








-
Dendrobium is one of the largest genera of orchids, with most species distributed in tropical and subtropical Asia and eastern Australia[1]. Importantly, interest in Dendrobium species is broad, ranging from traditional medicine and specialized cosmetic materials to ornamental horticulture[2−4]. Nowadays, Dendrobium orchids have become increasingly popular for ornamental use due to their floriferous nature and availability in a wide range of colors, sizes, shapes, and prolonged flowering periods. Among the large number of species and cultivars of Dendrobium, two groups are suited for ornamental sale: the Nobile-group and the Denphal-group. The Nobile-group produces inflorescences and flowers that are distributed along the pseudobulbs, while the Denphal-group produces one or more terminal inflorescences from the tip of pseudobulbs[5].
The Denphal-group Dendrobium is most commonly distributed in tropical regions such as the Philippines, Malaysia, Indonesia, and other South Pacific island countries, and is most cultivated in tropical and subtropical regions such as Thailand, Singapore, Malaysia, etc., where the overall temperature is suitable for growth. The optimal ambient temperatures for Denphal-group Dendrobium spp. and their hybrids occur when night temperatures stay above 18 °C, with daytime temperatures ranging between 24 and 29 °C. Low temperatures (below 15 °C) result in significant limitations on the growth and development of Denphal-group Dendrobium spp. and their hybrids, such as leaf discoloration, foliage loss, and diminished vegetative growth. Moreover, lower temperatures and shorter daylight periods have been observed to alter the concentration of internal growth regulators, prompting the initiation of flowering in Dendrobium orchids[6]. Therefore, in the majority of subtropical regions, uncertain cold waves cause low temperature damage to Denphal-group Dendrobium plants, including gradual leaf yellowing, slower growth, and decreased flower longevity[7]. Under low temperature stress, the relative electrical conductivity (REC), as well as soluble protein, soluble sugar, free proline, malondialdehyde (MDA) content, and defoliation rate of Denphal-group Dendrobium cultivars increased with the decrease of treatment temperature and the extension of treatment time, while the content of chlorophyll decreased gradually.
Dendrobium Sonia 'Hiasakul', a representative cultivar of the Denphal group and a popular commercial variety at present, is highly sensitive to low temperatures. Its cold damage symptoms include rapid yellowing and falling of leaves, damage to membrane structure, decline in photosynthetic capacity and metabolic disorder[7]. Comparative physiological and transcriptomic studies with the cold-tolerant cultivar Den. 'Hongxing' revealed that Den. Sonia 'Hiasakul' suffers more acute morphological damage under low-temperature conditions, characterized by a higher leaf abscission rate, accelerated leaf yellowing, a more rapid increase in REC and MDA content, and a sharper decline in chlorophyll levels[8]. Transcriptomic investigations have further identified significant enrichment of cold-responsive genes and metabolic pathways associated with the observed physiological traits, including those involved in signal transduction, plant hormones, transcription factors, protein translation and modification, functional proteins, biosynthesis and metabolism, cellular structure, light signaling, and the circadian rhythm[8,9]. Compared with other Dendrobium species, the drought and salt tolerance of Sonia 'Hiasakul' remains uninvestigated. Its tropical origin, however, suggests potential sensitivity to both drought and salinity—a hypothesis that awaits further experimental validation.
Previous studies have shown that plant chilling stress involves cooperative regulation of multiple transcription factors. The three main cold-responsive gene families in plants are Inducer of CBF Expression (ICE), C-repeat Binding Factors (CBFs), and the Cold-Regulated genes (CORs)[10]. These three abovementioned key players form an imperative signaling pathway, the ICE-CBF-COR cascade, which alleviates cold stress in plants[11]. Nowadays, the ICE-CBF-COR signal transduction pathway has been widely studied and has been shown to play a crucial role in the regulation of plant chilling tolerance[12]. In the pathway, COR genes, as key regulators, are considered to be those most closely related to the cold stress response. COR413, a member of the COR gene family, is a subfamily of low-temperature-responsive genes unique to plants. The COR413 protein is classified into three forms based on its subcellular localization: COR413PM in the plasma membrane, COR413TM in the inner capsule membrane, and COR413IM in the chloroplast inner membrane[13]. Since being first identified in Arabidopsis, COR413 genes have been identified in several other plants, such as peach[14], wintersweet[15], tomato[16,17], Saussurea involucrata[18], Saccharum spontaneum[19], and Phlox subulata[20]. These studies showed that COR413 genes respond to low temperature and enhance plant cold tolerance, as well as other abiotic stresses such as drought and salt.
Studies on the COR413 gene have been conducted in various plant species; however, this gene has not been investigated in Dendrobium. Therefore, in this study, DhCOR413PM1, a member of the COR413 family, was identified in Denphal-group Dendrobium. The expression pattern of DhCOR413PM1 was analyzed across various organs and under different abiotic stresses. Transgenic Arabidopsis thaliana plants overexpressing DhCOR413PM1 were evaluated under low-temperature, drought, and salt stress conditions. Collectively, the findings provide insights into the role of DhCOR413PM1 in the cold, drought, and salt stress resistance mechanisms of Denphal-group Dendrobium.
-
The Dendrobium Sonia 'Hiasakul' used in this study was cultivated at the Tropical Flower Resource Garden, Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences (CATAS), located in Danzhou, Hainan Province, China.
Wild-type Arabidopsis thaliana (ecotype Columbia-0, Col-0) for genetic transformation was maintained by the Tropical Crops Genetic Resources Institute, CATAS. Arabidopsis seeds were surface-sterilized with 15% (v/v) sodium hypochlorite (NaClO) for 10 min and rinsed five times with sterile distilled water. Sterilized seeds were then plated on solid Murashige and Skoog (MS) medium supplemented with Hygromycin B (30 mg/L) for the selection of transformants. After two weeks, positive seedlings were transplanted into the soil. All Arabidopsis plants were grown in a controlled environment growth chamber under the following conditions: 22 °C, 85% relative humidity, and a 16 h light/8 h dark photoperiod cycle.
Cloning and bioinformatic analysis of DhCOR413PM1
-
The sequence information for DhCOR413PM1 was obtained from the transcriptome of the Dendrobium cultivar Sonia 'Hiasakul'. The full-length coding sequence (CDS) of DhCOR413PM1 was subsequently cloned from Sonia 'Hiasakul' cDNA using gene-specific primers (F: 5'-ATGGGAAAACTAGGGTTCCTAGCG-3', R: 5'-CTAAATCAAAATGACAATGAGTCC-3').
