








-
Water lilies are perennial aquatic angiosperms widely recognized for their significant ornamental value due to their distinctive floral morphology and vibrant coloration. These plants possess exceptional ornamental attributes, characterized by diverse floral coloration, pleasant fragrance, and unique floating leaf structures, rendering them indispensable visual focal points in global water horticulture and landscape design. Beyond their aesthetic merit, water lilies demonstrate robust environmental adaptability and broad ecological tolerance. Consequently, they facilitate water purification and ecological restoration while simultaneously enhancing aquatic landscapes. It is this synergy between 'ornamental aesthetics' and 'ecological functionality' that has established the dominant status of water lilies in landscape applications in China and globally, underscoring their significant economic and cultural value[1]. Water lilies are native to northern and eastern Europe, northern Asia to Japan, and North America. There are approximately 50 species worldwide, of which five species are native to China[2]. Currently, there is no comprehensive global report on the chemical composition of water lilies[3]. This review provides a detailed overview of the research progress on the chemical constituents and biological activities of water lilies, with the aim of offering a scientific reference for their further development, utilization, and value-added applications.
-
The order Nymphaeales is subdivided into three distinct families: Nymphaeaceae, Cabombaceae, and Hydatellaceae. Among these, Nymphaeaceae comprises diverse genera including Euryale, Victoria, Barclaya, Nuphar, and Nymphaea, while Cabombaceae contains Brasenia and Cabomba, and Hydatellaceae consists of the single genus Hydatella. The genus Nymphaea itself is further categorized into five subgenera: Brachyceras, Lotos, Nymphaea, Anecphya, and Hydrocallis. In terms of horticultural classification, Nymphaea species are generally classified into two horticultural groups: tropical and hardy, based on their growth temperature requirements[4]. The hardy water lilies, belonging primarily to the subgenus Nymphaea, are capable of overwintering outdoors in temperate zones. In contrast, the tropical group, which encompasses the subgenera Brachyceras, Anecphya, Lotos, and Hydrocallis, is sensitive to low temperatures and cannot survive outdoors during temperate winters[5].
Biological characteristics of Nymphaeales
Biological characteristics of Nymphaea plants
-
Nymphaea species are perennial aquatic flowering plants whose biological characteristics, including phenology, growth habits, and flowering behaviors, vary significantly in response to environmental conditions. Morphologically, they possess fibrous roots and thickened rhizomes, which may be tuberous or subterranean and exhibit diverse forms. The floating leaves typically feature a basal notch and diverse shapes (circular, elliptical, or ovate), supported by cylindrical petioles that elongate in response to water depth. Leaf coloration is highly variable, ranging from dark green to reddish-brown, a trait linked to genetic adaptations such as specialized stomata that facilitate aquatic survival[6]. Florally, Nymphaea is renowned for its wide spectrum of colors and petal shapes. Scent profiles also vary by climatic adaptation: cold-tolerant varieties tend to be faintly fragrant, whereas heat-tolerant ones are intensely scented. Nymphaea species exhibit distinct flowering rhythms, classified as either diurnal or nocturnal; however, the precise timing of anthesis is modulated by temperature fluctuations. Physiologically, these opening and closing movements are mediated by epidermal cells located at the base of the petals. Following pollination, the fruit develops as a berry submerged in water, containing numerous seeds that vary significantly in morphology. Generally, cold-tolerant varieties produce larger seeds with a smooth coat, whereas heat-tolerant varieties are characterized by smaller seeds with a hairy coat, displaying further diversity in color and distribution patterns[7]. In addition to sexual reproduction through seeds, Nymphaea species are also capable of vegetative propagation via rhizome division and vivipary.
Beyond Nymphaea, the family Nymphaeaceae encompasses other genera with distinct biological traits. The genus Nuphar, comprising approximately 25 species distributed across the Northern Hemisphere (Fig. 1), is characterized by floating leaves with long petioles and thick, glossy, ovate blades, accompanied by relatively small yellow flowers[8]. In contrast, the genus Euryale, predominantly distributed in China (ranging from Heilongjiang to Guangdong), is noted for its distinct dimorphic foliage: submerged leaves are spineless and arrow-shaped, while the large (10–130 cm), leathery floating leaves are heavily armed with sharp spines. Euryale is valued not only for its starch-rich, edible seeds, which possess traditional medicinal properties for tonifying the spleen and kidney, but also for its utility as fodder and green manure[9].
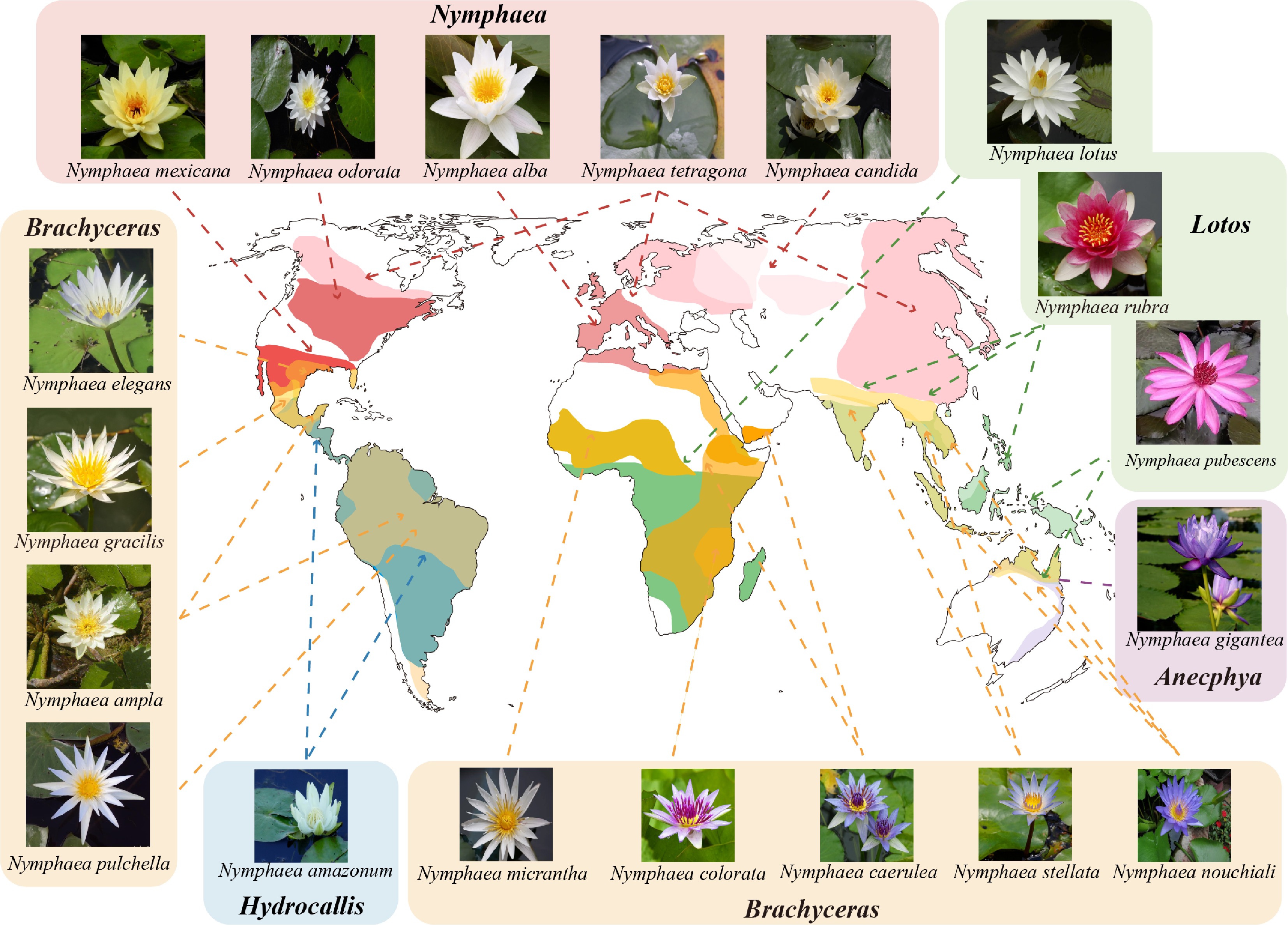
Figure 1.
Global geographical distribution of representative Nymphaea species. The map illustrates the distinct distribution patterns of five Nymphaea subgenera. The subgenus Nymphaea (pink) is predominantly distributed in the northern temperate regions (e.g., N. alba, N. odorata). Tropical and subtropical regions are inhabited by the other four subgenera: subgenus Lotos (green) in Africa and Asia (e.g., N. lotus, N. rubra); subgenus Anecphya (purple) endemic to Australia (e.g., N. gigantea); subgenus Hydrocallis (blue) distributed in the Neotropics (e.g., N. amazonum); and the widely distributed subgenus Brachyceras (yellow/orange), which spans across the Americas, Africa, and Asia (e.g., N. caerulea, N. nouchiali, N. colorata). Representative flower phenotypes for each subgenus are shown in the panels.
