methodology described by Yang et al.[42]. Soil pH was measured using an acidity meter with a soil-to-water dilution ratio of 1:5. Soil organic carbon (SOC) was determined by the K2Cr2O7 oxidation method. Soil available nitrogen (AN) was detected using the alkaline diffusion method. Soil available phosphorus (AP) was measured using the molybdenum antimony colorimetric method. Soil available potassium (AK) was extracted with 1.0 M NH4Ac and then measured with a flame photometer[42]. The soil urease activity was determined using the sodium phenol-sodium hypochlorite colorimetric method and soil sucrase activity was determined using the 3,5-dinitrosalicylic acid colorimetry method[43].
For the determination of easily-extractable glomalin-related soil protein (EE-GRSP), 1.0 g of soil was incubated with 8 mL of 20 mM sodium citrate (pH 7.0) at 121 °C for 30 min. The supernatant was then collected by centrifuging at 4 °C and 10,000 g for 6 min. Total GRSP (T-GRSP) was extracted using 8 ml of 50 mM sodium citrate (pH 8.0) at 121 °C for 60 min, and centrifuged under the same conditions. This extraction process was repeated five times, with the supernatants combined until the solution was nearly colorless. The protein content in the supernatant was assayed using the Bradford method, with bovine serum albumin serving as the standard, as described by Wright & Upadhyaya[44].
Observed mycorrhizal colonization of root
-
Mycorrhizal colonization rate was quantified as the percentage of root segments containing AMF structures (such as hyphae, arbuscules, vesicles, and/or spores), representing the degree of symbiotic association between fungal partners and host plant roots. The detailed description of the mycorrhizal colonization rate was as follows: root segments (1 cm length) were cleaned in 20% KOH for 40 to 120 min at 60 °C in a water bath and then acidified by 5% acetic acid for 5 min. Cleared roots were stained for 30 min in 5% ink-vinegar solution at 60 °C in a water bath. Roots were destained by immersing in acetic acid dye for 14 h[45].
Colonization rates were calculated using the grid-line intersection method[46]:
Vesicle infection rate = Number of vesicle infections / Total number of intersections) × 100%
Cluster infection rate = Number of cluster infections / Total number of intersection points) × 100%
Mycelium infection rate = Number of mycelium infections / Total number of intersections) × 100%
DNA extraction, PCR amplification, and Illumina MiSeq sequencing analysis
-
Soil DNA was extracted from 0.5 g of fresh root-zone soil samples using the E.Z.N.A. Soil DNA Kit (Omega Bio-tek, USA), following the manufacturer's instructions. The concentration and purity of the extracted genomic DNA were then measured using a NanoDrop NC2000 (Thermo Scientific) spectrophotometer.
The genomic DNA was amplified using a nested polymerase chain reaction (PCR) procedure. The first PCR amplification was carried out using the primer pair GeoA2 and Geo11[47] in a final reaction mixture of 25 μL, which consisted of 2.0 μL of DNA template, 1 μL each (10 μM) of GeoA2 and Geo11 primers, 5 μL of 5× reaction buffer, 5 μL of 5× GC buffer, 2 μL of dNTPs (2.5 mM), 0.25 μL of Q5 DNA Polymerase, and 8.75 μL of sterile water. The amplification conditions were as follows: an initial denaturation at 98 °C for 3 min, followed by 25−30 cycles of denaturation at 98 °C for 15 s, annealing at 55 °C for 30 s, extension at 72 °C for 30 s, and a final extension period at 72 °C for 10 min.
The second PCR amplification was performed using the primer pair AMV4.5NF and AMDGR[48], with conditions similar to those used for the first PCR. The PCR products from the second step were loaded onto a 1.0% agarose gel containing PicoGreenTM (Invitrogen) to confirm the success of amplification.
All PCR products were purified using Vazyme VAHTSTM DNA Clean Beads and quantified using a Microplate reader (BioTek, FLx800). The qualified samples were then sequenced on the Illumina MiSeqTM System platform at Personalbio Company (Shanghai, China) using a 250 bp paired-end strategy.
The resulting high-throughput sequences were clustered into OTUs using the QIIME2 dada2 analysis process[49]. Briefly, paired-end reads were first separated into individual samples based on barcodes and then filtered to remove chimeric, low-quality sequences, primer sequences, and barcode sequences. The valid reads were then clustered into OTUs using a 97% identity threshold. The most abundant sequence from each OTU was selected as the representative sequence for that OTU. These representative sequences were compared to the online MaarjAM database (
http://maarjam.botany.ut.ee ; status on 12 April 2015)[50] for taxonomic assignment using the BLAST algorithm with the following parameter settings: 97% similarity, coverage > 95%, and e-value < 1e-50. The alignment results grouped the representative sequences per OTU obtained in this study into corresponding molecular virtual taxa species (VTs).Bioinformatics and statistical analysis
-
Statistical analyses were performed using SPSS 19.0 (IBM Corp., Armonk, NY, USA). Data are presented as means ± standard error (SE, n = 3). One-way ANOVA with Tukey's honestly significant difference (HSD, p < 0.05) post hoc test was used to assess varietal differences in: (1) soil physicochemical properties, (2) AMF colonization rates, and (3) fungal relative abundance. Before ANOVA, data normality was confirmed by Shapiro-Wilk test (p > 0.05) and homogeneity of variance by Levene's test (p > 0.05). All figures were generated using Origin 2018 (OriginLab Corp., Northampton, MA, USA).
Sequence data analyses were mainly performed by using QIIME2 and R (version 3.6.1) packages. The relative abundance of a genus in each sample was calculated by dividing the OTU numbers of the genus in a sample by the total OTU numbers for that sample. A flower plot was generated to visualize the unique and common VTs among different mulberry varieties. Alpha diversity indices, including Chao1, Observed species, Shannon diversity index, and Simpson diversity index, were calculated using QIIME2. Beta diversity was analyzed to investigate the structural variation of AMF communities between samples using Principal Coordinate Analysis (PCA) with Bray-Curtis distance measurements. Hellinger transformation-based Redundancy Analysis (tbRDA) was utilized to estimate the proportion of AMF community variance that could be explained by explanatory variables across mulberry varieties. This analysis also helped to discern the variance contributions from varietal differences and soil property differences. To mitigate multicollinearity among explanatory variables, forward selection was applied to select the simplest variable combination that could substantially explain AMF community variance while avoiding strong correlations with filtered variables. The statistical significance of these analyses was tested using Student's t-test and the Monte Carlo permutation test with 1,000 permutations.
-
The contents of EE-GRSP and T-GRSP in the soil exhibited significant variation among six mulberry varieties (Fig. 1a, b). For EE-GRSP, QiangSang 1 demonstrated the highest content, whereas YunSang 1 recorded the lowest levels in both the 0−10 and 10−30 cm soil layers. In terms of T-GRSP, FengTian 5 showed the highest content within the 0−10 cm layer, while QiangSang 1 had the greatest concentration in the 10−30 cm layer. Both YunSang 1 and ChuanSang 98-1 displayed lower levels across both layers. Notably, both EE-GRSP and T-GRSP contents decreased with increasing soil depth, except for BaiYou Changsang 98-1 and QiangSang. Furthermore, it is important to highlight that EE-GRSP content was significantly higher in Gao County compared to Ningnan County across both soil layers (Fig. 1a, b).