Amino acid sequences of COR413 proteins from diverse plant species were retrieved from the National Center for Biotechnology Information (NCBI) database. The deduced DhCOR413PM1 amino acid sequence was aligned with homologous sequences from closely related species using DNAMAN 6.0 software. A phylogenetic tree was constructed from this alignment using the neighbor-joining (NJ) method implemented in MEGA 11 software, with bootstrap values set to 1,000 replicates. Conserved domain analysis of the DhCOR413PM1 protein was performed using the NCBI Conserved Domain Database (CDD,
www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). Transmembrane domains were predicted using the TMHMM Server v.2.0 (www.cbs.dtu.dk/services/TMHMM/ ). The subcellular localization of DhCOR413PM1 was predicted using a suite of online tools: WoLF PSORT (https://wolfpsort.hgc.jp/ ) for general localization; TargetP-2.0 (https://services.healthtech.dtu.dk/services/TargetP-2.0/ ) for N-terminal signal peptide analysis; and LOCALIZER (http://localizer.csiro.au/ ) for the specific detection of chloroplast, mitochondrial, or nuclear localization signals. All analyses were run with default parameters.Expression analysis of DhCOR413PM1 in Dendrobium
-
For tissue-specific expression analysis of DhCOR413PM1 in Dendrobium, the roots, stems, and leaves from young seedlings and middle-aged plants were collected. For mature plants, samples included roots, stems, leaves, peduncles, inflorescences, and flower buds at different development stages.
For abiotic stress expression analysis of DhCOR413PM1, middle-aged plants were subjected to low temperature (4 °C), high salt (50 mM NaCl), and drought (15% PEG-6000) treatments. Leaves from five plants per treatment were harvested at 0, 1, 2, 4, 8,12, 24, 48, 72, 96, and 120 h post-treatment. All samples were immediately frozen in liquid nitrogen and stored at −80 °C.
Total RNA was extracted using the RNAprep Pure Plant Plus Kit (TIANGEN, Nanjing, China). First-strand cDNA synthesis was performed with the PrimeScript™ FAST RT reagent Kit (Takara, Dalian, China). Quantitative real-time PCR (qRT-PCR) assays were conducted using TB Green® Premix Ex Taq™ II FAST (Takara, Dalian, China) with β-actin (F: 5'-CTTCGTCTTCCACTTCAG-3' and R: 5'-ATCATACCAGT CTCAACAC-3') as the reference gene[21]. The primer sequences used for qRT-PCR were F: 5'-GGATTCGGCACATACTTTCTC-3' and R: 5'-CCATTTTCCAACCTCACCTC-3' respectively. The 2−ΔΔCq method was employed to calculate relative expression levels. Three biological replicates were analyzed, with technical triplicates for each sample.
Vector construction and plant transformation
-
To validate the function of DhCOR413PM1, the full-length coding sequence was cloned into the pBWA(V)HS vector using the Eco31I (BsaI) restriction sites, generating the recombinant plasmid pBWA(V)HS-DhCOR413PM1. After confirmation by PCR and Sanger sequencing, the construct was transferred into Agrobacterium tumefaciens strain GV3101.
Arabidopsis thaliana transformation was performed using the floral dip method[22]. T1 transgenic plants were selected on Murashige and Skoog (MS) medium supplemented with 30 mg/L hygromycin B. Integration of the transgene was confirmed by PCR amplification with vector-specific primers (F:5'-CTTCGCAAGACCCTTCCTC-3'; R:5'-ATGACAATGAGTCCCCAGA-3'). In addition, the expression of DhCOR413PM1 in T3 homozygous lines was quantified by qRT-PCR using the Arabidopsis Actin gene as the reference (F:5'-CTTCGTCTTCCACTTCAG-3'; R:5'-ATCATACCAGTCTCAACAC-3'). Three independent overexpression lines (OE1, OE3, OE9) exhibiting high transgene expression levels were selected for subsequent abiotic stress assays.
Abiotic stress tolerance assay of DhCOR413PM1 transgenic Arabidopsis
-
To assay freezing tolerance in DhCOR413PM1 transgenic Arabidopsis, wild-type and T3 generation transgenic seedlings were grown on MS medium for 12 d. The plates with the plants were then placed at 4 °C for 12 h in the dark, followed by exposure to −7 °C for 12 h. After the low-temperature treatment, the plates were transferred to normal growth conditions (22 °C, 16 h light/8 h dark) for 5 d of recovery. Photographs were taken before freezing, and after recovery, respectively, and the survival rate was calculated.
For dehydration and salt stress treatments, the transgenic and WT seeds were sown on MS medium containing 250 mM mannitol and 150 mM NaCl, respectively. After 3 d of cold vernalization at 4 °C, the seeds were then transferred into an artificial climate box for culture. Germination rate was scored after two weeks, and a radicle length of 1 mm was used as a criterion for germination. Seeds grown under non-stress conditions served as the control. In addition, one-week-old transgenic and wild-type seedlings were vertically cultured on MS medium containing 150 mM mannitol or 50 mM NaCl, respectively. And then the primary root length was measured after one week. Drought and salt tolerance in the transgenic seedlings were assessed through phenotypic evaluation under these stress treatments.
Data analysis
-
Statistical analysis was conducted using SPSS Statistics 24. All data were obtained from three independent biological replicates and are presented as the mean. Statistical differences among treatments were analyzed by one-way ANOVA followed by Duncan's New Multiple Range Test (DNMRT).
-
The complete CDS sequence of the cold-adapted protein gene from Dendrobium Sonia 'Hiasakul' was successfully amplified (Fig. 1a). This gene is presumed to belong to the COR413-PM subclass of the COR413 protein family and was therefore named DhCOR413PM1. Sequence analysis indicated that DhCOR413PM1 contains 612 bases and encodes 203 amino acids with a calculated molecular weight of 22.42 kDa.