The genus Victoria, native to the Amazon River Basin, represents the pinnacle of aquatic gigantism. It's two species, V. amazonica and V. cruziana, possess the world's largest aquatic leaves, exceeding 3 m in diameter. Their smooth surfaces and vertically upturned leaf margins form a unique peltate structure, conferring exceptional ornamental value[10].
Biological characteristics of Cabombaceae
-
The genus Brasenia, a member of the family Cabombaceae, is represented by Brasenia schreberi J.F. Gmel., a perennial aquatic herb. The new shoots and curled young leaves, which are encased in a translucent gelatinous sheath, are not only edible but also possess high nutritional value[11]. Furthermore, the species exhibits significant medicinal properties. The mucilage coating the plant surface is identified as an acidic heteropolysaccharide that demonstrates antioxidant, anti-inflammatory, hypoglycemic, and hypolipidemic activities[12].
In contrast, plants of the genus Cabomba, also belonging to the Cabombaceae, are perennial aquatic herbs native to the eastern regions of South America. Characterized by a well-developed, slender, and branched root system, these plants exhibit distinct leaf phyllotaxy; specifically, the leaves are arranged in opposite or whorled patterns, with submerged leaves being opposite and floating leaves alternate. Predominantly inhabiting riverbanks, lakes, and canals in plain regions, these species are characterized by rapid growth and a tolerance for varying water quality. They possess a robust reproductive capacity, propagating primarily through asexual fragmentation, where individual nodes are capable of regenerating into complete plants under favorable conditions[13].
Biological characteristics of Hydatellaceae
-
The family Hydatellaceae exhibits a geographic distribution spanning the coastal regions of Australia, New Zealand, Tasmania, and eastern India, showing differentiation across tropical, subtropical, and temperate climatic zones. Ecologically adapted to both ephemeral and permanent aquatic environments, these plants are predominantly annuals, although the family includes a few perennial species capable of asexual reproduction. Reproductive morphology is characterized by minute flowers that are either unisexual or bisexual, supporting both monoecious and dioecious breeding systems[14]. To date, no secondary metabolites have been reported for the Hydatellaceae, a fact likely attributable to the limited species richness and restricted geographical range of this family.
-
Although early research was limited, recent metabolomic studies have revealed a rich diversity of compounds in Nymphaea species. Nymphaea plants are rich in flavonoids, phenolic acids, volatile oils, and other compounds, which exhibit a wide range of biological activities.
Chemical constituents in Nymphaea
-
The genus Nymphaea is characterized by a diverse phytochemical profile. As early as 2014, Zhao et al. cataloged 74 flavonoids and phenolic acids, alongside 13 other compounds isolated from these species[15]. However, given the significant advancements in phytochemical research since that time, this review aims to provide a comprehensive update on the genus, synthesizing key findings from the literature accumulated in recent years.
Flavonoids and phenolic acids in Nymphaea species
-
Nymphaea species are characterized by an abundance of secondary metabolites, particularly flavonoids and phenolic acids, which confer significant bioactivities ranging from antioxidant, antiviral, and antitumor effects to the modulation of glycolipid metabolism[15,16]. With respect to flavonoids, investigations have demonstrated a structural diversity dependent on the specific cultivar. Notably, myricetin glycoside derivatives have been isolated from N. lotus[17], whereas N. candida, N. stellata, and hybrid cultivars are predominantly enriched with kaempferol, quercetin, and their glycoside derivatives[18,19]. Studies focusing on floral color variation have further elucidated the distribution profiles of anthocyanins, chalcones, and flavonols within tropical water lilies[20]. Furthermore, structurally unique flavonoids have been documented, such as the quercetin xyloside derivatives exhibiting PDE5 inhibitory activity isolated from N. pubescens[21].
In terms of phenolic acids and polyphenols, gallic acid, caffeic acid, ferulic acid, and their esters are widely distributed across various species, such as N. candida, N. nouchali, and N. pubescens[22−24]. Beyond simple phenolic acids, hydrolyzable tannins (e.g., ellagic acid, geraniin, and pentagalloyl glucose) are also important active constituents of N. candida and N. alba[25,26].
In recent years, advancements in isolation techniques and metabolomics have led to a deeper understanding of the chemical composition of Nymphaea. Zhao et al. systematically summarized 87 compounds in this genus, encompassing categories such as lignans and alkaloids[15]. Advanced extraction and detection technologies, such as ultrasound-assisted extraction, high-speed counter-current chromatography (HSCCC), and high-resolution mass spectrometry (HRMS), have been applied for the rapid identification of complex components. For example, Naznin et al. identified 150 secondary metabolites, including novel flavonoids, in N. rubra[27]. Most recently, a novel resorcinol derivative was discovered in N. lotus, further enriching the known chemical diversity of the genus Nymphaea[28].
Other compounds in Nymphaea species
-
In addition to flavonoids and phenolic acids, Nymphaea species have been shown to produce a diverse array of other secondary metabolites. Zhang et al. isolated lignan compounds from N. odorata, including nymphaeoside A[29]. Metabolomic profiling of the floral exudate of N. 'Blue Bird' further revealed that its major constituents comprise D-gluconic acid, coumarins, glucuronic acid, and several flavonoids, highlighting its distinctive antioxidant potential[30].
Research on floral scent and volatile constituents in Nymphaea has been relatively extensive. Collectively, previous studies indicate that benzyl alcohol, benzaldehyde, and their esters, together with long-chain alkanes (e.g., pentadecane), constitute the principal structural components of the essential oils of hybrid water lilies[31−33]. Yu et al. reviewed 36 volatile compounds identified in N. hybrid, with particular emphasis on the structural diversity of terpenoids[19]. With the application of headspace solid-phase microextraction (HS-SPME) coupled with GC–MS, floral scent characteristics can now be captured with greater resolution. A comparative analysis of 62 Nymphaea taxa by Su et al. identified 72 volatile compounds, predominantly benzyl benzoate, 4-methoxybenzaldehyde, and various monoterpenes (such as ocimene and terpinene)[34]. More recently, investigations of the cultivar 'Eldorado' across different developmental stages have further elucidated dynamic changes in its volatile profile, revealing numerous monoterpenes and their oxygenated derivatives, including α-pinene, D-limonene, and terpinen-4-ol[35].
Compounds from the genus Brasenia
-
Brasenia schreberi are rich in bioactive polysaccharides, proteins, amino acids, and trace elements, and therefore exhibit considerable medicinal and nutritional value[12]. With respect to flavonoids and phenolic acids, a series of compounds have been isolated and identified from their extracts, including quercetin, kaempferol, isorhamnetin, and their glycosylated derivatives, as well as commonly occurring constituents such as gallic acid and luteolin. Among these, isorhamnetin and quercetin are regarded as the predominant components[36,37]. In addition, several flavonoids with unique structural features have been successively reported, exemplified by the novel flavonol glycoside Junsainoside A[38].
In recent years, investigations focusing on materials from different geographical origins and employing advanced extraction strategies have further expanded the chemical profile of B. schreberi. Vahekeni et al. isolated multiple compounds from Angolan populations of B. schreberi that were reported for the first time in this genus, including gossypetin glycosides and various polygalloyl glucoses (hydrolyzable tannins)[28]. Using an ultrasound-assisted aqueous two-phase extraction approach, Xu et al. successfully isolated a novel polysaccharide (BPS-1) and, for the first time in B. schreberi, identified schaftoside and its isomers, quercetin glucoside glucuronides, and phenolic constituents such as 3',4',7,8-tetrahydroxyflavanone[39].
Compounds from the genus Nuphar
-
Plants of the genus Nuphar are well recognized for their richness in alkaloids. Zhou et al. systematically summarized 57 alkaloids reported from this genus, which predominantly fall into two structural classes characterized by C15 and C30 skeletons[40]. Building upon this foundation, subsequent studies have further expanded the chemical diversity of Nuphar. Investigations of N. japonicum and N. pumilum have led to the isolation of a range of characteristic alkaloids, including deoxynupharidine, nuphamine, thiobinupharidine, and their respective isomers[8,41]. In addition to alkaloids, essential oils constitute another important chemical fraction of Nuphar species. Peng et al. identified 29 constituents in the essential oil of N. pumilum, which, besides alkaloids, also contained compounds such as hexadecanoic acid and muscopyridine[42].
In recent years, the application of advanced separation techniques and high-sensitivity analytical methods has facilitated the discovery of numerous structurally novel compounds. Orejola first isolated 1,4-dehydrohexahydroxydiphenoyl-α-D-glucopyranose, a compound featuring a complex tannin structure, from N. japonicum[43]. Using carboxyl-functionalized polymeric magnetic microspheres in combination with LC–MS, Lin et al. achieved the selective extraction and identification of bioactive constituents from N. pumila, leading to the characterization of 15 compounds, including seven previously unreported alkaloids: cyanopumiline A sulfoxide, cyanopumiline C sulfoxide, cyanopumiline A, cyanopumiline B, cyanopumiline C, cyanopumiline D, and cyanopumiline E[44].