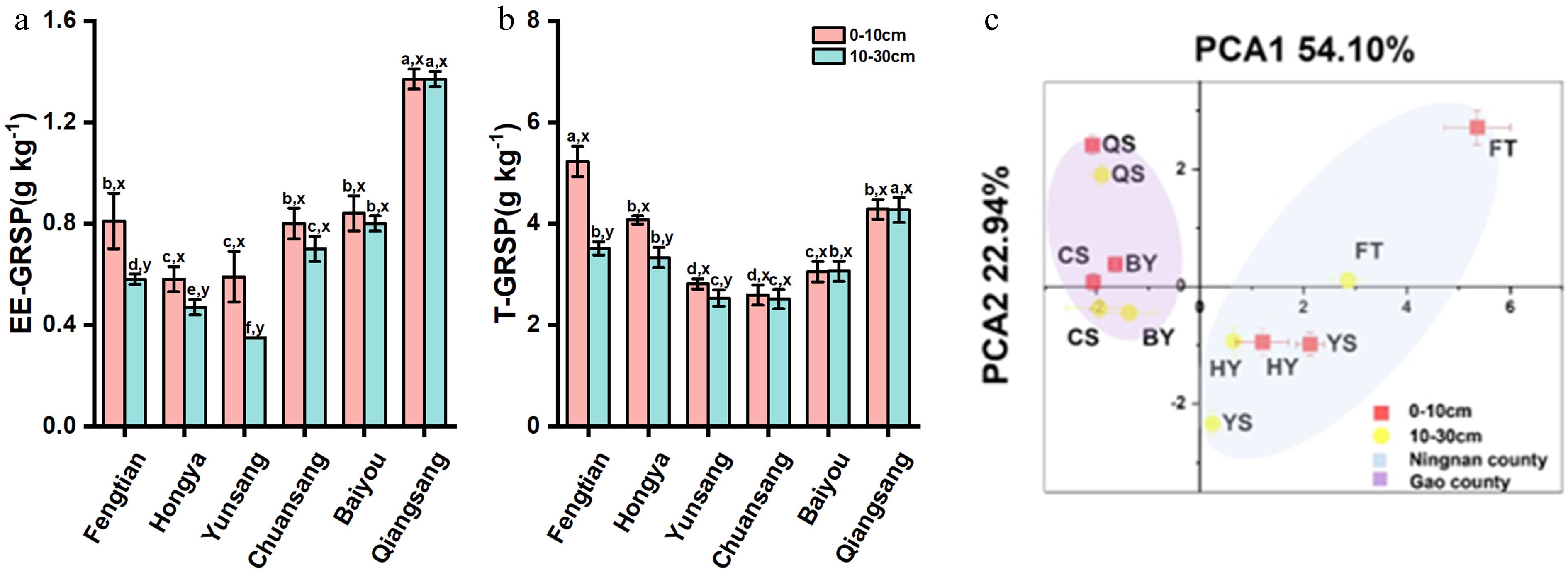
Figure 1.
(a) Variations of easily-extractable glomalin-related soil protein (EE-GRSP), and (b) total GRSP (T-GRSP) between different varieties of mulberry orchids across two soil layers. (c) Soil fertility index load chart in the rhizosphere soil of different varieties mulberry across two soil layers from Gao County and Ningnan County, Sichuan Province, China. Data (means ± SE, n = 3) followed by different letters (a, b, c, d) indicate significant differences among mulberry varieties within the same soil layer (p < 0.05) and different lowercase (x, y) means significant differences between different soil layers in mulberry fields of the same variety (p < 0.05). FT: FengTian; HY: HongYa; YS: YunSang; CS: ChuanSang; BY: BaiYou; QS: QiangSang.
The soil pH showed clear geographic differentiation, with Ningnan County ranging from 7.2 to 7.4 (neutral) and Gao County from 4.9 to 5.5 (acidic) (Table 2). Among the mulberry varieties, significant changes in soil pH across soil layers were only observed for YunSang 1 and BaiYou. Significant variations in SOC, AN, AP, AK, and sucrase activities were observed among the varieties (Table 2). FengTian 5 had exceptionally high levels of SOC, AP, and AK, while ChuanSang 98-1 had the lowest SOC and AK, and QiangSang 1 had the lowest AP in both soil layers. In terms of AN content, ChuanSang 98-1 ranked highest, followed by BaiYou, QiangSang 1, HongYa, FengTian 5, and YunSang 1 in the 0−10 cm layer; in the 10−20 cm layer, the order was ChuanSang 98-1, BaiYou, FengTian 5, QiangSang 1, HongYa, and YunSang 1. Nutrient contents were higher in the 0−10 cm layer than in the 10−30 cm layer. Yuansang 1 exhibited the highest sucrase and urease activities, while QiangSang 1 showed the lowest (Table 2).
Table 2. Variations in soil chemical properties and enzymatic activities within the rhizosphere of different varieties mulberry across two soil layers.
Variables Soil depth (cm) Gao County Ningnan County F p BaiYou ChuanSang 98-1 QiangSang 1 FengTian 5 HongYa YunSang 1 pH 0−10 5.3 ± 0.2b, y 5.5 ± 0.1b, x 4.9 ± 0.2b, x 7.2 ± 0.2a, x 7.4 ± 0.2a, x 7.2 ± 0.2a, x 97.60 < 0.001 10−30 6.3 ± 0.1b, x 5.4 ± 0.2c, x 5.0 ± 0.2c, x 7.4 ± 0.3a, x 6.9 ± 0.2a, x 6.7 ± 0.2a, y 145.16 < 0.001 SOC (g/kg) 0−10 9.7 ± 0.1e, x 8.0 ± 0.6f, x 17.5 ± 0.7b, x 118.7 ± 28.6a, x 10.5 ± 0.1d, x 14.3 ± 0.1c, x 41.95 < 0.001 10−30 9.3 ± 0.2c, x 7.1 ± 0.2c, y 15.4 ± 0.3b, y 69.6 ± 10.9a, y 7.8 ± 0.3d, y 6.9 ± 0.2d, y 93.88 < 0.001 AN (mg/kg) 0−10 123.0 ± 3.7a, x 136.0 ± 11.0a, x 115.7 ± 3.0b, x 88.5 ± 5.9d, x 104.2 ± 6.5c, x 84.3 ± 12.5d, x 15.89 < 0.001 10−30 83.0 ± 4.7b, y 107.5 ± 10.6a, y 63.6 ± 6.1b, y 79.8 ± 11.4b, x 52.5 ± 15.4c, y 38.3 ± 10.3c, y 14.55 < 0.001 AP (mg/kg) 0−10 62.2 ± 4.5d, x 136.1 ± 14.6c, x 25.9 ± 2.6e, x 170.9 ± 16.4a, x 166.0 ± 8.3c, x 257.8 ± 11.2b, x 227.76 < 0.001 10−30 32.7 ± 2.0d, y 112.2 ± 2.7b, y 16.7 ± 3.1e, y 95.5 ± 5.3a, y 50.7 ± 1.6c, y 25.9 ± 3.3d, y 785.51 < 0.001 AK (mg/kg) 0−10 47.7 ± 2.1d, x 37.7 ± 0.6e, x 52.3 ± 1.5d, x 179.7 ± 7.5a, x 102.0 ± 2.0c, x 115.3 ± 2.1b, x 741.98 < 0.001 10−30 40.0 ± 1.0e, y 34.7 ± 1.5f, y 46.3 ± 1.5d, y 126.5 ± 2.6a, y 82.3 ± 0.6c, y 94.7 ± 2.1b, y 1371.3 < 0.001 Sucrase (mg/g/h) 0−10 0.41 ± 0.10c, x 0.15 ± 0.16d, x 0.11 ± 0.00d, x 2.55 ± 0.21a, x 1.53 ± 0.30b, x 2.81 ± 0.42a, x 30.94 < 0.001 10−30 0.10 ± 0.08c, y 0.05 ± 0.00c, y 0.06 ± 0.01c, y 1.58 ± 0.23a, y 1.38 ± 0.06a, x 0.80 ± 0.06b, y 48.96 < 0.001 Urease (mg/g/h) 0−10 0.05 ± 0.00c, x 0.04 ± 0.00d, x 0.03 ± 0.00d, x 0.14 ± 0.00b, x 0.14 ± 0.00b, x 0.20 ± 0.00a, x 87.63 < 0.001 10−30 0.04 ± 0.00d, x 0.04 ± 0.00d, x 0.04 ± 0.00d, x 0.12 ± 0.01a, y 0.08 ± 0.01b, y 0.06 ± 0.01c, y 92.78 < 0.001 Values are the means ± SE, n = 3. Different lowercase letters (a, b, c, d) mean significant differences among mulberry varieties within the same soil layer (p < 0.05) and different lowercase letters (x, y) mean significant differences between different soil layers in mulberry fields of the same variety (p < 0.05). Principal Component Analysis (PCA) revealed similarities in soil properties between ChuanSang 98-1 and BaiYou, as well as between YunSang 1 and HongYa (Fig. 1c). Notably, FengTian 5 had the highest soil fertility value while ChuanSang had the lowest, with Ningnan County overall showing higher fertility than Gao County (Fig. 1c).