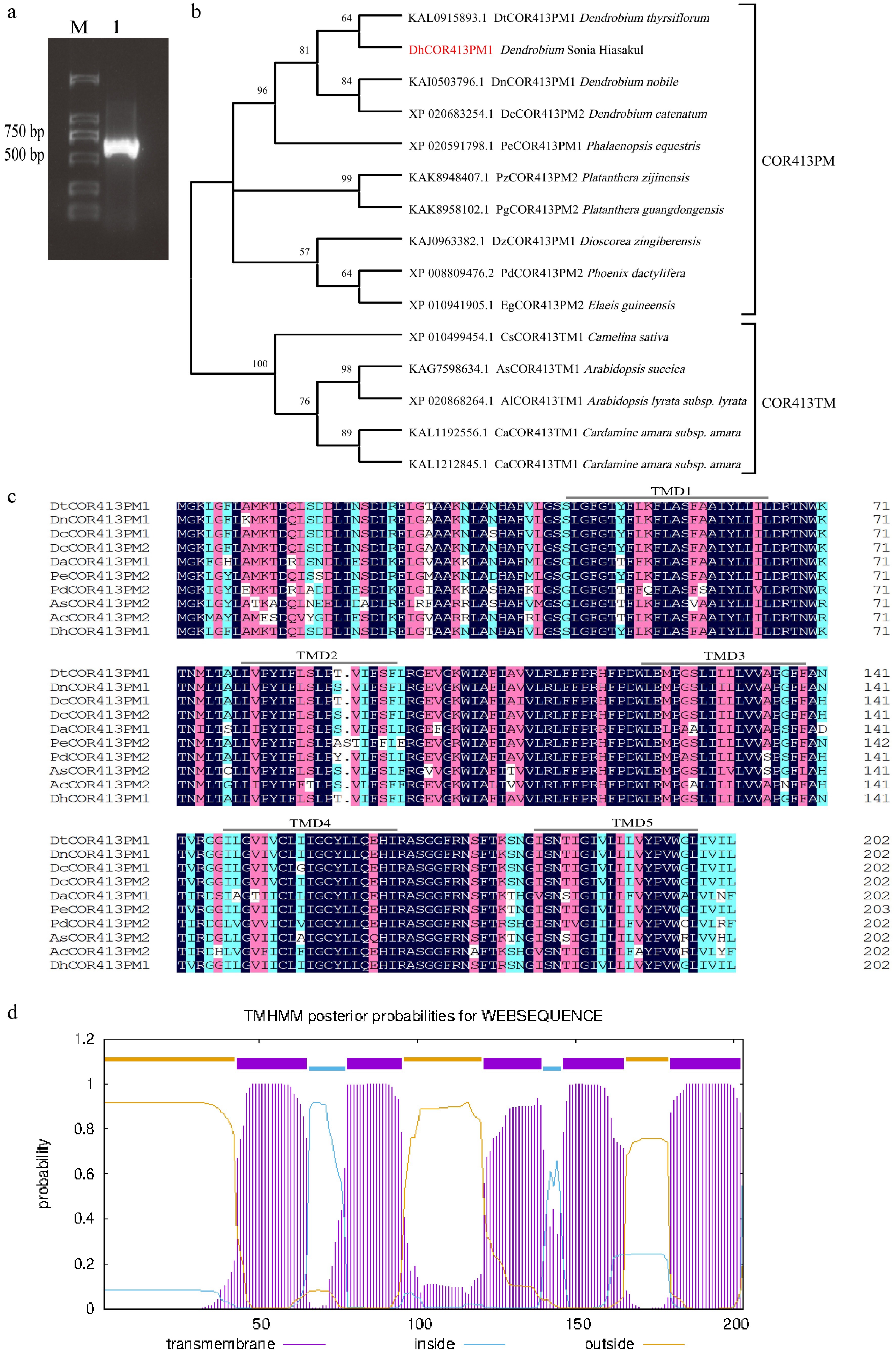
Figure 1.
Amplified CDS product, phylogenetic analysis, and sequence comparison of COR413 in Dendrobium. (a) Amplification of the DhCOR413PM1 CDS from Dendrobium. M, Marker; 1, DhCOR413PM1. (b) Phylogenetic analysis of DhCOR413PM1. (c) Sequence alignment of COR413 proteins across plant lineages. Conserved transmembrane domains (TMD1–5) were presented in lines. (d) The putative transmembrane domains of DhCOR413PM1.
Using the NCBI database, 14 evolutionarily conserved COR413 protein sequences from diverse plant species were identified and selected for phylogenetic analysis. BLASTP analysis revealed that DhCOR413PM1 shares extremely high amino acid sequence identity (96.55%) with DtCOR413PM1 from Den. thyrsiflorum (Accession: KAL0915893.1). Phylogenetic analysis showed that Den. Sonia 'Hiasakul' clustered in the same branch as Den. thyrsiflorum (Fig. 1b). The results showed that DhCOR413PM1 is most closely related to DtCOR413PM1 from Den. thyrsiflorum. Furthermore, multiple sequence alignment revealed a high degree of evolutionary conservation and significant homology between DhCOR413PM1 and COR413 proteins from other species (Fig. 1c). Analysis using the NCBI CDD and TMHMM online tools indicated that DhCOR413PM1 contains one full COR413 conserved domain (amino acids 10–190) and five transmembrane domains (Fig. 1c, d), which are characteristic features of the COR413 protein family. WoLF PSORT predicted that DhCOR413PM1 is localized to the plasma membrane. TargetP-2.0 and LOCALIZER further showed that it lacks chloroplast, mitochondrial, and nuclear targeting signals but possesses a signal peptide. These consistent results confirm its plasma membrane localization, supporting its classification as a COR413PM protein.
Expression patterns of DhCOR413PM1 in Dendrobium
-
The tissue-specific expression pattern of DhCOR413PM1 was examined by qRT-PCR to characterize its potential function in Dendrobium. The results showed that DhCOR413PM1 was ubiquitously expressed across all tissues but exhibited significant spatial divergence. The highest transcript accumulation occurred in roots at three developmental stages (juvenile, vegetative, and reproductive), while leaves of mid-vegetative and mature plants showed minimal expression (Fig. 2a). This gene exhibited high expression in reproductive organs. Its transcript level was relatively high in 1 cm inflorescence meristems (IM), followed by an upward trend peaking in 3 cm IM, and subsequently declined gradually. Notably, detectable expression persisted even in 9 cm IM (Fig. 2b). During floral bud development, significant expression was observed, with the highest level in 1 mm diameter buds. Expression progressively decreased from 1 to 9 mm diameter buds (Fig. 2b). To further analyze expression dynamics during floral development, four developmental stages of floral organs were examined. DhCOR413PM1 was expressed throughout all floral whorls, with the strongest signals detected in the labellum and gynandrium of 5 mm floral buds. In contrast, all organs exhibited reduced expression in fully opened flowers (Fig. 2c).
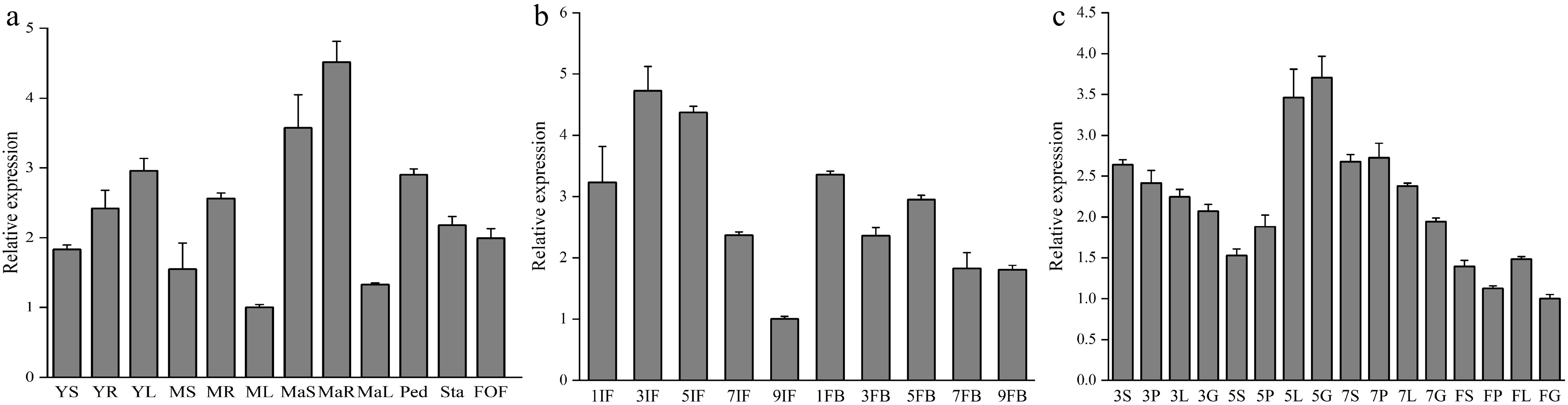
Figure 2.