-
The ornamental value of Nymphaea is intimately associated with specific secondary metabolites that determine their olfactory and visual phenotypes. Floral scent, a key trait for pollinator attraction and horticultural appeal, is highly correlated with the composition of volatile organic compounds (VOCs) (Fig. 2). A comprehensive GC-MS analysis of 62 Nymphaea cultivars revealed that the diversity of scent profiles, ranging from herbal to sweet, depends primarily on the ratios of alkenes, alkanes, and alcohols. Among these, benzyl acetate and cis-ocimene were identified as the major contributors determining scent intensity[34]. Genomic analysis of Nymphaea colorata reveals that the biosynthesis of these attractive volatiles involves the expansion of the TPS-b subfamily and SABATH methyltransferases, which catalyze the production of sesquiterpenes and fatty acid derivatives, respectively[45]. Understanding these metabolic signatures holds significant application potential: it enables breeders to utilize volatile 'fingerprints' for precise cultivar identification and to screen parental lines with high ester or terpene content for breeding novel aromatic varieties or developing essential oil products[34,46].
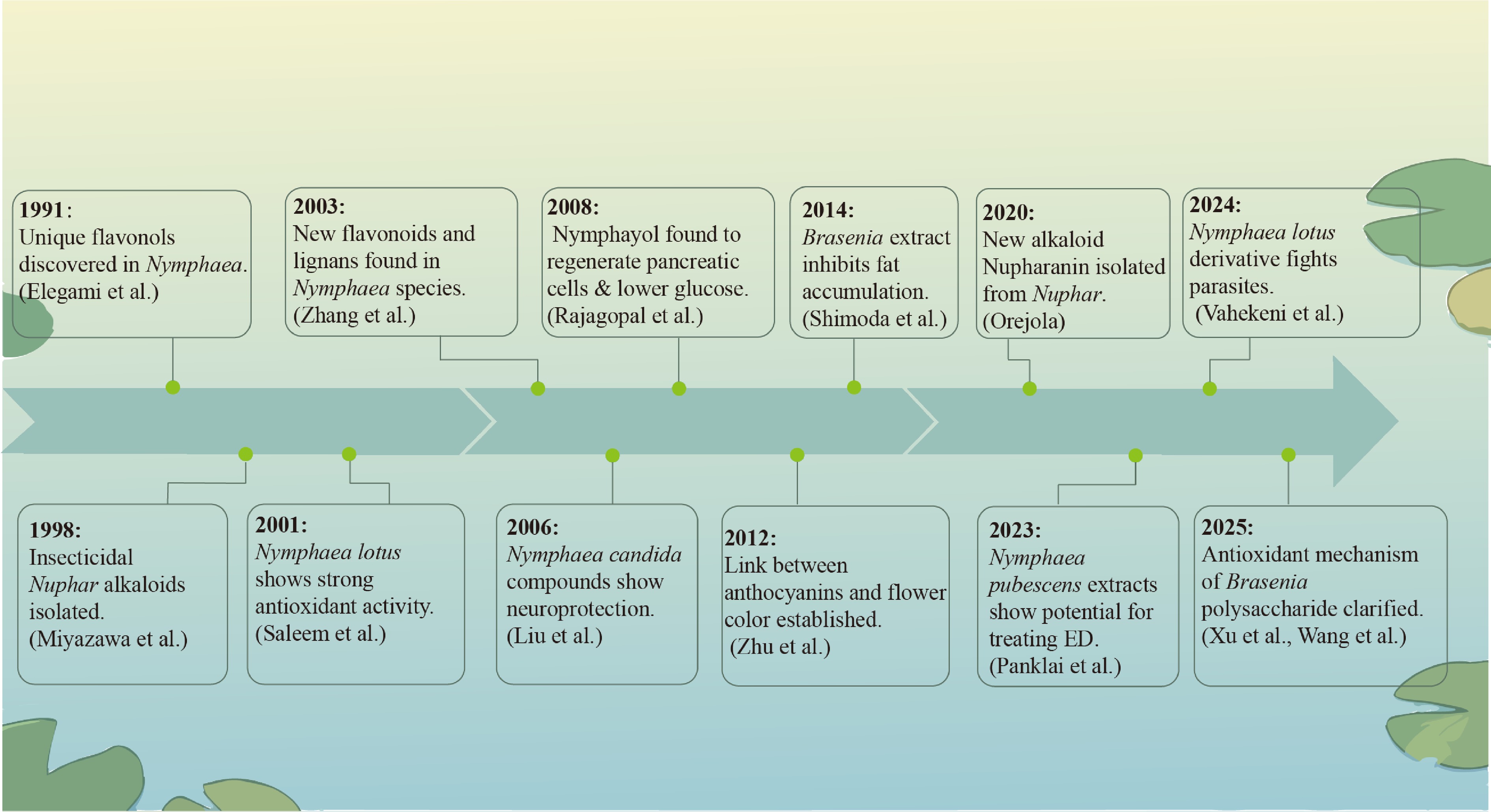
Figure 2.
Historical development of metabolites.
Pigment diversity and flower coloration
-
Petal coloration in Nymphaea is directly controlled by the accumulation and specific modification of anthocyanins, providing a chemical basis for flower color breeding. In hardy water lilies, a study of 57 cultivars demonstrated a significant linear correlation between anthocyanin composition and colorimetric parameters (L*, a*, b*). Specifically, the accumulation of delphinidin-3-O-β-galactopyranoside, delphinidin-3-O-(2''-O-galloyl-β-galactopyranoside), and cyanidin-3-O-galactosyl-(1→2)-galactoside was found to be positively correlated with redness values (a*) and negatively correlated with lightness values (L*), indicating their role in deepening the red hue of petals[2]. This establishes these specific anthocyanins as metabolic markers for screening deep red varieties. Furthermore, comparative genomic analyses highlight that the blue phenotype in tropical water lilies (e.g., N. colorata) is determined by specific delphinidin derivatives, whose biosynthetic pathway requires the synergistic high expression of anthocyanin synthase (ANS) and UDP-glucose : flavonoid glucosyltransferase (UDPGT)[45]. This pathway provides a roadmap for molecular breeding: introducing or upregulating these key genes through genetic engineering or distant hybridization holds immense potential for breaking color barriers and cultivating the highly anticipated blue hardy water lilies[2,45].
Volatile compounds and scent profiles
-
The olfactory profile of Nymphaea is characterized by a highly diverse array of volatile organic compounds (VOCs), which vary significantly among species and cultivars. A systematic GC-MS analysis of 62 Nymphaea cultivars identified 72 distinct volatile constituents, composed primarily of alkenes, alkanes, and alcohols. Key aroma-active compounds included benzyl acetate (jasmine-like), cis-ocimene (mint/herbal-like), benzyl alcohol (fresh/faint scent), and farnesene (fruity). Significant differences in the composition and abundance of volatiles were observed between tropical and hardy water lilies. The study found that tropical water lilies contained a much higher number of detected volatile components (56) compared to hardy water lilies (37), rendering the scent of tropical varieties generally more complex and intense[34]. Genome analysis of N. colorata revealed that the expansion of the TPS-b subfamily (regulating monoterpene/sesquiterpene synthesis) and the SABATH family (involved in ester metabolism) may be associated with the evolution of floral scent. This chemical diversity classifies Nymphaea scents into categories ranging from herbal to fruity and sweet, providing rich genetic resources for breeding aromatic varieties. Spatiotemporal variations are also closely linked to Nymphaea scent, influencing pollination efficiency and harvest value. Research on the prized cultivar Nymphaea 'Eldorado' indicated that volatile emission is dynamic, varying significantly with flowering stages and floral parts[35]. Similarly, in the night-blooming Victoria cruziana, floral scent is dominated by methyl hexanoate. The biosynthesis of this compound is rhythmically regulated, peaking during the flower's secondary opening phase to attract beetle pollinators[47]. Understanding these temporal emission patterns is critical for the horticultural industry, particularly for determining optimal harvest times to maximize scent retention for cut flowers or essential oil extraction.
The unique volatile characteristics of water lilies offer broad prospects for product development in the cosmetics and fragrance additive industries. For instance, varieties rich in sesquiterpenes and benzyl alcohol have garnered attention for their potential in developing high-quality essential oils[46]. Moreover, unique volatile profiles can serve as reliable chemotaxonomic markers, enabling precise cultivar differentiation based on chemical composition. Utilizing these metabolic fingerprints, breeders can scientifically select parental lines with complementary scent traits to cultivate novel varieties with enhanced aromatic complexity and functional value[34,46].
-
The order Nymphaeales boasts a long-standing history of application within traditional medicinal systems globally. In China, particularly in traditional Uyghur medicine, Nymphaea candida is valorized as a critical medicinal resource. It constitutes a pivotal ingredient in prominent compound formulations such as 'Zukamu Granules' and 'Yanxiaodinaer Syrup', which are extensively prescribed for respiratory infections and inflammatory disorders[48,49]. In Traditional Chinese Medicine, the seeds of Euryale ferox, revered as 'aquatic ginseng', are celebrated for their spleen- and kidney-tonifying properties[9]. Moreover, Nuphar rhizomes are traditionally employed in folk remedies for rheumatoid arthritis and traumatic injury. Recent studies have correlated these ethnomedicinal uses with the potent immunosuppressive activity of thioalkaloids inherent to Nuphar, thereby substantiating the pharmacological rationale for their therapeutic potential[40,44].