Observed mycorrhizal colonization rate of six mulberry varieties
-
To confirm the observed colonization rate of two tea varieties with AMF The root mycelial infection rates for FengTian 5, HongYa, YunSang 1, and ChuanSang 98-1 were all greater than 40%, with no statistically significant differences among these four varieties. This was followed by QiangSang 1 (38% ± 2.1%) and BaiYou (33% ± 3.2%) (Fig. 2a). Additionally, there were no significant differences in arbuscular infection rates among FengTian 5, HongYa, ChuanSang 98-1, and BaiYou (24%−30% ± 2.2%−3.7%), but these rates were significantly higher than those observed for YunSang 1 and QiangSang 1 (18%−19% ± 2.0%−2.2%) (Fig. 2b). Furthermore, the vesicle infection rate was highest in ChuanSang 98-1 (35% ± 0.9%), which was substantially higher than that observed for the other five varieties (12%−18% ± 1.9%−3.3%) (Fig. 2c).
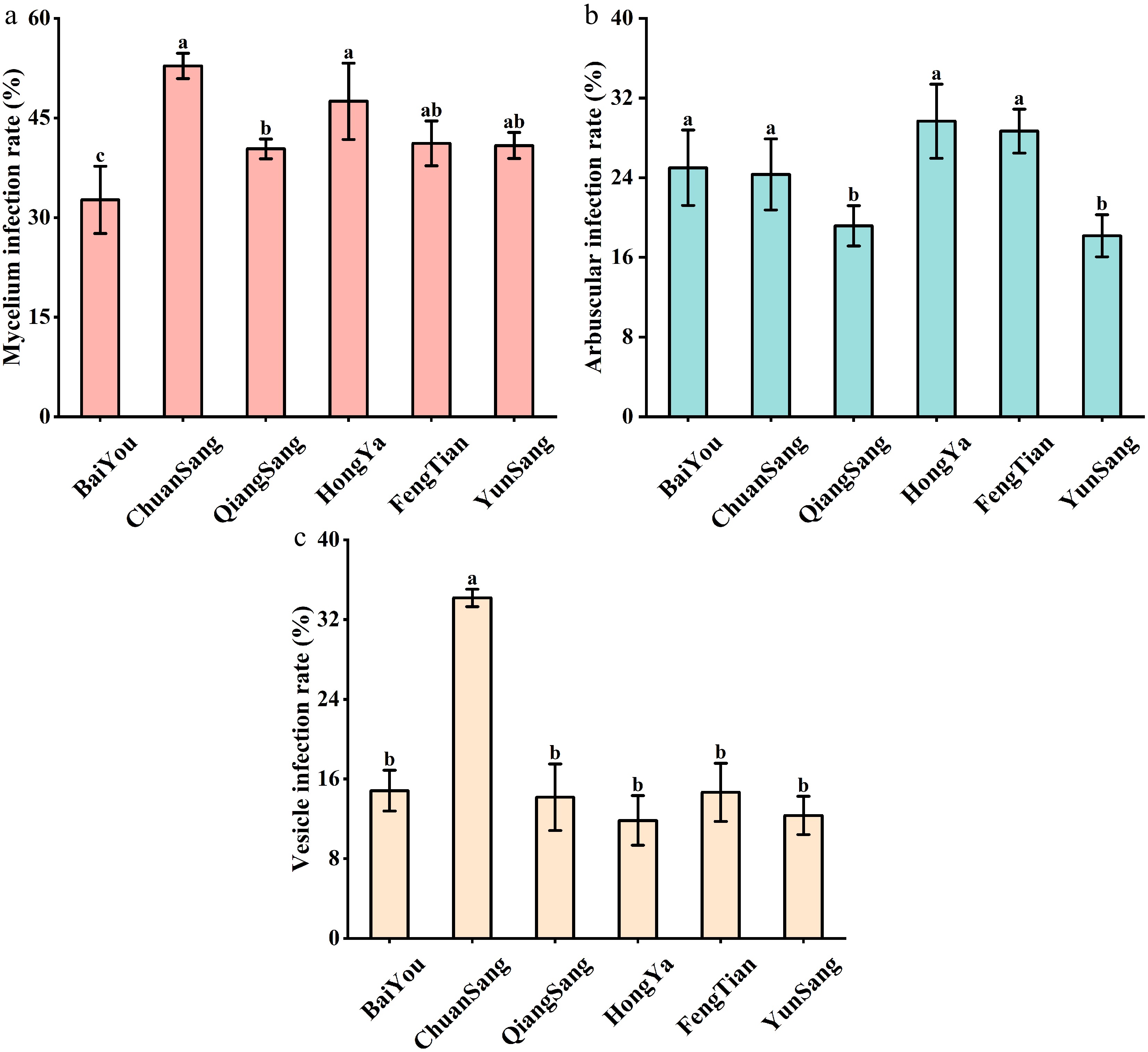
Figure 2.
Variations in observed (a) mycelium colonization, (b) arbuscular colonization, and (c) vesicle colonization in different cultivars of mulberry orchards. Data (means ± SE, n = 3) followed by different letters (a, b, c, d) indicate significant differences among mulberry varieties (P < 0.05).
AMF community composition in different varieties of mulberry
-
To evaluate the root-zone soil AMF community structure, we conducted a comparative analysis of AMF composition and abundance in the root-zone soil of mulberry plants, as well as the co-occurrence of AMF communities across various locations and varieties. A total of 1,343 AMF OTUs were classified into seven genera within the phylum Glomeromycota, namely Archaeospora, Ambispora, Claroideoglomus, Glomus, Gigaspora, Paraglomus, and Scutellospora. Notably, Glomus and Paraglomus were the dominant genera, accounting for approximately 61% and 35% of the total OTUs, respectively (Fig. 3a).