Expression of DhCOR413PM1 in different tissues and floral organs during development. (a) Expression of DhCOR413PM1 in different tissues at various development stages. YS,YR, and YL are stem, root, and leaf of young seedlings (juvenile stage), respectively; MS, MR, and ML are stem, root, and leaf of middle-age seedling (vegetative stage), respectively; MaS, MaR, and MaL are stem, root, and leaf of mature plant (reproductive stage), respectively; Ped, peduncle; Sta, stalk; FOF, fully open flower. (b) Expression of DhCOR413PM1 in inflorescences and flower buds at different development stages. IF, inflorescence meristem (1–9 cm in length); FB, flower bud (1–9 mm in diameter). (c) Expression of DhCOR413PM1 in floral organs at different development stages (3–7 mm diameter flower bud and fully open flower). S, Sepals; P, Petals; L, Labellum; G, gynandrium. Error bars represent ± SE of the three biological replicates.
Abiotic stresses inducible expression of DhCOR413PM1
-
To further investigate the potential roles of the DhCOR413PM1 in response to abiotic stress, transcript abundance was analyzed in leaves of middle-aged plantlets of Dendrobium Sonia 'Hiasakul' under low temperature (4 °C), drought (15% PEG-6000) and salt (50 mM NaCl) stress. Under 4 °C treatment, DhCOR413PM1 expression was initially suppressed (1–2 h) but strongly induced at 4–12 h. The highest expression level was observed at 8 h, reaching 2.13 times that of the control group (Fig. 3a). Under 15% (w/v) PEG-6000-induced osmotic stress, DhCOR413PM1 expression was significantly downregulated during 1–12 h, but upregulated at 24–48 h, peaking at 24 h with 1.57-fold induction compared to the control (Fig. 3b). Under 50 mM NaCl treatment, DhCOR413PM1 expression was rapidly upregulated within 1 h, followed by a slight decrease from 2–4 h. It peaked initially at 8 h, then gradually declined, ultimately reaching its highest level (1.88-fold higher than controls) at 120 h (Fig. 3c).
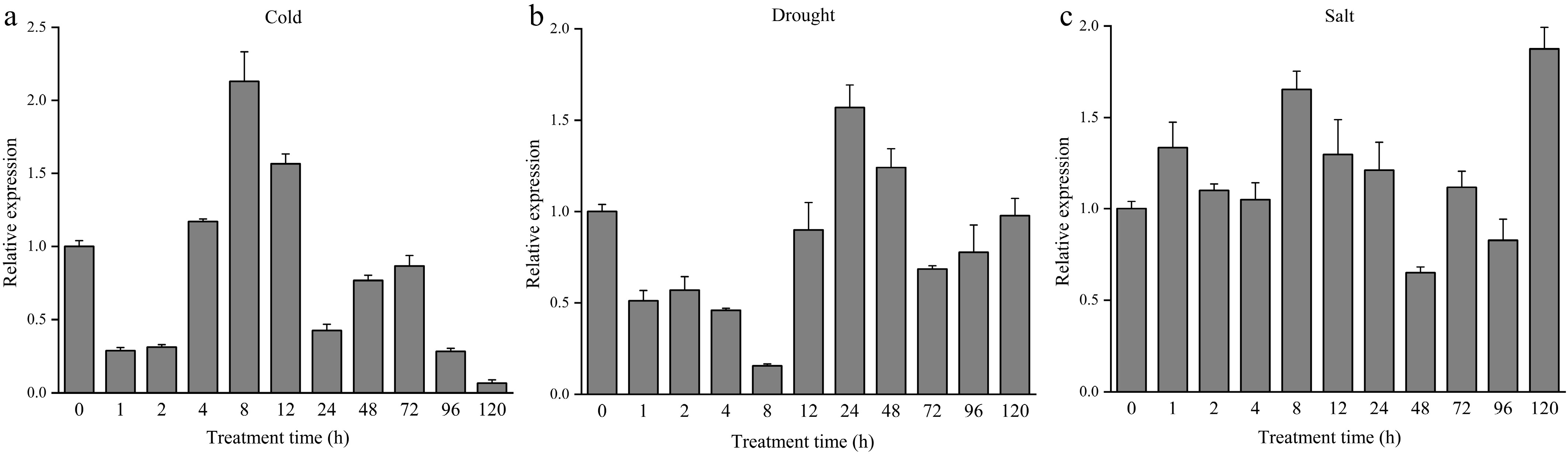
Figure 3.
Expression of DhCOR413PM1 in Dendrobium under abiotic stresses over time. (a) Expression of DhCOR413PM1 under low-temperature treatment (4 °C). (b) Expression of DhCOR413PM1 under drought stress (15% PEG-6000). (c) Expression of DhCOR413PM1 under salt stress (50 mM NaCl). Error bars represent ± SE of the three biological replicates.
In contrast to low-temperature (4 °C) and drought (15% PEG-6000) treatments, where upregulation occurred only after 8 and 24 h, respectively, NaCl induced rapid transcriptional activation of DhCOR413PM1 within 1 h. These findings demonstrate that DhCOR413PM1 is potentially involved in responses to multiple abiotic stresses. Notably, its transcriptional activation under salt stress (50 mM NaCl) occurred more rapidly than under cold (4 °C) or drought (15% PEG-6000) treatments.
Cold tolerance analysis of DhCOR413PM1 overexpression transgenic Arabidopsis
-
To functionally characterize DhCOR413PM1, Arabidopsis thaliana overexpressing DhCOR413PM1 was generated via the floral dip transformation. Ten independent T1 transgenic lines were confirmed by PCR amplification using primers targeting the 35SCaMV promoter and DhCOR413PM1 coding sequence (Fig. 4a). Subsequent qRT-PCR analysis revealed DhCOR413PM1 expression in eight lines, with OE1, OE3, and OE9 exhibiting the highest transcript levels (Fig. 4b). The T3 homozygous progenies of these three high-expression lines were selected for the subsequent freezing, dehydration, and salttolerance assays.