Dietary and nutritional value
-
Beyond their medicinal utility, Nymphaeales species represent important sources of aquatic vegetables and functional foods. Brasenia schreberi is distinguished by its tender shoots and young foliage, enveloped in a characteristic transparent mucilage. Celebrated for its unique mouthfeel, it is processed into beverages, soups, and pastries, and is chemically characterized by high nutritional value, including abundant proteins, polysaccharides, and trace elements[11,50]. Euryale ferox, a starch-rich crop, yields seeds that are utilized not only for human consumption but also as livestock feed and green manure[9].
Industrial applications
-
Driven by the burgeoning concept of 'natural skincare', Nymphaea extracts are garnering increasing attention in the cosmetics and personal care sectors due to their exceptional bioactivities. Research has demonstrated that the polyphenols and flavonoids enriched in these extracts exert significant antioxidant and enzyme-inhibitory effects, positioning them as optimal natural additives for whitening serums, moisturizers, and facial masks[51]. Additionally, the distinctive olfactory signatures of water lilies underpin their utility in the fragrance industry (Fig. 3). Cultivars characterized by high concentrations of benzyl alcohol or sesquiterpenes can be selectively exploited for the extraction of high-value essential oils, which find extensive application in perfumery, soaps, and aromatherapy[34,46] (Fig. 4).
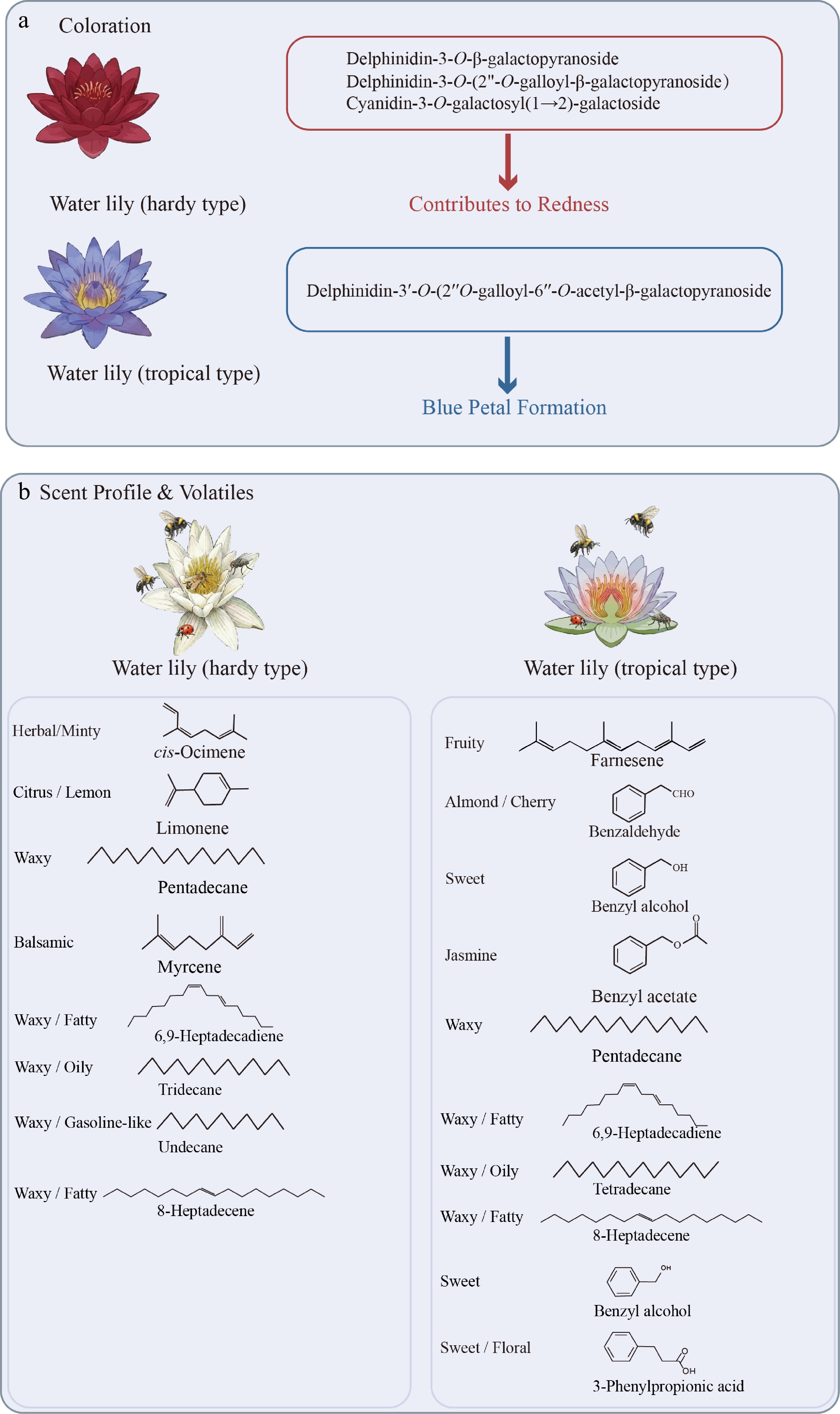
Figure 3.
Metabolic basis of coloration and scent divergence in Nymphaea. (a) Differential accumulation of anthocyanins determines petal color. Redness in hardy types is decided by non-acetylated delphinidin and cyanidin glycosides. Blue formation in tropical types relies on complex modifications, yielding delphinidin-3'-O-(2'' O-galloyl-6''-O-acetyl-β-galactopyranoside). (b) Representative volatile organic compounds (VOCs) and their odor descriptors. The scent of hardy water lilies comprises monoterpenes (cis-ocimene, limonene, myrcene) and alkanes/alkenes (e.g., undecane, tridecane). Tropical water lilies produce a complex bouquet of benzenoids (benzaldehyde, benzyl alcohol, benzyl acetate, 3-phenylpropionic acid) and farnesene.
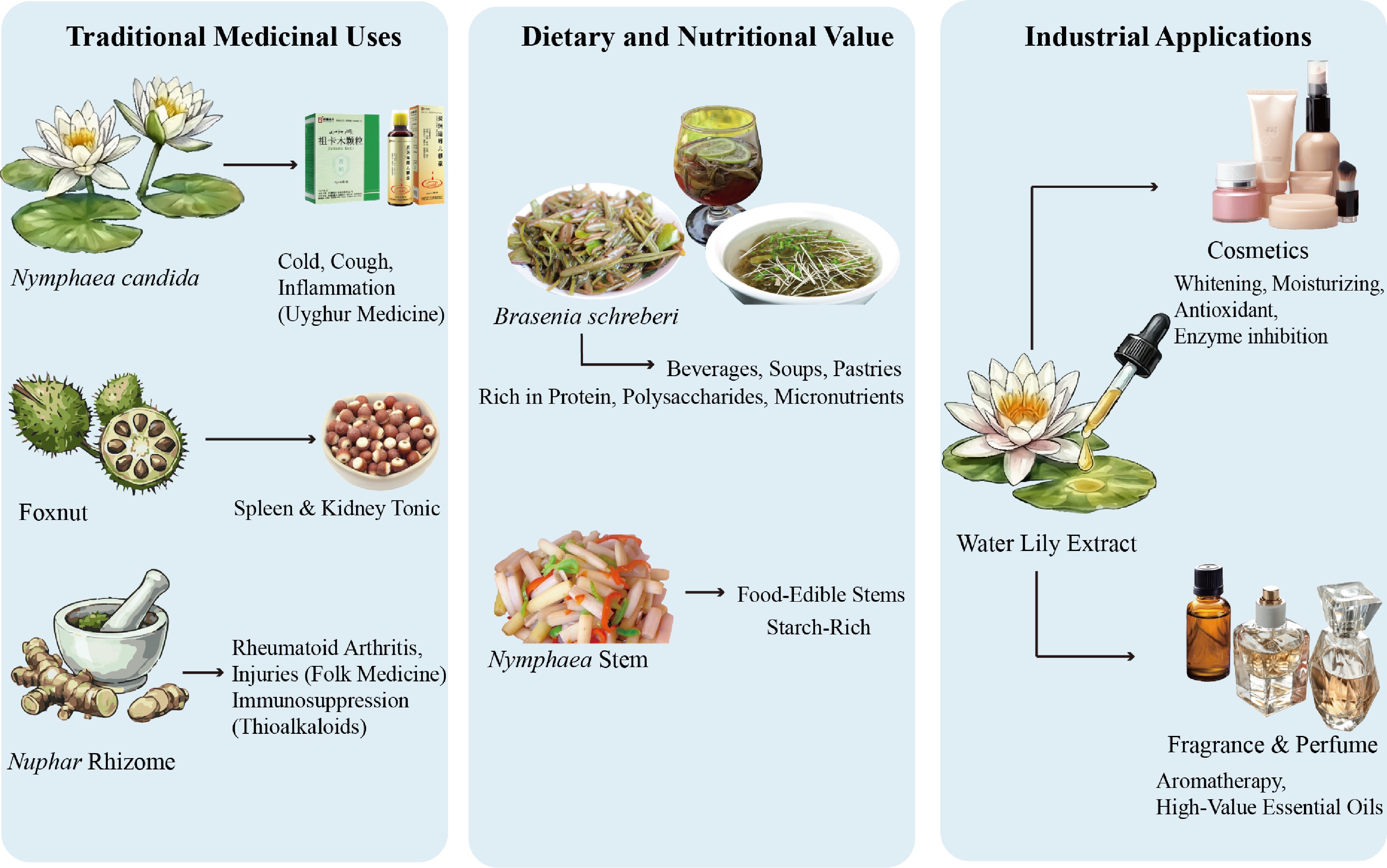
Figure 4.