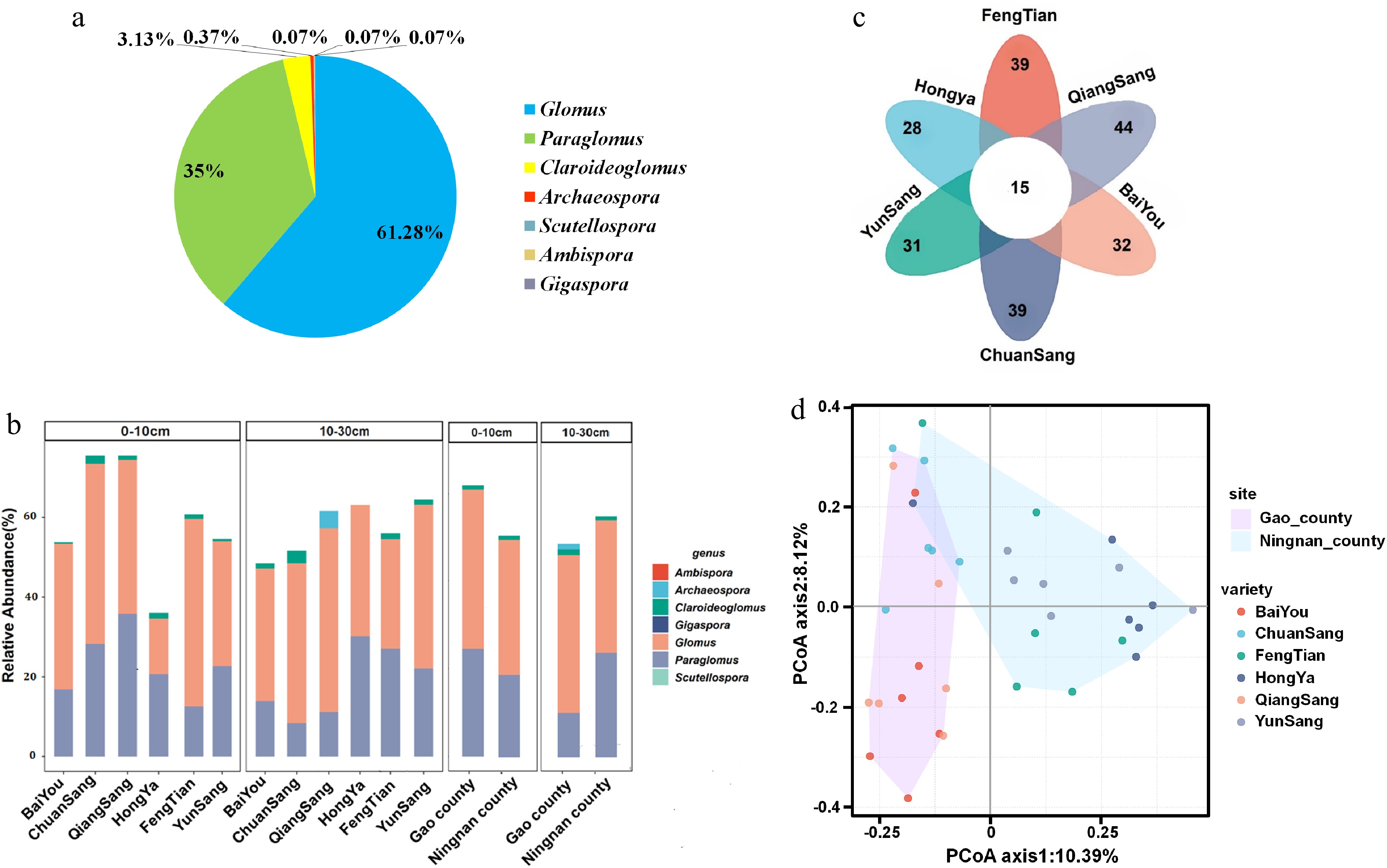
Figure 3.
(a) Composition of AMF communities in the rhizosphere soil of mulberry from Gao and Ningnan Counties, Sichuan Province. (b) Relative abundances of AMF genus in the rhizosphere soil of mulberry across two soil layers from Gao and Ningnan Counties, Sichuan Province. (c) Numbers of the Virtual taxa species in the rhizosphere soil of six different varieties mulberry. (d) Principal coordinate analysis (PCoA) based on OTUs results showing community dissimilarities between different mulberry varieties.
The relative abundance of AMF genera varied among different mulberry varieties, with the differences in AMF composition in soil primarily attributed to Glomus and Paraglomus (Fig. 3b). In the topsoil, ChuanSang 98-1 and QiangSang 1 exhibited significantly higher relative abundance compared to other varieties, followed by FengTian 5, YunSang 1, and BaiYou. In contrast, HongYa showed a significantly lower abundance. In the subsoil, no significant differences were observed in the relative abundance among all varieties. Notably, the Archaeospora genus was detected in QiangSang 1 at 10−30 soil layer but was absent in other varieties. Futhermore, the relative abundance of Paraglomus decreased with increasing soil depths in BaiYou, ChuanSang 98-1, and QiangSang 1. Conversely, the relative abundance of Paraglomus increased in FengTian 5, while that of Glomus decreased in Fengtiaan 5 and YunSang 1 with increasing soil depth. Interestingly, both Paraglomus and Glomus showed an increase in relative abundance with increasing soil depth in HongYa. Geographic comparisons showed significantly higher AMF abundance in Gao County topsoil but lower abundance in subsoil compared to Ningnan County.
Virtual taxa distribution
-
We identified an additional 54 VT (virtual taxa) species (Supplementary Table S1). The abundance of VT species observed ranked as follows: QiangSang 1 (45) > ChuanSang 98-1 (39) = FengTian 5 (39) > BaiYou (32) > YunSang 1 (31) > HongYa (28). There was a higher abundance of VT species in Gao County compared to Ningnan County. Fifteen VT species were common to all six mulberry varieties (Fig. 3c, Supplementary Table S1). Regarding the detailed composition of VT species in the soils of different mulberry varieties, the data indicate that the soil of QiangSang 1 was richest in Glomus VT species (37), whereas the soil of HongYasang had the fewest Glomus VT species (20). Additionally, the soils of HongYasang, YunSang 1, and ChuanSang 98-1 contained the highest number of Paraglomus VT species (6), while the soil of QiangSang No. 1 had the lowest number of Paraglomus VT species (4). In the soil of ChuanSang 98-1, the Claroideoglomus VT species exhibited the greatest diversity, with four species present; in contrast, the soils of BaiYousang and YunSang 1 had the least Claroideoglomus VT species, with only one species found in each. Further analysis revealed that 15 common VT species across the six mulberry varieties showed that 12 belonged to the Glomus genus, two belonged to the Paraglomus genus, and the remaining one belonged to the Claroideoglomus genus (Supplementary Table S1). These findings indicated the composition and abundance of VT species varied significantly among the mulberry varieties.
Alpha diversity in different varieties of mulberry
-
The investigation into soil AMF alpha diversity revealed significant variations among different mulberry varieties (Fig. 4). Specifically, the AMF diversity indices, including observed species index, Chao1 index, Shannon index, and Simpson index, were significantly higher in topsoil than in subsoil, with the exception of ChuangSang 98-1 and BaiYou. In the topsoil, the observed species index, Chao1 index, and Shannon index were ranked as follows: HongYa ≥ FengTian 5 > YunSang1 ≥ ChuanSang 98-1 ≥ QiangSang 1 ≥ BaiYou. In contrast, in the subsoil, the Shannon index and observed species index were highest in ChuanSang 98-1, followed by HongYa, BaiYou, FengTian 5, and YunSang 1, with the lowest value observed in QiangSang 1. The Chao1 index didn't show significant differences among six mulberry varieties in the subsoil. Additionally, across both soil layers, the Simpson index was equally high for ChuanSang98-1, FengTian5, HongYa, and YunSang1, followed by BaiYou, and then QiangSang1 showing the lowest value. Consequently, the AMF diversity indices were found to be higher in Ningnan County compared to Gao County (Fig. 4).