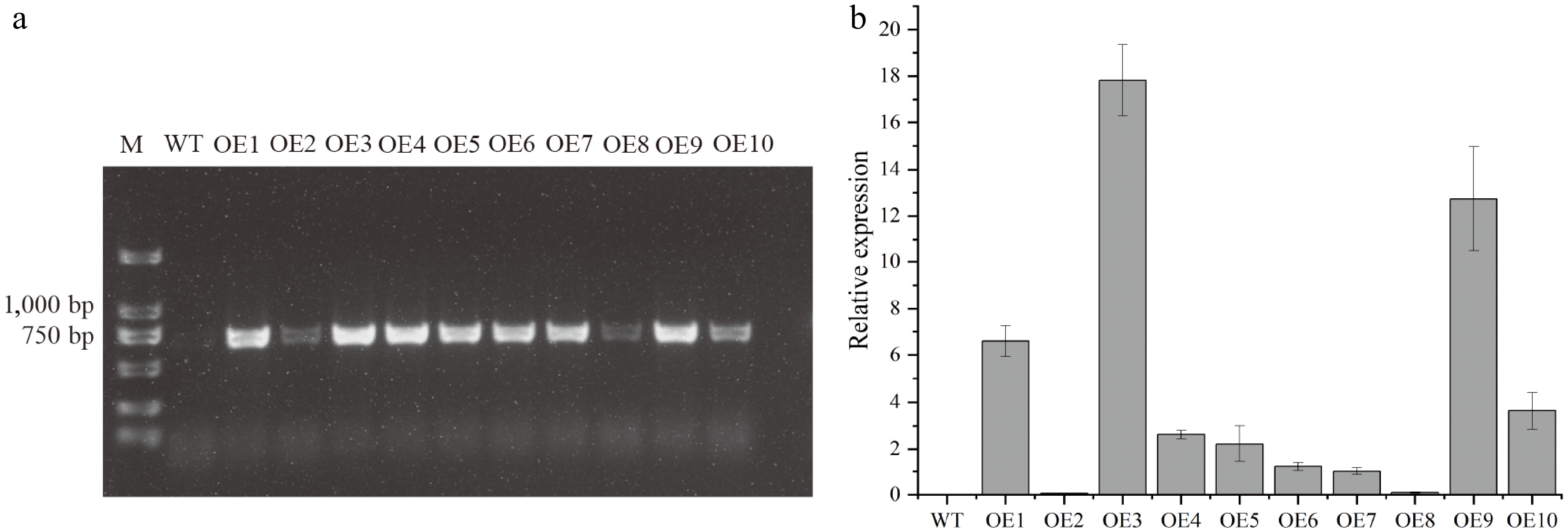
Figure 4.
Transgenic validation and gene expression in Arabidopsis thaliana. (a) PCR verification of DhCOR413PM1 integration using CaMV35S promoter and gene-specific primers. M: Marker; WT: Wild-type; OE1–10: Independent transgenic lines. (b) DhCOR413PM1 transcript levels in T3 homozygous lines as quantified by qRT-PCR. Error bars represent ± SE of the three biological replicates.
Under normal conditions, no morphological differences were observed between the transgenic plants and wild type (Fig. 5a). Following cold acclimation (4 °C for 12 h) and subsequent freezing stress (−7 °C for 12 h), transgenic plants suffered less severe freezing damage than wild-type (WT) controls. After a 5-d recovery period, WT plants showed significantly higher mortality (44%) than transgenic lines (Fig. 5b). The survival rates of the transgenic plants (68.4% for OE9, 68% for OE3, and 64.7% for OE1) were higher than that of WT plants (56%) (Fig. 5c). These results demonstrate that overexpression of DhCOR413PM1 enhances cold tolerance in Arabidopsis.

Figure 5.
Freezing tolerance analysis of 35S::DhCOR413PM1 overexpressing Arabidopsis. (a) Phenotypes under non-stress conditions. (b) Phenotypes after freezing stress. (c) Survival rates after freezing stress. WT: wild-type; OE: 35S::DhCOR413PM1 overexpression lines. Error bars represent ± SE of the three biological replicates. Different letters above the bars indicate significant differences (p < 0.01) according to the DNMRT.
Overexpression of DhCOR413PM1 enhanced dehydration and salt tolerance in Arabidopsis
-
Abiotic stress expression analysis showed that DhCOR413PM1 expression was induced not only by low temperature treatment, but also by salt and drought treatments in Sonia 'Hiasakul'. Therefore, the drought and salt tolerance of transgenic Arabidopsis overexpressing DhCOR413PM1 was evaluated using mannitol (simulated drought) and NaCl treatment, respectively.
At 12 d post-sowing, DhCOR413PM1 overexpressing lines exhibited higher germination rates than the wild-type (WT) under osmotic stress (250 mM mannitol) and salinity stress (150 mM NaCl) (Fig. 6b–e), with no phenotypic difference observed in the controls (Fig. 6a). Moreover, radicle emergence in wild-type (WT) was significantly inhibited compared to transgenic lines on media containing these stressors (Fig. 6b, c).