Traditional uses, dietary value, and industrial applications.
-
Water lily plants contain numerous chemical constituents, such as polyphenols, flavonoids, polysaccharides, and amino acids. Previous studies have demonstrated that the chemical components in water lily flowers exhibit significant bioactivities, including antioxidant, hypoglycemic, tyrosinase inhibitory, and hepatoprotective effects, indicating their broad potential applications in functional foods, pharmaceuticals, and cosmetics[52] (Fig. 5). Water lily plants contain geraniin, a polyphenolic compound, which demonstrates multiple pharmacological activities such as antioxidant, radioprotective, antitumor, and antiviral effects[16] (Fig. 6).
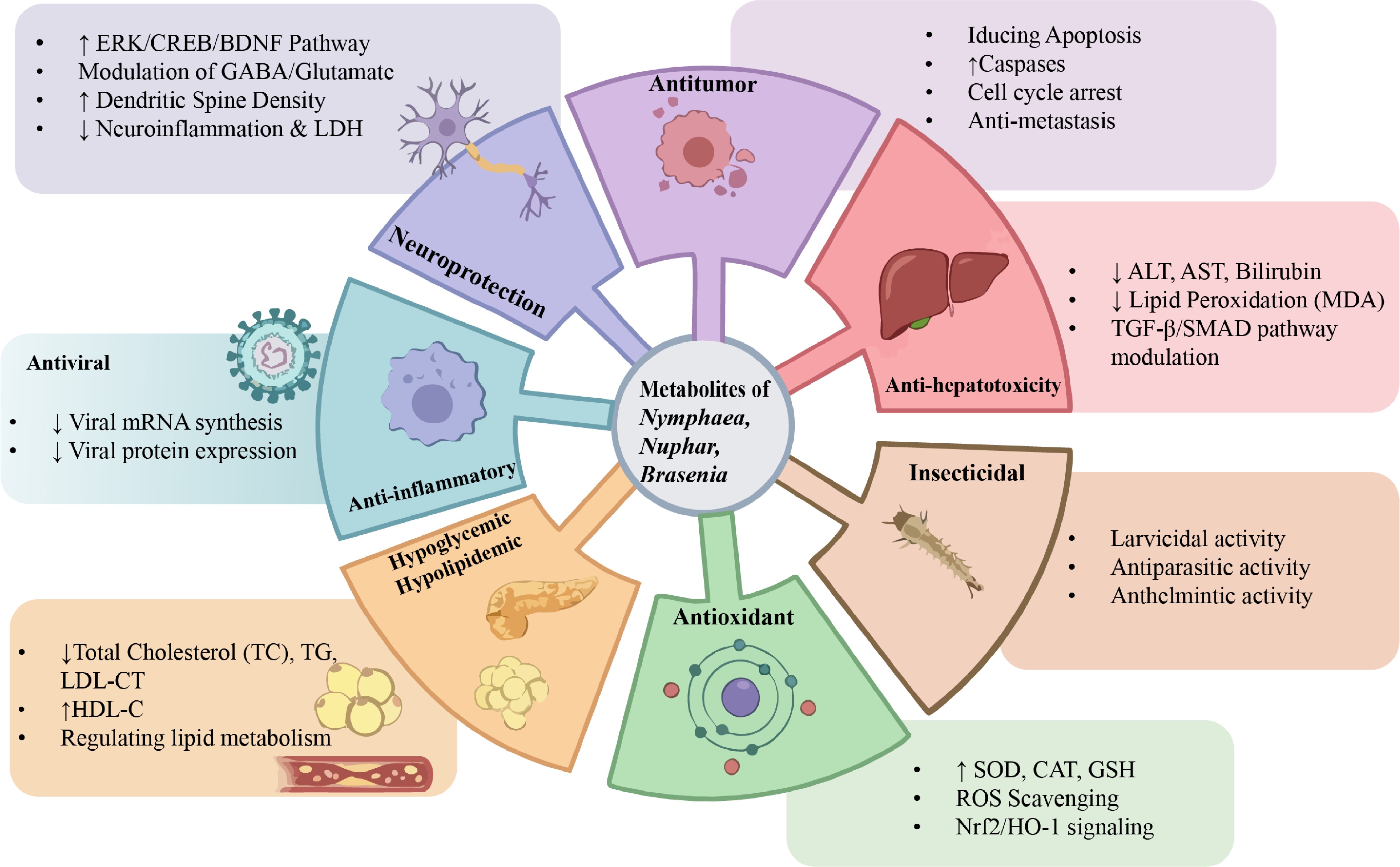
Figure 5.
Overview of the diverse bioactivities and underlying mechanisms of water lilies.
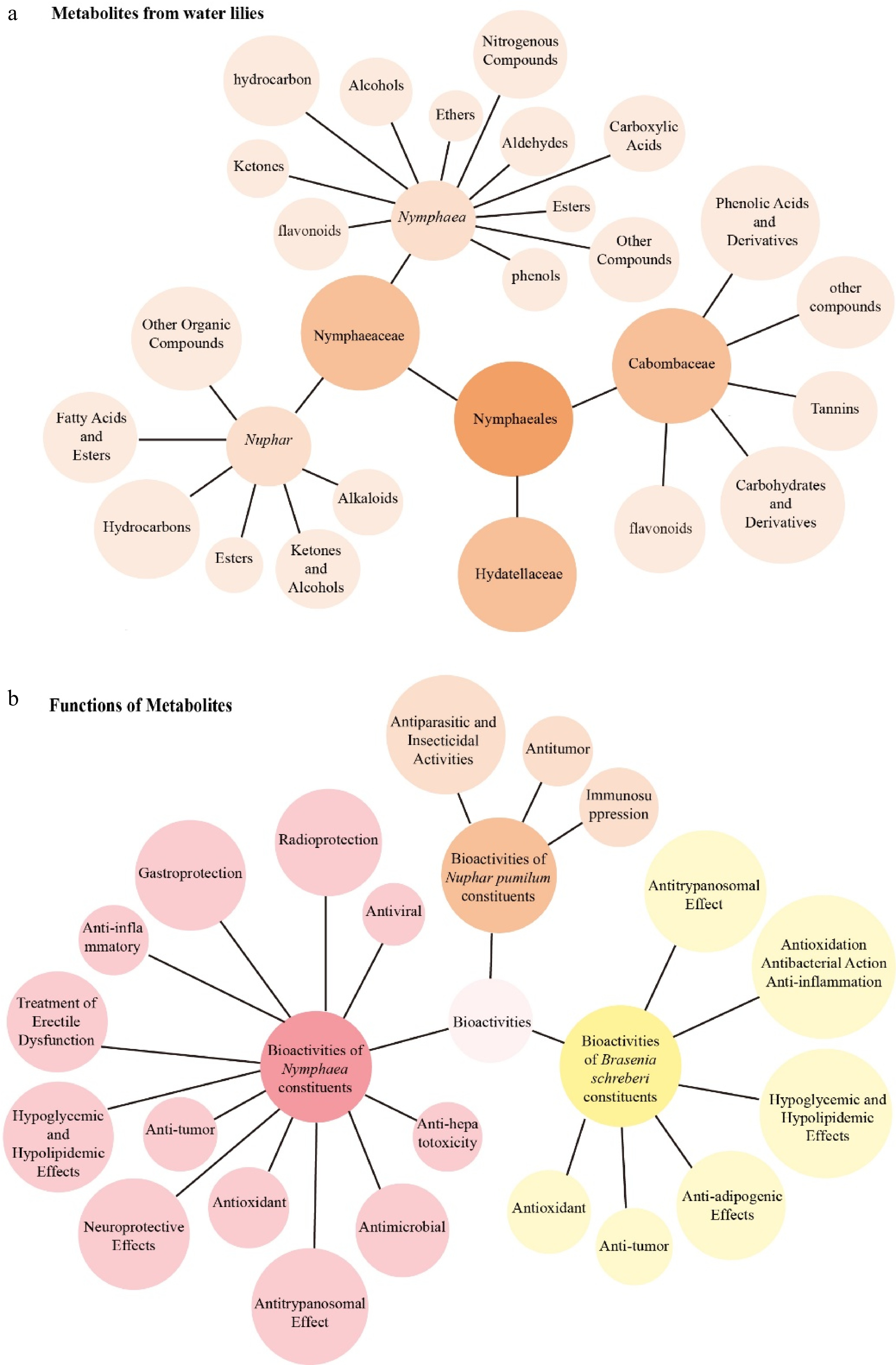
Figure 6.