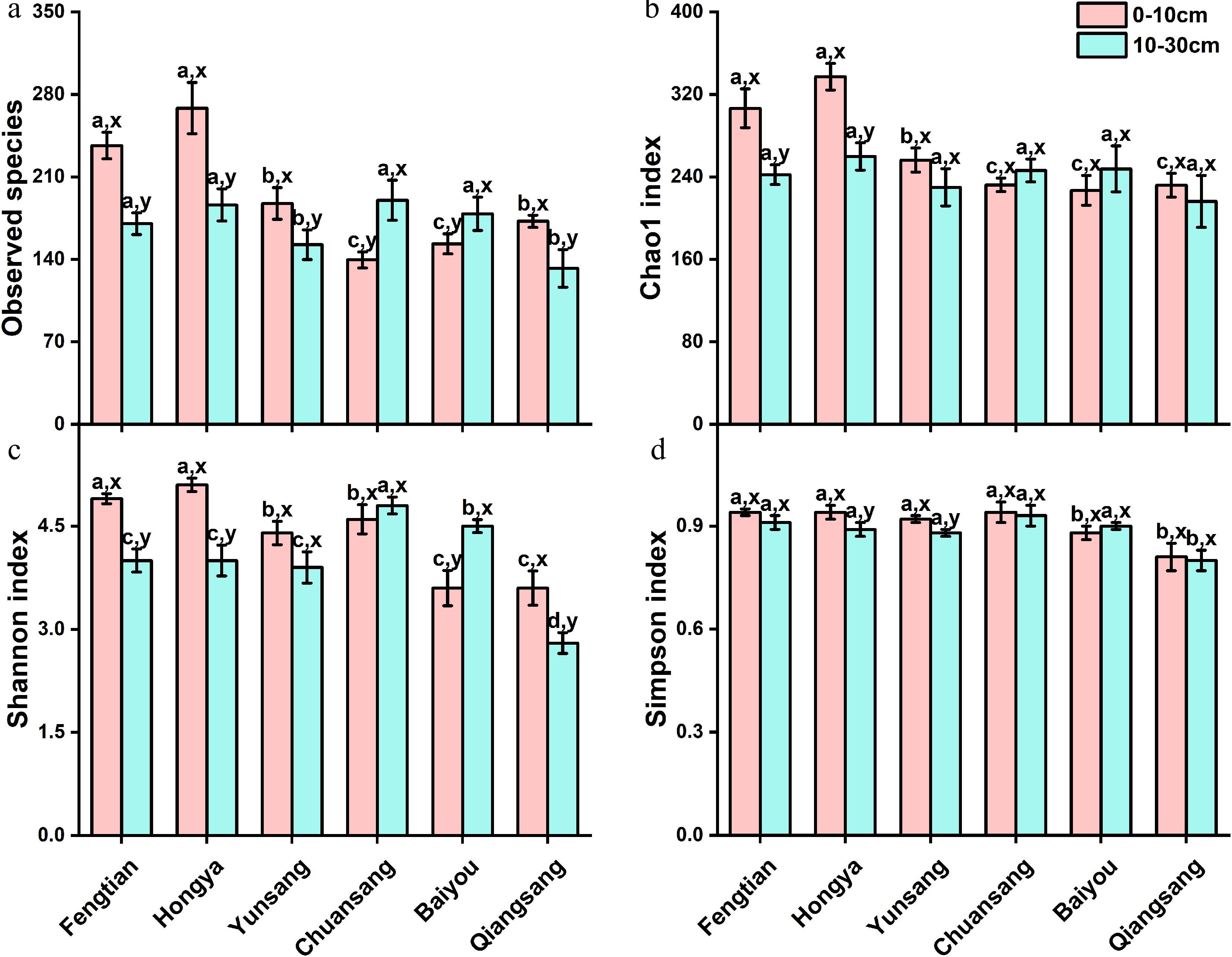
Figure 4.
Alpha diversity indices, (a) Chao 1, (b) observed species, (c) Simpson, and (d) Shannon of AMF in the rhizosphere soil of mulberry across two soil layers from Gao and Ningnan counties, Sichuan Province. Data (means ± SE, n = 3) followed by different letters (a, b, c, d) indicate significant differences among mulberry varieties within the same soil layer (p < 0.05) and different lowercase (x, y) means significant differences between different soil layers in mulberry fields of the same variety (p < 0.05).
Geographic pattern synthesis
-
The distinct soil conditions between counties (acidic in Gao vs neutral in Ningnan) corresponded with differential AMF community patterns. While Glomus dominated in both locations, its relative abundance was higher in Gao County (Fig. 3b), potentially linked to the observed higher GRSP levels in acidic soils. Conversely, Ningnan's neutral pH soils showed greater representation of other genera. However, varietal differences maintained significance within each county, as evidenced by consistent patterns like ChuanSang 98-1's high AN content across both locations (Table 2). This suggests that while geographic factors influence soil properties and microbial communities, host varieties remain a primary driver of AMF composition.
The relationship between the AMF community with varieties and soil properties
-
The results of PCA revealed significant differences in soil AMF community structure among six mulberry varieties across two regions (Fig. 3d). The distinct clustering patterns also indicated clear regional differences in AMF communities between Gao County and Ningnan County. Specifically, the PCA plot shows that FengTian 5, HongYa, and YunSang 1 in Ningnan County exhibited high similarity in AMF composition, while QiangSang 1, BaiYou, and ChuanSang 98-1 in Gao County showed significant variance in AMF community composition (Fig. 3d). Furthermore, Permutational Multivariate Analysis of Variance (PERMANOVA) statistical analysis demonstrated that the AMF community composition of QiangSang 1 significantly differed from BaiYou, HongYa, YunSang 1, and FengTian 5 (p = 0.02−0.04, Table 3), but not from ChuanSang 98-1 (p = 0.08, Table 3). Significant differences were observed between BaiYou and HongYa as well as YunSang 1 (p = 0.015−0.03, Table 3); however, no significant difference was found between BaiYou and ChuanSang 98-1, or FengTian 5 (p = 0.11−0.18, Table 3). Additionally, significant differences were noted between ChuanSang 98-1 and YunSang 1, but not between ChuanSang 98-1 and HongYa or FengTian 5, nor between HongYa and YunSang 1 or FengTian 5. Thus, among the six varieties, the AMF compositions of ChuanSang 98-1, BaiYou, and QiangSang 1 in Gao County exhibited significant differences compared to other varieties. Conversely, the AMF compositions of HongYa, YunSang 1, and FengTian 5 in Ningnan County showed greater similarity with limited specificity.
Table 3. PERMANOVA statistical results of AMF communities among six mulberry varieties.
Group F model R2 Padj QiangSang 1 BaiYou 1.94 0.16 0.045 ChuanSang 98-1 2.00 0.17 0.075 HongYa 2.55 0.20 0.015 YunSang 1 2.31 0.19 0.045 FengTian 5 2.26 0.18 0.045 BaiYou ChuanSang 98-1 1.88 0.16 0.18 HongYa 2.75 0.22 0.03 YunSang 1 2.44 0.20 0.015 FengTian 5 2.21 0.18 0.105 ChuanSang 98-1 HongYa 2.26 0.18 0.195 YunSang 1 2.10 0.17 0.045 FengTian 5 2.14 0.18 0.21 HongYa YunSang 1 1.12 0.10 1 FengTian 5 1.43 0.13 1 YunSang 1 FengTian 5 1.62 0.14 0.165 Driving factors for AMF richness and diversity
-
Redundancy analysis (RDA) of Hellinger-transformed AMF community data revealed that soil properties and mulberry varieties collectively explained 50.7% of the observed variation in fungal composition (F = 1.54, p < 0.001; Fig. 5a). Variance partitioning analysis demonstrated that soil characteristics alone accounted for 26.4% of the variation, while host varieties explained 14.7%, with an additional 9.6% attributed to their interactive effects (Fig. 5c). Monte Carlo permutation tests showed distinct significance patterns: although no individual soil parameter exhibited significant independent effects (all p > 0.05; Table 4), mulberry cultivars exerted strong deterministic selection on AMF assemblages (F = 1.25, p = 0.006). When controlling for varietal effects, soil-specific RDA still explained 36.0% of community variation (F = 1.62, p < 0.001; Fig. 5b), with AP (F = 1.63, p = 0.004), AK (F = 1.94, p = 0.003), SOC (F = 1.38, p = 0.023), and T-GRSP: (F = 1.67, P = 0.003) emerging as significant drivers (Table 4). These results demonstrate a hierarchical assembly process where host genotypes dominate initial AMF recruitment through strong selective pressure (high statistical significance), while soil properties subsequently shape communities through broader environmental filtering (greater explanatory power but distributed across multiple factors). The substantial interactive component (9.6%) highlights the complex interdependence between biotic selection and abiotic constraints in structuring AMF communities.