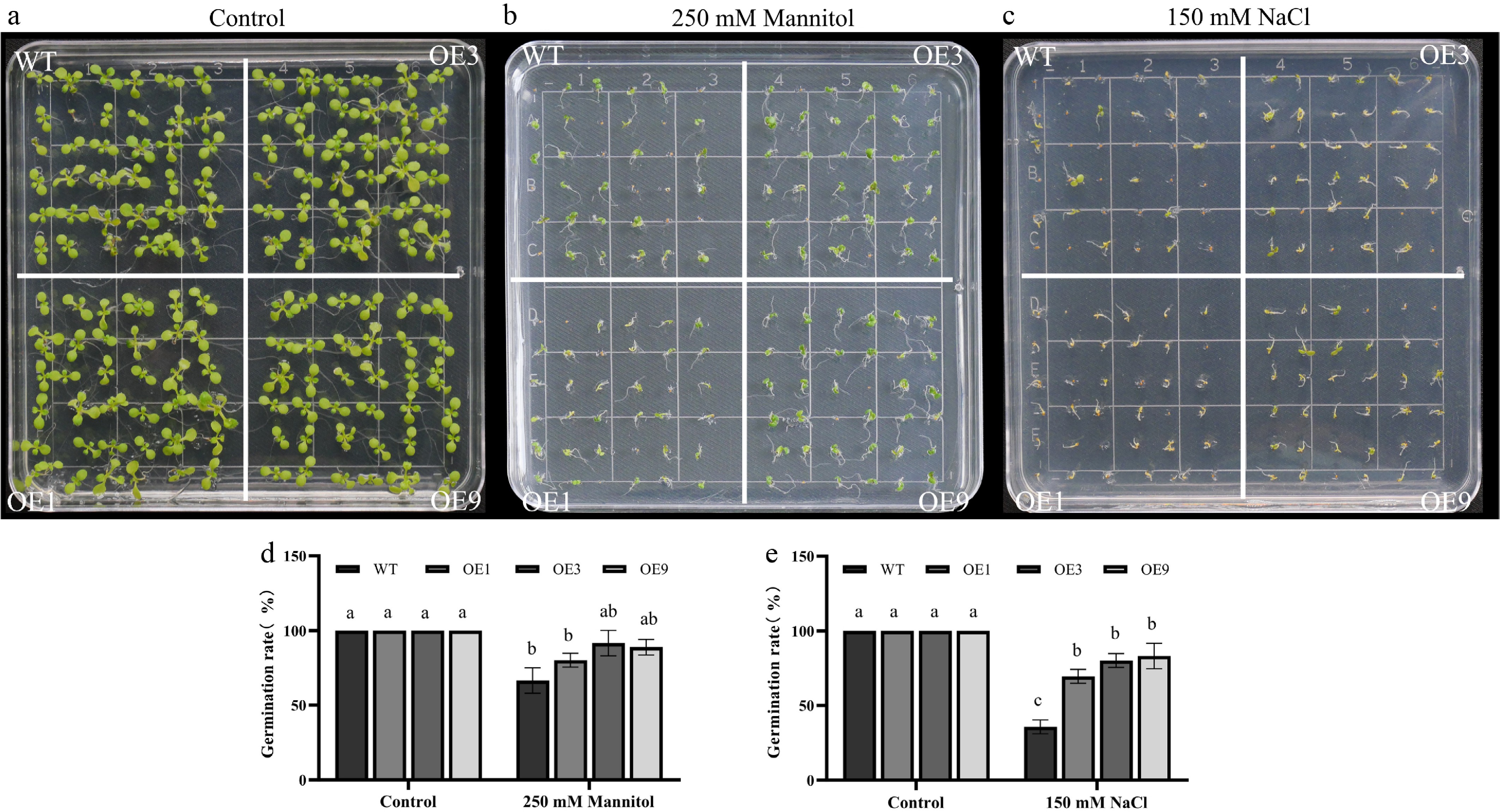
Figure 6.
Drought and salinity tolerance of 35S::DhCOR413PM1 overexpressing Arabidopsis. (a) Phenotypes under control conditions. (b) Response to osmotic stress (250 mM mannitol). (c) Response to salt stress (150 mM NaCl). (d) Germination rates under osmotic stress. (e) Germination rates under salt stress. WT: wild-type; OE: 35S::DhCOR413PM1-overexpression lines. Error bars represent ± SE of the three biological replicates. Different letters above the bars indicate significant differences (p < 0.01) according to the DNMRT.
There was no significant difference in root length between the transgenic lines and the wild type under normal conditions (Fig. 7a). However, compared with the root length of DhCOR413PM1 overexpression transgenic lines, wild type plants showed shorter roots when exposed to 150 mM mannitol (Fig. 7b, d). Under 50 mM NaCl stress, the root length of both the WT and DhCOR413PM1 overexpression transgenic plants was significantly reduced compared with normal conditions, but the wild-type plants were significantly more sensitive to salt stress than the transgenic lines (Fig. 7c, e). Collectively, these results suggest that overexpression of DhCOR413PM1 confers enhanced drought and salinity stress tolerance in transgenic Arabidopsis thaliana.
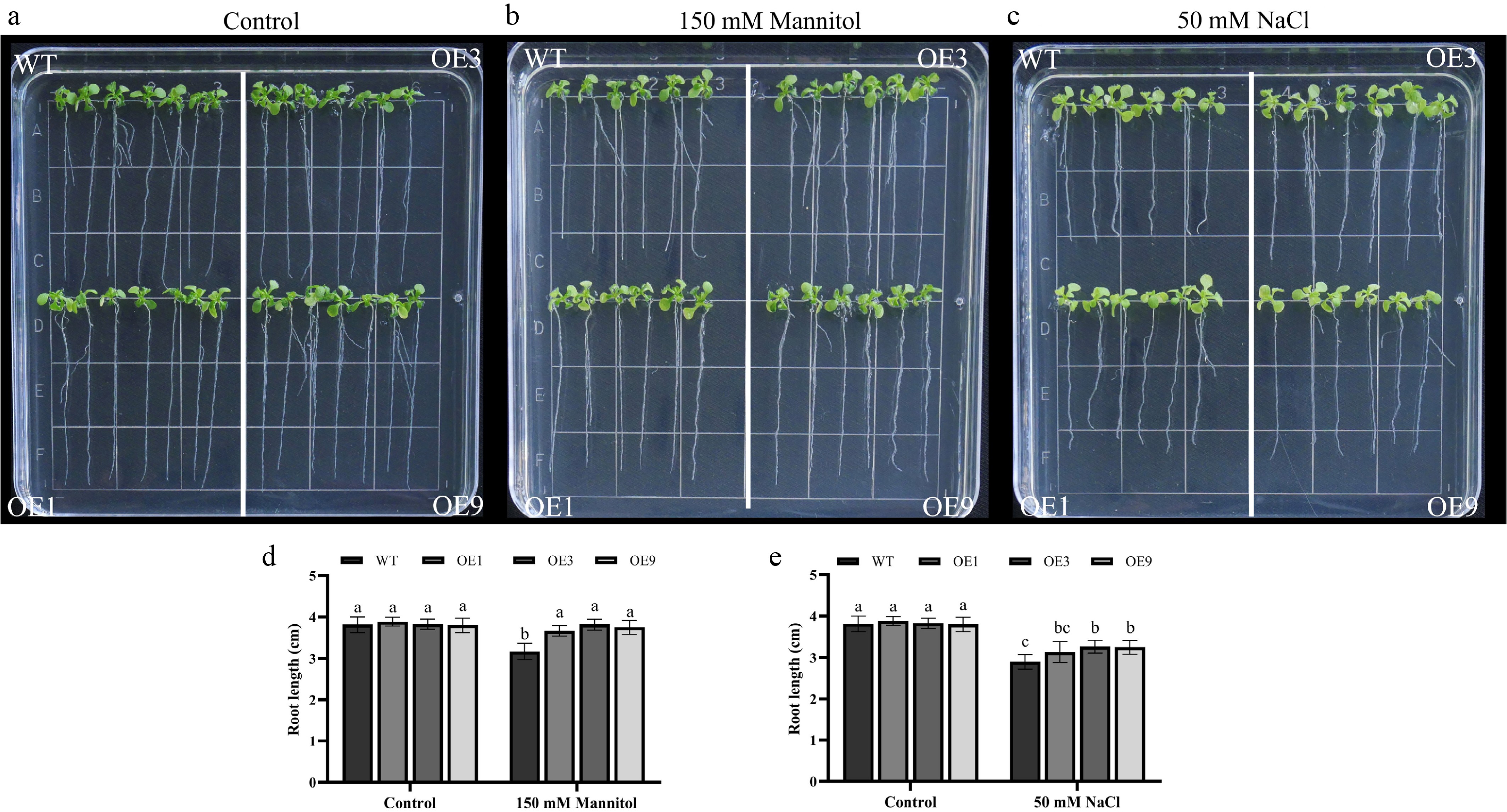
Figure 7.