Summarized chemical constituents and bioactivities of Nymphaeales species. (a) Distribution of the main classes of phytochemical constituents identified in Nymphaeales species. The figure shows the relationship between plant taxa (families Nymphaeaceae, Hydatellaceae, and Cabombaceae) and their major groups of compounds, including flavonoids, alkaloids, phenolic acids, tannins, terpenoids, fatty acids, and other organic compounds. (b) Summary of reported bioactivities of Nymphaeales constituents. The network highlights multiple pharmacological properties, including antioxidant, anti-inflammatory, antitumor, antimicrobial, hypoglycemic, and neuroprotective effects. Notably, compounds from Nymphaea exhibit broad bioactivities such as antioxidation, radioprotection, antiviral, and anti-trypanosomal activities.
Antiviral
-
Nymphaea candida serves as a pivotal ingredient in traditional Uyghur anti-inflammatory and antiviral polyherbal formulations, notably 'Zukamu Granules' and 'Yanxiaodinaer Syrup'[48]. While the specific bioactive constituents responsible for the therapeutic effects of Nymphaea in these formulas remain to be fully elucidated, investigations have corroborated that Zukamu granules achieve complete inhibition of Respiratory Syncytial Virus (RSV) replication at a non-toxic concentration (15.94 mg/mL)[49]. Beyond composite formulations, isolated extracts of the genus Nymphaea also exhibit pronounced antiviral potential. Specifically, extracts and purified fractions (notably fractions N1 and N8) of Nymphaea alba have been demonstrated to significantly downregulate HCV NS3/4A mRNA synthesis and NS3 protein expression, suggesting that they exert anti-HCV activity by disrupting the viral replication machinery[53].
Antitumor
-
The antitumor efficacy of Nymphaea alba has been extensively substantiated using both isolated compounds and crude extracts. Liu et al. characterized ten flavonol glycosides from the plant, among which kaempferol-3-O-α-L-rhamnopyranosyl glucopyranoside, kaempferol-3-O-(2"-O-galloylrutinoside), and myricetin displayed significant cytotoxicity against adrenal pheochromocytoma PC12 cells[18]. At the extract level, root and leaf extracts of N. alba exhibited substantial inhibitory effects on prostate (LNCaP), breast (MCF-7), and ovarian (A2780) cancer cell lines, while maintaining minimal toxicity towards normal cells (V79), thereby indicating a favorable safety profile and selectivity[54].
Mechanistic studies on Nymphaea stellata have provided insights into its anticancer pathways. The chloroform extract was found to induce apoptosis and arrest proliferation in human breast cancer MCF-7 cells by upregulating tumor suppressor genes (p53, Cdkn2a, pRb2), triggering the Caspase cascade, and modulating mitochondrial-dependent apoptotic signaling[55].
Moreover, Nymphaea lotus and Nymphaea caerulea demonstrate specific therapeutic potential against hematological malignancies. N'Guessan et al. reported that the N. lotus leaf extract possessed significantly more potent inhibitory activity against Jurkat lymphoma cells (IC50 = 87.29 μg/mL) compared to solid tumor MCF-7 cells, implying a cell-type specific sensitivity[56]. Recent findings further reveal that N. caerulea flower extract exerts targeted anti-leukemic activity by selectively inducing apoptosis in acute myeloid leukemia THP-1 cells via the modulation of cytokine expression and activation of the extrinsic apoptotic pathway[57].
Antioxidant
-
Polyphenols and flavonoids constitute the fundamental material basis underpinning the antioxidant activity of Nymphaea species. Early investigations identified a spectrum of monomers with potent free radical scavenging capacity in N. alba, N. caerulea, and N. candida. Key bioactive contributors were confirmed to include kaempferol derivatives (e.g., kaempferol-3-O-(2"-O-galloylrutinoside)), quercetin, myricetin, and their glycosides, along with gallic acid[8,18,22]. Beyond direct radical scavenging, these polyphenolic constituents confer profound cytoprotective effects. For instance, polyphenolic extracts of N. nouchali effectively scavenge ROS, prevent DNA damage, and attenuate oxidative stress-induced cell death by upregulating detoxification enzymes[23]; similarly, its rhizome extract (NNBR) safeguards erythrocytes against hemolysis and lipid peroxidation[58].
Notably, water lily extracts frequently exhibit dual functionality, combining antioxidant activity with tyrosinase inhibition, highlighting their potential for skin-whitening applications. The ethanol extract of Nymphaea has shown tyrosinase inhibition rates comparable to those of kojic acid[51]. Research on N. stellata and N. candida further corroborates that specific polyphenols (e.g., isostrictiniin) and flavonoids (e.g., kaempferol-3-O-rutinoside) act as the key agents responsible for this simultaneous antioxidant exertion and enzyme inhibition (targeting tyrosinase and maltase)[59].
In addition to typical phenolics, diverse plant parts and fractions also possess antioxidant properties. The volatile oil of N. stellata, crude polysaccharides of N. hybrid, and floral hydrosols of Nymphaea have all demonstrated significant efficacy in inhibiting oxidation and lipid peroxidation[30,60,61]. Gueye et al. further identified the seeds of N. lotus and N. micrantha as rich reservoirs of antioxidant compounds, including tannins and alkaloids[62].
Recently, research has advanced to cellular and in vivo models to unravel the molecular mechanisms governing Nymphaea antioxidant effects. Alam et al. demonstrated that N. nouchali extract mitigates oxidative stress-induced DNA damage by not only scavenging ROS but also upregulating detoxification enzymes[23]. In erythrocyte models, the rhizome extract effectively inhibited hemolysis and lipid peroxidation[58]. Furthermore, in vivo studies by Ishola et al. confirmed that N. lotus extract significantly enhances antioxidant enzyme activity and ameliorates lipid peroxidation levels in mice[63].
The robust antioxidant capacity of water lilies not only validates their pharmacological merit but also positions them as promising candidates for functional beverages and natural cosmetics. Given their potent radical scavenging and tyrosinase inhibitory activities, water lily flower extracts can be integrated into cosmetic formulations, such as serums, creams, and masks, to deliver whitening, anti-aging, and moisturizing benefits. Concurrently, flower teas derived from polyphenol-rich petals offer both aesthetic appeal and a daily source of antioxidants, satisfying the modern consumer's dual pursuit of beauty and wellness.
Anti-hepatotoxicity
-
Crude extracts of the genus Nymphaea have exhibited pronounced hepatoprotective efficacy across diverse models of chemical liver injury. Utilizing carbon tetrachloride (CCl4)-induced rat models, Bhandarkar et al. and Debnath et al. substantiated that extracts of Nymphaea stellata and Nymphaea pubescens effectively attenuated serum transaminase (AST, ALT) and bilirubin levels, while restoring the activity of hepatic antioxidant enzymes (SOD, GSH)[64,65]. In other chemical injury paradigms, Nymphaea hybrid extract was demonstrated to suppress alcohol-induced triglyceride (TG) accumulation in murine livers[66], whereas the ethanol extract of Nymphaea alba significantly ameliorated isoniazid (INH)-induced hepatotoxicity[67].
As investigations have deepened, specific bioactive constituents and their underlying mechanisms have been elucidated. Isostrictiniin and kaempferol-3-O-rutinoside, isolated from Nymphaea candida, were identified as pivotal hepatoprotective agents. Mechanistic studies revealed that isostrictiniin not only mitigated Concanavalin A (Con A)-induced acute hepatic injury but also improved hepatocellular necrosis and inflammatory infiltration[68]. Concurrently, kaempferol-3-O-rutinoside protected mice against D-galactosamine (D-Gal)-induced injury by boosting antioxidant enzyme activity and suppressing pro-inflammatory mediators[69].
Recently, research focus has expanded from acute injury to chronic liver pathologies. In models of chronic liver injury and fibrosis, Oyeyemi et al. observed that Nymphaea lotus extract not only inhibited oxidative stress (evidenced by reduced MDA and TBARS) but also impeded the expression of fibrosis-associated proteins by modulating the TGFβ/SMAD signaling pathway, thereby effectively attenuating the progression of CCl4-induced hepatic fibrosis[70,71].
Antibacterial and anti-inflammatory
-
The genus Nymphaea displays broad-spectrum antimicrobial properties. Investigations into the leaf and flower extracts of Nymphaea alba and the flower extract of Nymphaea nouchali have confirmed that their ethanolic fractions possess significant inhibitory or bactericidal activity against a range of pathogens, including Escherichia coli, Staphylococcus aureus, Bacillus subtilis, and Candida glabrata[54,72−74]. Beyond foliar and floral parts, seed extracts of Nymphaea nouchali were identified as rich sources of bioactive constituents, such as catechins, holding potential antibacterial value[75]. Notably, mechanistic insights have advanced from direct bactericidal effects to the modulation of bacterial virulence. For example, Nymphaea tetragona extract was shown to interfere with the bacterial Quorum Sensing (QS) system, significantly suppressing virulence factor expression in pathogenic bacteria[76]. Regarding in vivo efficacy, Batool et al. employed a murine listeriosis model to demonstrate that Nymphaea alba extract effectively restricted the colonization of Listeria monocytogenes across multiple organs, highlighting its prophylactic therapeutic potential[77].