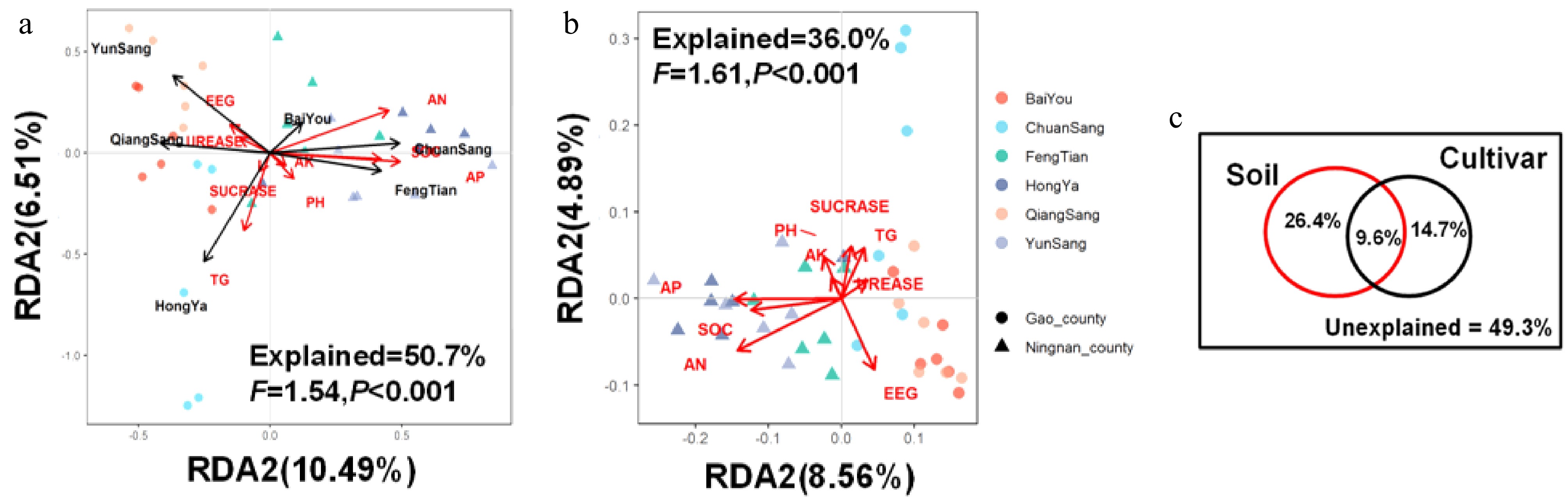
Figure 5.
Hellinger transformation-based redundancy analysis (tbRDA) of AMF abundance with (a), (b) soil properties, and (c) variation partitioning.
Table 4. Monte Carlo permutation tests were used to detect the variance explained by varieties and soil properties.
Explaining variable RDA model explained By variety and soil R2 = 0.507 By soil R2 = 0.350 Variance F Pr (> F) Variance F Pr (> F) Variety 0.115 1.25 0.006 − − − AK 0.019 1.05 0.34 0.037 1.94 0.003 T-GRSP 0.020 1.09 0.28 0.032 1.67 0.003 Urease 0.023 1.27 0.07 0.023 1.18 0.147 AP 0.016 0.85 0.85 0.031 1.63 0.004 EE-GRSP 0.018 0.95 0.60 0.024 1.24 0.100 SOC 0.021 1.13 0.22 0.027 1.38 0.023 pH 0.022 1.19 0.14 0.021 1.08 0.292 Sucrase 0.020 1.11 0.24 0.020 1.02 0.401 AN 0.022 1.17 0.14 0.021 1.10 0.233 To explore relationships among mulberry variety, soil properties, and AMF richness and diversity, a structural equation model (SEM) was constructed (Fig. 6). The SEM results indicated that mulberry varieties exert significant direct effects on AP, AK, and SOC levels in soil, while indirect effects on T-GRSP. T-GRSP, in turn, had a significant direct impact on SOC. Furthermore, AP and AK had a direct negative effect on the AMF diversity, while SOC exerted a notable direct positive effect on soil AMF abundance. T-GRSP also had indirect positive effects on soil AMF abundance and diversity, however, these effects were not significant.
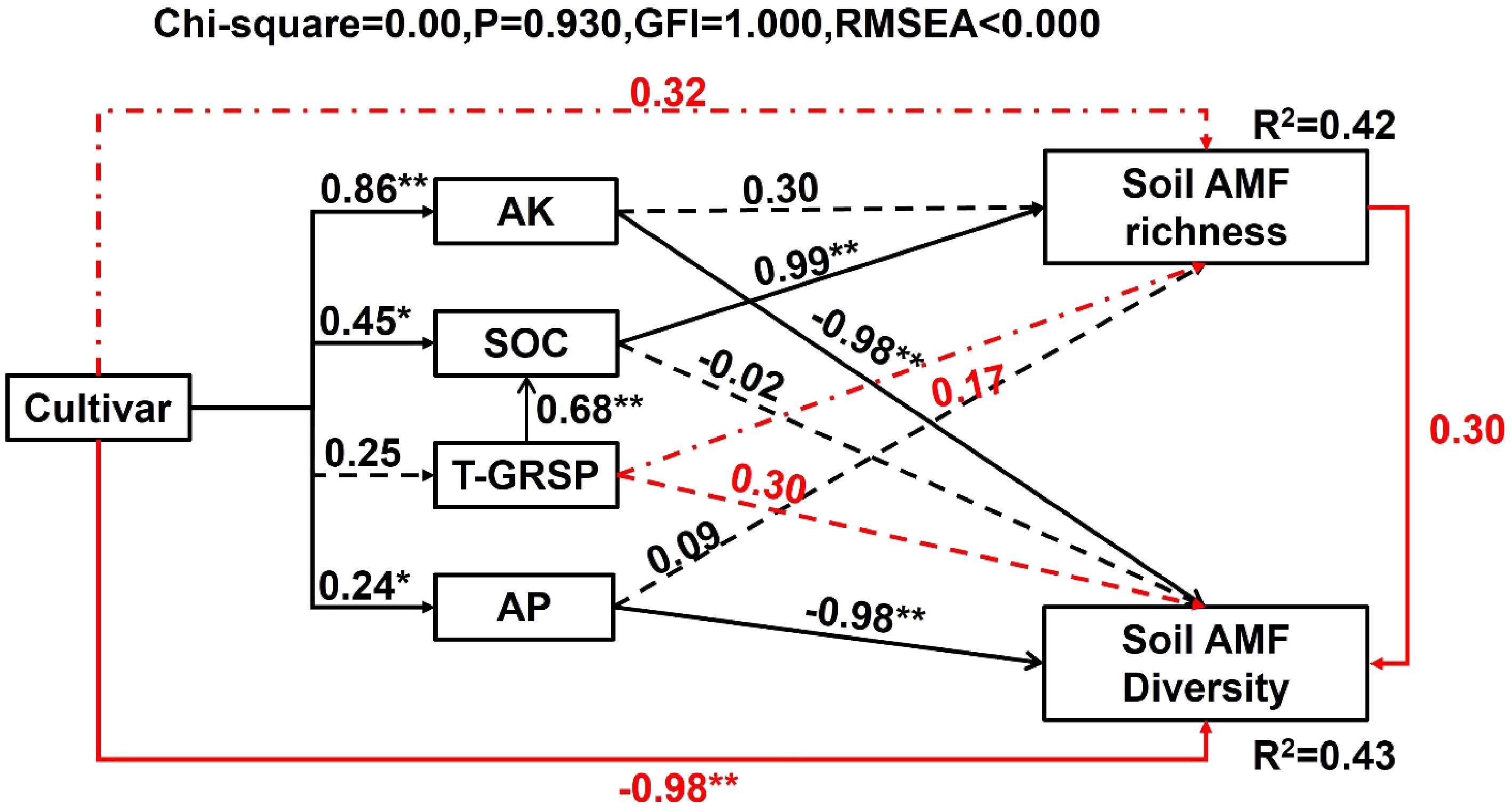
Figure 6.