Root elongation analysis of 35S::DhCOR413PM1 overexpressing Arabidopsis under osmotic and salt stress. (a) Root growth status under normal conditions. (b) Response to osmotic stress (150 mM mannitol). (c) Response to salt stress (50 mM NaCl). (d) Primary root length under osmotic stress. (e) Primary root length under salt stress. WT: wild-type; OE: 35S::DhCOR413PM1 overexpression lines. Error bars represent ± SE of the three biological replicates. Different letters above the bars indicate significant differences (p < 0.01) according to the DNMRT.
-
Low temperature is an important environmental stress that directly affects the survival and development of plants[23,24]. Low temperature stress mainly affects plants by causing damage to the membrane system, resulting in increased membrane permeability, inactivation of membrane-associated enzymes, and ultimately inducing disorder of cell metabolism and function[25−27]. Therefore, maintaining the integrity and stability of the membrane system is crucial for plants to withstand chilling stress. COR413, as a member of the COR gene family, represents a group of plant-specific, low-temperature-responsive genes[28]. COR413 proteins are mostly located in the membrane of organelles in plant cells: COR413PM in the plasma membrane, COR413TM in the inner capsule membrane, and COR413IM in the chloroplast inner membrane[13]. Moreover, the COR413 protein family is evolutionarily conserved across diverse plant lineages, including cereal crops such as wheat[29]. In this study, DhCOR413PM1 was determined to encode a 203-amino-acid protein with a calculated molecular mass of 22.42 kDa. The protein contains a completely conserved COR413 domain (residues 10–190) and five transmembrane domains (Fig. 1c, d). Subcellular localization predictions indicate that DhCOR413PM1 is located in the plasma membrane. The analysis of conserved and transmembrane domains in this study is consistent with previous reports in other plant species, e.g., wintersweet[15] and wheat[29].
The low temperature sensitivity of different plant tissues varies[15,30]. In the present study, the expression levels of DhCOR413PM1 showed tissue-specific variation. The highest expression levels were detected in the roots of plants at three development stages, a pattern similar to that observed in wintersweet, which also exhibited the highest expression of CpCOR413PM1 (Chimonanthus praecox) in roots[15]. This may indicate that the roots of Dendrobium are more sensitive to low temperatures than other tissues. In leaves, the expression level of DhCOR413PM1 in seedling plants was higher than that in middle-aged plantlets and mature plants of Dendrobium. Previous studies have shown that the leaves of young Dendrobium plants suffer more severe damage from low temperature than those of middle-aged and mature plants[7,9], indicating that the leaves of young seedlings are more sensitive to low temperature than those of middle-aged and mature plants. DhCOR413PM1 expression peaked in inflorescence meristems (at the 3 and 5 cm stages) and in labellum and gynostemium tissues of 5 mm floral buds, exhibiting significantly higher transcript levels than other sampled tissues. Tissue expression analysis of CpCOR413PM1 in wintersweet showed higher expression levels in the stamen, pistil, and inner petals[15], suggesting that the labellum and gynostemium of Dendrobium are the most vulnerable to low-temperature chilling, similar to the inner organs of wintersweet.
The expression of DhCOR413PM1 in response to abiotic stresses showed that it was induced by cold stress and reached its highest expression level after 8 h of treatment at 4 °C. As a cold-responsive gene, COR413PM is most extensively induced by cold stress. For example, CpCOR413PM1 in wintersweet peaked at 12 h of 4 °C treatment[15], PsCor413pm2 in Phlox subulata also showed highest expression at 12 h of 4 °C treatment[20], SsCor413 in leaves of Saccharum spontaneum peaked at 24 h under cold treatment[19], and SikCOR413PM1 in Saussurea involucrate reached its highest expression at 6 h of low temperature treatment[31]. Functional characterization confirms that DhCOR413PM1 mediates cold stress regulatory pathways.
Previous studies showed that COR413PM is not only induced by low temperature, but also induced by drought or salt stress, as observed in PsCor413pm2 in Phlox subulata[20], LeCOR413PM2 in wild-type tomato[32], SsCor413 in Saccharum spontaneum[19], SikCOR413PM1 in Saussurea involucrate[31], and GbCOR413 in Gossypium barbadense[33]. In this study, the expression of DhCOR413PM1 increased under drought (15% PEG-6000) and high salt (50 mM NaCl) stress treatment. Under drought stress, DhCOR413PM1 expression was downregulated from 1–12 h but upregulated at 24–48 h, with a peak at 24 h (Fig. 3b). Under salt stress, expression was rapidly induced at 1 h, decreased slightly (2–4 h), and reached its maximum at 120 h after an initial peak at 8 h (Fig. 3c). As mentioned above, the research indicates that COR413PM is functionally implicated in drought and salt stress responses.
Previous studies have shown that heterologous overexpression of CpCOR413PM1 from wintersweet enhances cold and drought tolerance in Arabidopsis[15]. Overexpression of LeCOR413PM2 from tomato enhances the cold tolerance of transgenic tomato[32]. Overexpression of SikCOR413PM1 from Saussurea involucrata enhances cold and drought tolerance in tobacco and cotton[18,31]. Overexpression of PsCor413pm2 from Phlox subulata enhances cold tolerance in Arabidopsis[20]. In this study, overexpression of DhCOR413PM1 from Den. Sonia 'Hiasakul' not only enhanced cold tolerance in Arabidopsis, but also improved drought and salt tolerance. These results suggest that DhCOR413PM1 is involved in the response to low-temperature, drought, and salt stress in Denphal-group Dendrobium cultivars.
In this study, it was found that under cold and drought stresses, the expression of DhCOR413PM1 was first down-regulated and then up-regulated at 4 and 12 h after treatment, respectively. This pattern suggests that DhCOR413PM1 is crucial for maintaining membrane integrity during normal growth and stress responses, but its induction under stress is maintained by other regulators. This process may involve physiological and biochemical adjustments in the plasma membrane[34]. Furthermore, molecular regulatory networks fine-tune gene expression and modification to maintain metabolic stability. These networks integrate multiple signaling pathways to mount a coordinated defense.