In the context of anti-inflammatory activity, Nymphaea serves as a cornerstone in traditional anti-inflammatory formulations (e.g., Yanxiaodinaer Syrup), with its efficacy validated in multi-tiered cellular and animal models. Naringenin, isolated from Nymphaea mexicana, exhibited significant anti-inflammatory activity in RAW264.7 macrophages via a mechanism involving the suppression of ERK phosphorylation and NF-κB activation, subsequently downregulating iNOS, COX-2, and TNF-α expression[78]. Studies on Nymphaea hybrid revealed that its ethyl acetate fraction (NHE-EA) similarly curtailed the overexpression of pro-inflammatory cytokines through the dual blockade of NF-κB and MAPK pathways[79]. At the in vivo level, Nymphaea lotus leaf extract was proven to significantly attenuate carrageenan-induced paw edema in rats[80]. Moreover, Sun et al. elucidated that a polyphenol-enriched fraction (NCTP) exerted therapeutic effects in diverse inflammation models by reducing serum PGE2 levels and simultaneously inhibiting COX-2 and 5-LOX activities[48].
Hypoglycemic and hypolipidemic
-
Nymphaea species have a well-documented history in traditional diabetes management, and modern pharmacological research has substantiated their potent hypoglycemic and hypolipidemic activities. Multiple studies on Nymphaea stellata indicate that ethanolic or hydroalcoholic extracts of its leaves and flowers are efficacious in diabetic rat models. Rajagopal et al. further differentiated its action profile, noting that while the flower extract exerted no hypoglycemic effect on normoglycemic rats, it displayed significant antihyperglycemic activity in alloxan-induced diabetic rats[81]. Recently, Anand et al. reported that the powder of boiled Nymphaea nouchali rhizome (NNBR) was superior to the positive control Acarbose in mitigating sucrose-induced postprandial hyperglycemia[58].
Beyond the direct modulation of blood glucose and lipids, the inhibition of digestive enzymes represents a critical mechanism of metabolic regulation by Nymphaea. Regarding lipid metabolism, Zhang et al. first verified the hypolipidemic and hepatoprotective effects of Nymphaea hybrid in a quail hyperlipidemia model[82]. Subsequent in vitro screening revealed that various plant parts (whole flower, stamen, petal, and receptacle) possessed pancreatic lipase inhibitory activity, with the stamen extract exhibiting the most potent efficacy. This finding positions Nymphaea hybrid, particularly its stamens, as a promising source for developing natural lipid-lowering functional products[83].
Given their substantial inhibitory activities against α-glucosidase and pancreatic lipase, Nymphaea plants (especially stamens and rhizomes) hold immense potential for development as low-GI functional foods or dietary supplements. This insight informs breeding strategies to screen for varieties with high enzyme-inhibitory activity, facilitating the cultivation of dual-purpose (medicinal and ornamental) cultivars tailored to the metabolic health market.
Effect on neuronal cells
-
Flavonol glycosides within Nymphaea species have demonstrated remarkable potential in counteracting neuronal ischemic injury. Liu et al. substantiated that kaempferol-3-O-(2''-O-galloyl)-rutin glycoside, myricetin, and diverse kaempferol glycoside derivatives isolated from Nymphaea candida significantly attenuated cell death in rat primary cortical neurons and PC12 cells under conditions of oxygen-glucose deprivation (OGD) induced by glucose-free medium and sodium dithionite. These compounds concurrently reduced lactate dehydrogenase (LDH) release, exhibiting distinct neuroprotective properties[18,84].
Beyond direct cytoprotection, Nymphaea extracts are capable of ameliorating a spectrum of neurological dysfunctions. In the context of cognitive function, Nymphaea hybrid extract was shown to significantly reverse scopolamine-induced learning and memory deficits in mice. The underlying mechanism involves the mitigation of oxidative stress and modulation of cholinergic transmission, specifically via the activation of the ERK–CREB–BDNF signaling cascade in the hippocampus and cortex[85]. Regarding mood and behavioral regulation, the ethyl acetate fraction of Nymphaea candida displayed significant antidepressant activity, enhancing hippocampal dendritic spine density, promoting neural plasticity, and suppressing neuroinflammation[86]. Furthermore, Nymphaea lotus extract was confirmed to exert anticonvulsant effects by modulating key inhibitory (GABA) and excitatory (glutamate) neurotransmitters, while simultaneously alleviating epilepsy-associated cognitive and emotional comorbidities[63]. Notably, this species also regulates blood pressure and restores erectile function through synergistic central and peripheral mechanisms[87].
Moreover, the capacity of water lily extracts to enhance mood and cognition provides a scientific rationale for their application in aromatherapy. Aromatic cultivars enriched with neuroprotective volatile constituents can be selectively utilized for the extraction of high-value essential oils or integrated into stress-relieving therapeutic landscapes, thereby significantly expanding the industrial value chain of Nymphaea.
Others
-
The genus Nymphaea exhibits multifaceted potential in dermatology and anti-aging applications. Investigations indicate that Nymphaea nouchali extract significantly suppresses melanogenesis by modulating the MAPK/MITF/Tyrosinase signaling axis, offering therapeutic utility for hyperpigmentation[88]. Concurrently, Nymphaea rhizome extract protects keratinocytes against UVB-induced mitochondrial apoptosis and upregulates the expression of natural moisturizing factors (AQP3, FLG), thereby reinforcing skin barrier function and hydration[89,90]. In terms of systemic anti-aging, Nymphaea hybrid extract not only extends the lifespan and stress resistance of Caenorhabditis elegans[91], but also demonstrates significant 5α-reductase inhibitory activity, holding promise for the management of benign prostatic hyperplasia[92].
In the realm of metabolic disorders, Nymphaea rubra and its hybrids display potential in combating insulin resistance and obesity. Bioactive fractions have been shown to reverse TNF-α-induced insulin resistance in myocytes[93], exert hypoglycemic and hypolipidemic effects in STZ/high-fat diet-induced diabetic rats[33], and effectively prevent high-fat diet-induced obesity and hepatic oxidative stress in mice[94].
Regarding the digestive and urogenital systems, Nymphayol isolated from Nymphaea stellata exhibited significant anti-ulcer activity via antioxidant and anti-apoptotic pathways[95], while Nymphaea lotus was confirmed to possess potent anti-diarrheal properties[96]. Additionally, flavonoids in Nymphaea pubescens were characterized as potent PDE5 inhibitors, presenting a novel therapeutic avenue for erectile dysfunction[21].
In immunomodulation and antiparasitic research, Nymphaea rubra polysaccharides (NR-PS) have been shown to promote dendritic cell maturation and augment Th1 cytokine secretion[97], whereas the potent antioxidant Geraniin safeguards radiosensitive immune cells against DNA damage[98]. Notably, the genus also exhibits anthelmintic and antitrypanosomal activities; a recently identified resorcinol derivative demonstrated significant inhibitory efficacy against the pathogen responsible for sleeping sickness[28,99].
Bioactivities of chemical constituents from the genus Nuphar
-
Plants of the genus Nuphar are widely distributed, primarily across Europe, Asia, and the Americas. Their main bioactive constituents are sesquiterpene alkaloids, which exhibit diverse pharmacological activities, including inhibition of tumor metastasis, immunosuppression, anti-inflammatory effects, and antimicrobial properties.
Antitumor
-
Alkaloids derived from the genus Nuphar show significant promise as antitumor agents, with 6-hydroxythiobinupharidine characterized as a pivotal bioactive constituent. In vivo investigations demonstrated that oral administration of either this purified alkaloid or the alkaloidal fraction of Nuphar pumilum significantly suppressed pulmonary tumor formation in B16 melanoma-bearing mice, evidencing distinct anti-metastatic activity[100]. Subsequent mechanistic studies at the cellular level corroborated that this compound potently induces apoptosis in human leukemia U937 cells, thereby fulfilling a dual role in tumor suppression and metastasis inhibition[40].
Others
-
Nuphar pumilum is extensively utilized in traditional medicine for the management of rheumatoid arthritis and lumbocrural pain. Contemporary pharmacological evaluations have unveiled the potent immunomodulatory activities of its rhizome extracts and alkaloidal constituents, with (−)-deoxynupharidine standing out as a primary active agent. Recent investigations have further substantiated that Nuphar dimeric alkaloids (NDAs) isolated from the plant, comprising a suite of 12 compounds including cyanopumiline A and C sulfoxides, effectively abrogate anti-CD3/CD28-induced proliferation of human peripheral T cells. These results underscore the potent immunosuppressive function of these thioalkaloids, providing a pharmacological basis for the therapeutic application of this genus in autoimmune disorders[44].
In addition to immunomodulation, Nuphar species display broad-spectrum antipathogenic and insecticidal profiles. Nuphar lutea extract has been documented to exhibit activity against the measles virus[101]. Concurrently, a variety of alkaloids isolated from N. japonicum and N. lutea have demonstrated significant insecticidal and antiparasitic potential, notably exerting larvicidal effects against Drosophila melanogaster and inhibitory activity against Leishmania species[41,102]. Most notably, the novel dimeric sesquiterpene thioalkaloid, 6,6'-dihydroxythiobinupharidine, has been identified as a critical molecular entity responsible for antiparasitic activity[102].