Structural equation model (SEM) showing direct and indirect effects of cultivar and AP, AK, SOC, and T-GRSP on AMF richness and diversity. The solid lines indicate significant effect; dashed lines indicate insignificant effect. The black and red lines indicate direct and indirect effect, respectively. The number on each line indicate path coefficients; and R2 values represent the proportion of variance explained for AMF richness and diversity (n = 36).
-
In the present study, we employed a combination of morphological analysis and molecular genetics to explore the diversity and community compositions of native AMF in mulberry soils. Our findings revealed that, in Ningnan County, there was no statistically significant difference in the mycorrhizal infection rate among the roots of FengTian 5, HongYa, and Yunshang 1 varieties. However, in Gao County, significant variations in mycorrhizal colonization percentages were observed among Chuanshang, Qiangshang 1, and BaiYou varieties (Fig. 2). These differences may be attributed to the distinct mulberry series, which possess significant differences in their genetic backgrounds, potentially influencing the extent of root colonization by AMF. Consistent with these observations, Singh et al. inoculated five durum wheat cultivars under low and medium fertility conditions and identified cultivar type as a critical factor affecting root colonization percentages at both soil fertility levels[51]. Specifically, the cultivars exhibiting significantly lower or higher root colonization percentages depended on the soil fertility levels. In our study, the cultivar 'Chuanshang' displayed the highest percentage of arbuscular colonization (Fig. 2). Notably, soil PCA results indicated the soil fertility where Chuanshang grows is relatively poor (Fig. 1c). Despite this, the relative abundance of AMF, especially the Glomus genus, was higher in Chuanshang 98-1 compared to other varieties (Fig. 3b). This finding aligns with previous research indicating that better AM symbiotic development occurs at low fertility levels, both in domesticated crops and their wild progenitors[52,53]. Moreover, the soil fertility in Ningnan County is better than Gao County (Fig. 1c). In contrast, the relative abundance in the 0−10 cm soil layer was higher in Gao County than Ningnan County (Fig. 2). These results indicated that AMF formation will be inhibit under extremely fertile soil conditions[16,54−56], and Chuanshang may have adapted to thrive in low-fertility soils through enhanced AMF colonization. While our data reveal varietal differences in colonization intensity, future work should couple microscopic assessments with molecular analysis of root-associated AMF communities to determine whether specific fungal taxa preferentially associate with 'Chuanshang'.
AMF community composition of mulberry orchards
-
The symbiotic relationship between AMF and woody plants has garnered increasing attention, particularly due to AMF's potential to enhance the water and nutrient uptake capabilities of their host plants[57−61]. However, comprehensive studies investigating the AMF community structure in the root-zone soil of mulberry plants under field conditions remain limited. Our study identified seven AMF genera in the root-zone soil of six different mulberry varieties cultivated under field conditions in the Gao and Ningnan Counties of Sichuan Province, including Ambispora, Archaeospora, Claroideoglomus, Glomus, Gigaspora, Paraglomus, and Scutellospora (Fig. 3). Notably, Glomus and Paraglomus were the predominant genera, accounting for approximately 61% and 35% of the total OTUs, respectively. These findings align with previous research on dominant AMF genera in mulberry soils across various ecosystems. For instance, Shi et al. morphologically characterized a total of 16 AMF morphotypes from the mulberry rhizosphere soil in the rock desertification area of Chongqing, southwest China, including 10 Glomus species, five Acaulospora species, and one Gigarspora species[62]. Similarly, Xing et al. identified eight genera in karst soils of mulberry fields in Guizhou Province, southwest China, with Glomus being the dominant genus in both roots and soil (accounting for 68% of the species detected), followed by lower abundances of Ambispora, Archaeospora, Acaulospora, Diversispora, Redeckera, Claroideoglomus, and Paraglomus. They also found that converting cropland to mulberry forests benefited AMF diversity and abundance[41]. Furthermore, Zheng conducted high-throughput 454-sequencing analysis and identified five genera (Diversispora, Glomus, Gigaspora, Paraglomus, and Scutellospora) in the rhizospheric soil of mulberry plantations of varying ages (5, 10, 15, 25, and 35 years) in Shanxi Province, northwest China[63]. Glomus also emerged as the dominant genus in this study. Collectively, these findingssuggest that Glomus exhibits a competitive advantage in mulberry fields,potentially due to its high sporulation rates and rapid colonization ability[64,65]. This competitive edge could explain its dominance across different mulberry ecosystems.
The effect of mulberry variety on AMF community assembly
-
It is plausible that the AMF community can be significantly influenced not only by host species[66], but also by various environmental factors, including soil characteristics[56,67], land-use intensity[68−70], fertilization practices[35,71], and vegetation status of the study areas[41]. Our findings indicate that AMF abundance (Fig. 3b) and diversity (Fig. 4) varied significantly across different mulberry varieties. Specifically, RDA revealed that mulberry variety was the dominant factor for influencing variance in AMF community structure (Fig. 5). This aligns with previous studies, such as the study by Song et al.[72], who suggested that the genotype of the scion has a more significant impact on AMF community in citrus than the rootstock, where physical root-AMF associations occur. Similarly, Xiao et al.[73] observed that the AMF community composition is more sensitive to Indigofera atropurpurea compared to Amorpha fruticosa planting, indicating that different legume species exhibit varying abilities to adapt to microenvironmental changes. Zhang et al. further corroborated this finding in four independent tea gardens, revealing that the relative abundance of Acaulospora, Gigaspora, Glomus, and Rhizophagus in AMF was significantly higher in Zhongcha108 compared to Longjing43, suggesting that tea varieties are also the main factor affecting the rhizosphere microbial community[74]. In our study, among the six mulberry varieties studied, QiangSang 1, HongYa, YunSang 1, and FengTian 5 belong to Morus multicaulis perr., whereas BaiYou and ChuanSang 98-1 belong to Morus alba L. (Table 1). AMF community compositions were significantly different among three varieties planted in Gao County (QiangSang 1, BaiYou, and ChuanSang 98-1) but not among those in Ningnan County (FengTian 5, HongYa, YunSang 1) (Fig. 3d). This suggests that the distinct genetic backgrounds of the three varieties planted in Gao County may account for the more pronounced differences in soil AMF composition, even though they were planted in the same habitat. Additionally, significant differences in soil AMF composition were also observed between QiangSang 1 planted in Gao County and the three varieties planted in Ningnan County, despite them belonging to the same mulberry species. This likely reflects both intrinsic genetic divergence and environmental modulation, where Gao's acidic soils (pH 4.9−5.5) may amplify subtle varietal differences in AMF recruitment through pH-dependent changes in root exudation or fungal viability. Notably, these host-driven patterns persist despite county-level variations in absolute abundance (e.g., elevated Glomus in Gao County), with variety-specific signatures like QiangSang 1's exclusive Archaeospora association remaining stable across soil depths. Such robust genetic control aligns with recent findings in tea[74], and citrus[72] systems, suggesting perennial crops maintain strong host filtering of AMF communities. While our data demonstrate clear varietal associations, long-term manipulative experiments across diverse soil types are needed to fully disentangle these interactions.