As a plasma membrane-localized protein, COR413PM1 may serve as a membrane stabilizer under stress-countering lipid phase transitions during freezing and mitigating osmotic and oxidative damage during drought[35]. Plasma membrane fluidity is closely linked to cold tolerance, and stress-induced rigidification can trigger COR gene expression, as shown in Medicago sativa and Brassica napus[36−38]. COR413PM1 may also contribute to osmotic adjustment and oxidative balance. For example, SsCor413-1(Saccharum spontaneum) overexpression increased proline accumulation, enhanced activities of sodium dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), and reduced MDA content under stress[39].
This study demonstrates that overexpression of DhCOR413PM1 leads to an increase in the survival rate of transgenic Arabidopsis thaliana under freezing conditions, an improvement in germination rate under osmotic conditions, and a longer primary root under salt stress conditions. These phenotypic enhancements highlight the functional importance of the COR413PM1 protein in coordinating broad-spectrum stress resistance. The molecular and cellular basis of this cross-protection effect may involve the synergistic action of multiple mechanisms. Expression of the COR413PM is often regulated by the ICE-CBF-COR signaling pathway[11]. Promoter analyses of COR413PM1 homologs in Prunus persica and Chimonanthus praecox confirm enrichment of cold- and dehydration-responsive elements (DRE/ABRE), supporting their function as transcriptional hubs in stress responses[14,15]. Abscisic acid (ABA) signaling also contributes critically: in Arabidopsis, AtCOR413PM1 is strongly ABA-induced, and its loss disrupts downstream ABA responses, linking it to hormone signaling and membrane protection[40,41]. Similarly, OsCor413tm1 (Oryza sativa) improves drought tolerance in rice via an ABA-dependent pathway directly activated by OsABF1[42]. Calcium signaling represents another regulatory layer- PsCOR413PM2 in Phlox subulata amplifies cold-responsive gene expression, including CBFs, by enhancing Ca2+ signaling, forming a positive feedback loop[20]. Furthermore, COR413 proteins help maintain osmotic homeostasis through osmoprotectants such as LEA proteins, thereby reducing dehydration damage[18,43−45]. Therefore, the expression of DhCOR413PM1 is likely achieved by integrating these multiple signaling pathways (CBF, ABA, Ca2+), enabling it to make precise responses to various stresses.
In summary, DhCOR413PM1 is crucial for combined cold, drought, and salt stress tolerance. Under multi-factor regulation, it stabilizes membranes, maintains homeostasis, and sustains metabolism to enhance resistance. Future studies are needed to fully elucidate its molecular mechanisms.
-
DhCOR413PM1 encodes a 203-amino-acid protein with a calculated molecular mass of 22.42 kDa. The protein contains a completely conserved COR413 domain (residues 10–190) and five transmembrane domains. DhCOR413PM1 is ubiquitously expressed across all tissues of the plant, with predominant transcript accumulation in roots and reproductive organs, notably within 3 cm inflorescence meristems and in the labellum and gynandrium of 5 mm floral buds. DhCOR413PM1 exhibited significant transcriptional upregulation in response to low temperature (4 °C), drought (15% PEG-6000), and salinity (50 mM NaCl) stresses, with peak inductions of 2.13-fold at 8 h under 4 °C, 1.57-fold at 24 h under 15% PEG, and 1.88-fold at 120 h under 50 mM NaCl. Furthermore, transgenic Arabidopsis lines overexpressing DhCOR413PM1 exhibited markedly enhanced tolerance to multiple stresses: higher survival under freezing stress, increased germination rates, and longer roots under osmotic and salt stress, indicating significantly enhanced tolerance to cold, drought, and salt stresses, and demonstrating the gene's unique role in conferring multi-stress resistance. These results suggest that DhCOR413PM1 is a stress-responsive gene, potentially involved in multiple abiotic stress tolerance pathways in Dendrobium. Further functional characterization of this gene may provide valuable insights into the molecular mechanisms underlying stress adaptation in orchids.
The Hainan Natural Science Fund Project (Grant No. 324QN311), the Central Public interest Scientific Institution Basal Research Fund (Grant Nos 1630032022004 and 1630032023014), the earmarked fund for CARS(CARS-23-G60), and the Hainan Major Science and Technology Program (Grant No. ZDKJ2021015). We would like to thank the editor and reviewers for their helpful comments on the manuscript.
-
The authors confirm their contributions to the paper as follows: study conception and design: Lu S; data curation: Luo X, Li C,Yin J; analysis and interpretation of results: Yi S, Mo S; resources: Yu X, Liao Y; writing−original draft: Yi S, Mo S, Lu S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article, and also available from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Shuangshuang Yi, Shunjin Mo
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yi S, Mo S, Yu X, Luo X, Liao Y, et al. 2026. Overexpression of the DhCOR413PM1 gene from Dendrobium Sonia 'Hiasakul' enhances cold, drought and salt tolerance in Arabidopsis. Ornamental Plant Research 6: e007 doi: 10.48130/opr-0025-0051 

Overexpression of the DhCOR413PM1 gene from Dendrobium Sonia 'Hiasakul' enhances cold, drought and salt tolerance in Arabidopsis
- Received: 15 September 2025
- Revised: 24 November 2025
- Accepted: 16 December 2025
- Published online: 02 February 2026
Abstract: Denphal-group Dendrobium, a globally renowned orchid variety cultivated for both cut flowers and potted plants, is primarily distributed and grown in tropical regions but remains highly susceptible to low-temperature injury in many subtropical areas. To improve the cold tolerance of Denphal-group Dendrobium, in this study, a cold-responsive protein gene, DhCOR413PM1, was identified from the cultivated variety Denphal-group Dendrobium Sonia 'Hiasakul'. The DhCOR413PM1 ORF (612 bp) encodes a 203-amino-acid protein, containing five transmembrane domains and a conserved COR413 domain, showing the closest homology to DtCOR413PM1 (Den. thyrsiflorum). DhCOR413PM1 is ubiquitously expressed across all tissues of the plant, with predominant transcript accumulation in roots and reproductive organs. DhCOR413PM1 expression was significantly upregulated not only under low temperature (4 °C) but also in response to drought (15% PEG-6000) and salinity (50 mM NaCl) stresses. Transgenic Arabidopsis lines overexpressing DhCOR413PM1 exhibited markedly enhanced tolerance to multiple stresses, including higher survival under freezing stress, increased germination rates, and longer roots under osmotic and salt stress, indicating significantly enhanced tolerance to cold, drought, and salt stresses and demonstrating the unique role of the gene in conferring multi-stress resistance. These results suggest that DhCOR413PM1 is a stress-responsive gene potentially involved in multiple abiotic stress tolerance pathways in Dendrobium. Further functional characterization of DhCOR413PM1 may provide valuable insights into the molecular mechanisms underlying stress adaptation in orchids.
-
Key words:
- Dendrobium /
- DhCOR413PM1 /
- Transgenic Arabidopsis /
- Cold, drought and salt stress /
- Overexpression