Bioactivities of chemical constituents from the genus Brasenia
Hypoglycemic and hypolipidemic
-
Polysaccharides derived from Brasenia schreberi exhibit remarkable clinical and pharmacological efficacy in diabetes management. Early clinical trials established that the administration of a 3% Brasenia polysaccharide gel solution significantly attenuated blood glucose levels in diabetic patients[103]. Subsequent mechanistic investigations suggested that this hypoglycemic action is partially attributable to the inhibition of α-amylase and α-glucosidase, which retards carbohydrate digestion and absorption[104].
In animal models, Brasenia exopolysaccharides were shown to effectively ameliorate the cardinal symptoms of diabetes (polydipsia, polyphagia, and weight loss) in C57BL/6 diabetic mice, while concurrently modulating glucose and lipid profiles (TC, TG, HDL-C), highlighting their therapeutic potential for Type 2 diabetes[105]. Recent studies have provided deeper insights into the molecular mechanisms and the involvement of the gut microbiome. Liu et al. demonstrated that in high-fat diet/STZ-induced diabetic mice, Brasenia polysaccharide fractions (BSP-1a and BSP-U100) not only potentiated the insulin-PI3K/Akt signaling pathway but also enriched beneficial taxa (e.g., Lactobacillus). This microbial remodeling further upregulated hepatic PI3K and Akt expression, thereby exerting a synergistic hypoglycemic effect via the 'gut-liver axis'[106].
Antioxidant, antibacterial, and anti-inflammatory
-
Flavonoids and polysaccharides represent the fundamental bioactive constituents underpinning the antioxidant and antibacterial properties of Brasenia schreberi. Research has identified quercetin-7-O-β-D-glucopyranoside as a potent antioxidant agent isolated from the plant[36]. Comparative analyses across geographical origins confirmed a robust positive correlation between total flavonoid content and antioxidant capacity; furthermore, monomers such as quercetin, luteolin, and kaempferol displayed significant inhibitory efficacy against Proteus spp., Escherichia coli, and Pseudomonas aeruginosa[37].
Beyond low-molecular-weight phenolics, Brasenia polysaccharides exhibit unique advantages in alleviating oxidative stress. Utilizing the Caenorhabditis elegans model, Wang et al. verified that these polysaccharides not only scavenge free radicals but also upregulate critical antioxidant enzymes, catalase (CAT), superoxide dismutase (SOD), and glutathione S-transferase (GST), by modulating the skn-1 and daf-16 signaling axes. This regulation significantly extended nematode survival under acrylamide-induced toxicity[107,108], providing a robust scientific foundation for applications in functional foods, such as antioxidant-fortified yogurt[107].
Antitumor
-
Investigations into the antitumor potential of Brasenia schreberi have predominantly centered on its polysaccharide fractions. Chen et al. observed that crude polysaccharides significantly inhibited the proliferation of human cervical cancer HeLa cells in a time- and dose-dependent manner. Mechanistic studies indicated that polysaccharide treatment induced cell death via the disruption of membrane integrity and the induction of morphological alterations[109].
Insecticidal and other activities
-
Beyond the primary bioactivities described above, Brasenia polyphenols possess pleiotropic pharmacological properties, including anti-adipogenic, dermatological protective, and antiparasitic effects. Shimoda et al. characterized a series of polyphenols, including ethyl gallate, junsainoside A, and gossypetin, revealing that specific compounds not only exert anti-adipogenic effects but also protect skin fibroblasts against TNF-α-induced injury while stimulating hyaluronic acid synthesis[38]. In antiparasitic research, Vahekeni et al. validated the ethnomedicinal utility of Brasenia in Angolan traditional medicine, confirming that extracts and constituents (such as gallic acid derivatives and poly-galloyl glucose) possess significant inhibitory activity against Trypanosoma, the etiological agent of African sleeping sickness[28].
-
This review underscores the distinct phytochemical disparities among genera within the Nymphaeales, which underpin the diversity of their pharmacological mechanisms. Nymphaea and Brasenia are distinguished by a rich profile of flavonoids (e.g., myricetin, quercetin) and phenolic acids. The bioactivities of these species, notably antioxidant, hepatoprotective, and neuroprotective effects, are primarily orchestrated by ROS scavenging and the modulation of oxidative stress pathways. For instance, their protection against ischemic and hepatic injury is largely attributed to the capacity of polyphenols to upregulate endogenous antioxidant enzymes (SOD, GSH) and suppress pro-inflammatory mediators. Conversely, Nuphar presents a unique chemotaxonomic profile characterized by sulfur-containing sesquiterpene alkaloids. Diverging from the broad cytoprotective nature of polyphenols, these alkaloids exhibit potent, specific cytotoxicity, immunosuppression, and antiparasitic activities. This dichotomy suggests that while Nymphaea and Brasenia are promising candidates for functional foods targeting chronic diseases (e.g., diabetes, inflammation), Nuphar holds superior potential for the development of targeted therapeutics against neoplasms and autoimmune disorders.
Notwithstanding the promising landscape revealed by current literature, critical gaps persist that hinder the translation of these findings into clinical and industrial realms. First, the preponderance of in vitro assays and elementary animal models presents a limitation. While cell-based models are effective for screening cytotoxicity or enzyme inhibition, they fail to recapitulate bioavailability, metabolism, and pharmacokinetic (ADME) profiles. For instance, flavonoids undergo extensive biotransformation in the intestine and liver; however, few studies have validated whether the effective concentrations observed in vitro are physiologically achievable in vivo. Second, the absence of standardized extraction protocols serves as a barrier. The diversity of solvents employed, ranging from methanol and ethanol to water, results in significant heterogeneity in metabolite recovery and bioactivity outcomes. This inconsistency complicates lateral comparisons across studies and impedes the quality control necessary for drug development. Third, investigations into the 'gut-liver axis' remain in their nascence. While recent insights into Brasenia polysaccharides have implicated gut microbiota modulation, analogous studies for Nymphaea and Nuphar are sparse. Elucidating the interplay between these phytochemicals and the microbiome could reveal novel metabolic pathways and therapeutic targets.
To surmount these challenges, future research endeavors should integrate multi-omics strategies, encompassing genomics, transcriptomics, and metabolomics, to systematically mine metabolic genes and delineate the biosynthetic pathways of key bioactive compounds. Such knowledge is critical for molecular breeding initiatives designed to concurrently enhance ornamental traits (floral coloration, scent) and functional metabolite accumulation. Moreover, the water lily industry is transitioning from traditional landscaping and cut flower markets toward high-value-added sectors. By capitalizing on their specific bioactive potentials, water lily resources can be diversified into functional foods, natural cosmetics, aromatherapy, and pharmaceuticals. This paradigm of 'multi-purpose' utilization is poised to significantly elevate the economic value and sustainability of the industry.
-
The authors confirm contributions to the paper as follows: study conception and design: Chen F; literature review, data analysis: Li J; draft manuscript preparation: Chen F, Li J. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
This work was supported by the National Natural Science Foundation of China (Grant No. 32172614).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li J, Chen F. 2026. Secondary metabolites and biological activities of water lilies: a comprehensive review. Ornamental Plant Research 6: e019 doi: 10.48130/opr-0026-0008 

Secondary metabolites and biological activities of water lilies: a comprehensive review
- Received: 21 November 2025
- Revised: 24 January 2026
- Accepted: 09 March 2026
- Published online: 31 May 2026
Abstract: Plants of the order Nymphaeales are representative tropical and subtropical ornamental aquatic species, valued for their diverse flower colors, distinctive floral scents, and strong environmental adaptability. Although extensive studies have reported their chemical constituents, the roles of secondary metabolites in shaping ornamental traits remain insufficiently integrated. In this review, relevant literature published up to September 2025 was systematically retrieved from PubMed and Web of Science, and the major classes of secondary metabolites reported in representative genera (Nymphaea, Brasenia, and Nuphar) were summarized, with particular emphasis on their contributions to flower color and scent formation. Distinct chemotaxonomic patterns were identified: Nymphaea and Brasenia are predominantly enriched in flavonoids, polyphenols, and polysaccharides, whereas Nuphar is characterized by unique sulfur-containing sesquiterpene alkaloids. Accumulating evidence indicates that differential accumulation of specific anthocyanin derivatives underlies flower color diversification, while the composition and relative abundance of volatile organic compounds (VOCs) determine floral scent profiles among species and cultivars. These metabolite-based insights provide valuable targets for ornamental trait improvement, including flower color modification and fragrance-oriented breeding. In addition, the bioactive metabolites enriched in Nymphaeales support their further development as functional ornamental plants and value-added horticultural resources. This review highlights the integrative potential of linking metabolite diversity with ornamental trait formation to advance the sustainable utilization of tropical ornamental aquatic plants.
-
Key words:
- Water lilies /
- Ornamental traits /
- Secondary metabolites /
- Flower color /
- Floral scent /
- Molecular breeding