The impact of soil characteristics on AMF abundance, diversity, and community composition
-
The abiotic environment factor exhibits variable effects on AMF community composition[75−77]. Different plant species exhibit varying mycorrhizal dependencies and produce distinct root exudates, which subsequently influence soil properties during their growth phases[78,79]. In this study, we observed that not only mulberry variety but also soil AP, AK, SOC, and T-GRSP significantly influenced AMF community structure (Figs 5b, 6, Table 4). Notably, soil AP has consistently been recognized as a critical determinant of AMF community composition. High levels of soil P can negatively impact the transduction of phosphatase exuded by AMF, thereby inhibiting AMF growth[80]. Xu et al. found that increased soil AP levels reduced AMF genus in cultivated paddy fields[81]. Luo et al. similarly demonstrated a negative effect of soil AP on AMF Chao1 richness in the topsoil of maize and wheat[35]. Chen et al. further indicated that increased P input also suppressed root growth, mycorrhizal infection, and phosphatase activity, ultimately resulting in decreased P uptake and productivity[82]. Consistent with these findings, our results revealed a negative correlation between soil AP and AK with AMF diversity (Fig. 6), suggesting that plants are highly dependent on AMF for P or K acquisition in low fertility soils[73], whereas low soil AP or AK levels may promote AMF diversity.
The GRSP dynamics provided further insights into soil-AMF interactions. The significantly higher levels of EE-GRSP in Gao County than in Ningnan County may reflect both enhanced AMF activity and a potential positive feedback mechanism in nutrient-limited soils. EE-GRSP, a glycoprotein secreted by AMF during symbiosis[44], not only serves as an indicator of AMF metabolic activity but also actively contributes to soil structure stabilization and nutrient retention[83]. We propose a positive feedback mechanism: under nutrient deficiency conditions, increased host plant reliance on AMF may stimulate higher EE-GRSP production, which in turn could improve soil microhabitat conditions and further facilitate AMF-plant nutrient exchange[84]. This self-reinforcing cycle maintains AMF diversity in low-P soils, whereas high P disrupts it by suppressing EE-GRSP synthesis and destabilizing hyphal networks[85]. Plant-specific effects were also evident in GRSP distribution patterns. Our findings indicated that the amount of EE-GRSP and T-GRSP varied across different mulberry species and decreased with soil depth in FengTian 5, HongYa, and YunSang 1 (Fig. 1a, b), suggesting that GRSP composition can vary among plant varieties. On one hand, distinct plant species establish unique mycorrhizal associations, which influence the soil's quantity and composition of Glomalin[86]. Some tree species foster the production of specific glomalin proteins, thereby affecting the functional diversity within the soil microbial community[87]. On the other hand, root exudate compositions are variable among different plant species, which significantly influence glomalin production as well as microbial community structure[88].
The carbon cycling dimension was particularly noteworthy. GRSP is a crucial component of SOC and serves as a significant source for soil C pools. Our study observed a strong positive correlation between T-GRSP and SOC, as well as a positive relationship between SOC and AMF richness (Fig. 6). This suggests that GRSP may contribute to soil C storage by stabilizing soil aggregates and protecting organic matter from decomposition[89], accounting for approximately 30%–60% of the carbon in undisturbed soils[44]. Furthermore, higher levels of SOC could support greater germination rates of AMF spores and mycelium in response to carbohydrate demands, thus directly influencing the composition of AMF communities within rhizosphere soils[90,91]. Consistent with our results, Luo et al. demonstrated that SOC shapes the composition of AMF communities in both soils and roots of Bermudagrass[69]. Zhu et al. assessed the diversity and composition of AMF communities across ten maize cropland soils within the black soil zone of northeast China using MiSeq sequencing techniques and revealed significant positive correlations between SOC content, total nitrogen levels, and OTU richness[92] In addition, geographic differences in baseline soil chemistry created distinct selective environments - Gao County's acidic soils (pH 4.9−5.5) promoted 37% higher EE-GRSP production than Ningnan's neutral soils, revealed pH-dependent glomalin stabilization (r = −0.72) as another regulatory layer. Thus, soil condition could be associated with a given habitat that influences the soil community structure of AMF. Therefore, these findings challenge conventional single-factor approaches in tree crop systems. The emergent significance of soil parameters only after accounting for varietal effects suggests that edaphic influences are typically underestimated. This has critical implications for experimental design in agroecological studies, emphasizing the need for multifactorial frameworks that integrate soil chemistry, microbial dynamics, and host plant genetics.
-
The study demonstrates that all tested mulberry varieties form beneficial symbiotic relationships with AMF, with 'Chuanshang' showing the highest colonization rate. Glomus and Paraglomus dominated the root-associated AMF communities. AMF community composition varied significantly among varieties and was jointly shaped by host genotype and soil factors. Mulberry varieties initially drive AMF recruitment, while soil properties (notably AP and AK) further refine community composition through environmental filtering. Critically, AP and AK negatively impacted AMF diversity, whereas SOC exhibited a notable direct positive effect on soil AMF abundance, revealing a nutrient-dependent trade-off in microbial balance. These findings highlight the need for tailored soil management—reducing excessive phosphorus/potassium inputs while increasing organic carbon—to optimize AMF-plant interactions, thereby improving soil health, increasing orchard resilience, and ensuring the long-term sustainability of mulberry orchards.
This study was jointly supported by National Youth Natural Science Foundation of China (31800612), National Natural Science Foundation of China (42267056), and Major Science and Technology Special Projects in Yunnan Province (202402AE090012), and the Science and Technology Department of Sichuan Province (2018JZ0027).
-
The authors confirm contribution to the paper as follows: conceptualization, project administration: Shi S, Wen M; data curation: Shi S, Huang L, Luo X; formal analysis: Shi S, Wang X; investigation: Shi S, Huang L, Wang X; methodology: Shi S, Huang L, Wang X, Luo X; writing – original draft: Shi S; writing – review & editing: Shi S, Wen M, Xu X, He X; software: Huang L, Wang X, Luo X; funding acquisition: Wen M; supervision: Wen M, Xu X, He X; validation: Xu X. All authors reviewed the results and approved the final version of the manuscript.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Composition of virtual taxa species in the total soil samples collected from different mulberry orchards.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Yunnan Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shi S, Huang L, Wang X, Wen M, Luo X, et al. 2025. Variation in arbuscular mycorrhizal fungal communities across mulberry varieties and soil properties in orchard ecosystems. Agrobiodiversity 2(2): 44−55 doi: 10.48130/abd-0025-0006 

Variation in arbuscular mycorrhizal fungal communities across mulberry varieties and soil properties in orchard ecosystems
- Received: 16 January 2025
- Revised: 16 May 2025
- Accepted: 28 May 2025
- Published online: 27 June 2025
Abstract: Mulberry (Morus alba L.), a prominent cash crop tree in China, depends on arbuscular mycorrhizal fungi (AMF) for soil nutrient acquisition. However, the composition and drivers of AMF communities in mulberry orchards remain poorly understood. Here, we characterized the AMF communities across six mulberry varieties using MiSeq sequencing of orchard soils in Sichuan Province. Our results revealed that all varieties formed symbiotic associations with AMF, with 'Chuanshang' showing the highest colonization rate. We identified 1334 operational taxonomic units (OTUs), predominantly from Glomus (61%) and Paraglomus (35%). AMF communities varied significantly across sampling sites, influenced by both mulberry varieties and soil properties. While mulberry varieties dominate initial AMF recruitment through genotype-specific selection, soil properties - particularly phosphorus availability (AP), potassium supply (AK), and soil organic carbon (SOC) subsequently shape community composition through resource-mediated environmental filtering. Variance partitioning indicated that 9.6% of community variation was jointly explained by host varieties and soil factors. Notably, AP and AK had direct negative impacts on AMF diversity, whereas SOC had a notable direct positive effect on soil AMF abundance. This study demonstrates that mulberry varieties and soil properties jointly govern AMF community assembly, with host selection and soil resource availability acting as sequential filters. Our findings highlight SOC as a key enhancer of AMF abundance, while high phosphorus and potassium levels may reduce AMF diversity. These insights provide a basis for optimizing AMF-mediated nutrient management in mulberry orchards, suggesting that maintaining organic carbon and balancing fertilizer inputs could enhance symbiotic efficiency and ecosystem sustainability.